新疆地区贝母属物种亲缘关系研究△
2018-05-31罗丽李会娟魏雪苹魏建和刘海涛李晓瑾樊丛照张本刚齐耀东
罗丽,李会娟,魏雪苹,2,魏建和,刘海涛,2,李晓瑾,樊丛照,张本刚,2,齐耀东,2*
(1.中国医学科学院 北京协和医学院 药用植物研究所 中草药物质基础与资源利用教育部重点实验室,北京 100193;2.中国医学科学院 北京协和医学院 药用植物研究所 中药资源教育部工程研究中心,北京 100193;3.新疆维吾尔自治区 中药民族药研究所,新疆 乌鲁木齐 830002)
百合科贝母属FritillariaL.物种丰富,全世界约140种,主要分布于北半球温带地区[1-3],属下常分为8个亚属:贝母亚属Subg.Fritillaria、多花亚属Subg.Rhinoptalum、Subg.Japonica、单鳞亚属Subg.Theresia、聚花亚属Subg.Petilium、多鳞亚属Subg.Liliorhiza、Subg.Davidii、Subg.Korolkowia[1]。中国约有24种[4],分别属于贝母亚属、多花亚属、多鳞亚属。我国贝母属植物按照分布区域划分,通常分为横断山区及邻近地区贝母群、长江中下游地区贝母群、新疆地区贝母群[5-8],其中新疆是一个相对独立分布区域,本区域分布的该属物种在我国其他地区没有分布。该地区贝母属植物约有9种[9],除砂贝母为多花亚属外,其余8种均属贝母亚属。贝母为传统药材,具清热化痰、止咳散结的功效,其来源为贝母属植物[10]。新疆地区分布的新疆贝母F.walujewii、伊犁贝母F.pallidiflora被《中华人民共和国药典》收录为伊贝母基原,然而我们的野外调查和文献报道[11]均表明,当地大多数贝母属物种均作伊贝母使用。
植物亲缘关系是植物类群在系统发生上显示的演化关系,在一定程度上是药用植物生物学本质的具体体现[12],形态性状与分子标记相结合的方法是研究植物亲缘关系的重要手段。形态学方面,前人对新疆地区贝母属物种叶、花粉、茎[13-16]显微特征及核型[17-18]进行了研究。分子标记方面:Cai等[19],Wang等[20]用RFLP的分子标记对贝母属部分物种的鉴定进行了研究;王果平等[21]和詹羽娇等[22]均利用ISSR探讨了新疆贝母属植物的遗传多样性,结果存在差异。Rønsted等[23]用matK、rpl16、ITS三个片段对贝母属及百合属构建了系统发育树,赖宏武等[24]也选用这三个片段联合对长江中下游地区贝母属物种亲缘关系进行了研究,Day等[25]选用matK、rpl16、rbcL和11对低拷贝核基因探究了贝母属93个物种间的进化关系,构建了贝母属的属下系统发育框架。但是上述研究或者焦点未集中在亲缘与进化探究,或者对新疆地区分布的物种取样不足。因此,对新疆地区贝母属物种的亲缘关系尚不明确。肖培根等[26]认为亲缘关系相近的物种生物活性和疗效往往有很大的相似性,该地区贝母属物种能否作为同一类贝母药材使用,分子系统学的研究能够为其亲缘关系提供数据支持。基于前人研究结果[23-25,27-29],我们选用叶绿体基因片段matK、rpl16和核基因间隔区ITS,探讨该地区贝母属物种亲缘关系,为该地区贝母类药材的合理使用提供依据。
1 材料与方法
1.1 实验材料
本研究选取17个物种,贝母属包含新疆地区分布的7个物种、东北地区分布的1个物种,横断山区及邻近地区分布的2个物种,共21个居群,其中16个为野生,5个为栽培。所用实验样品均为硅胶干燥的新鲜叶片;另有7个物种的序列来自GenBank。采集信息及序列号详见表1。
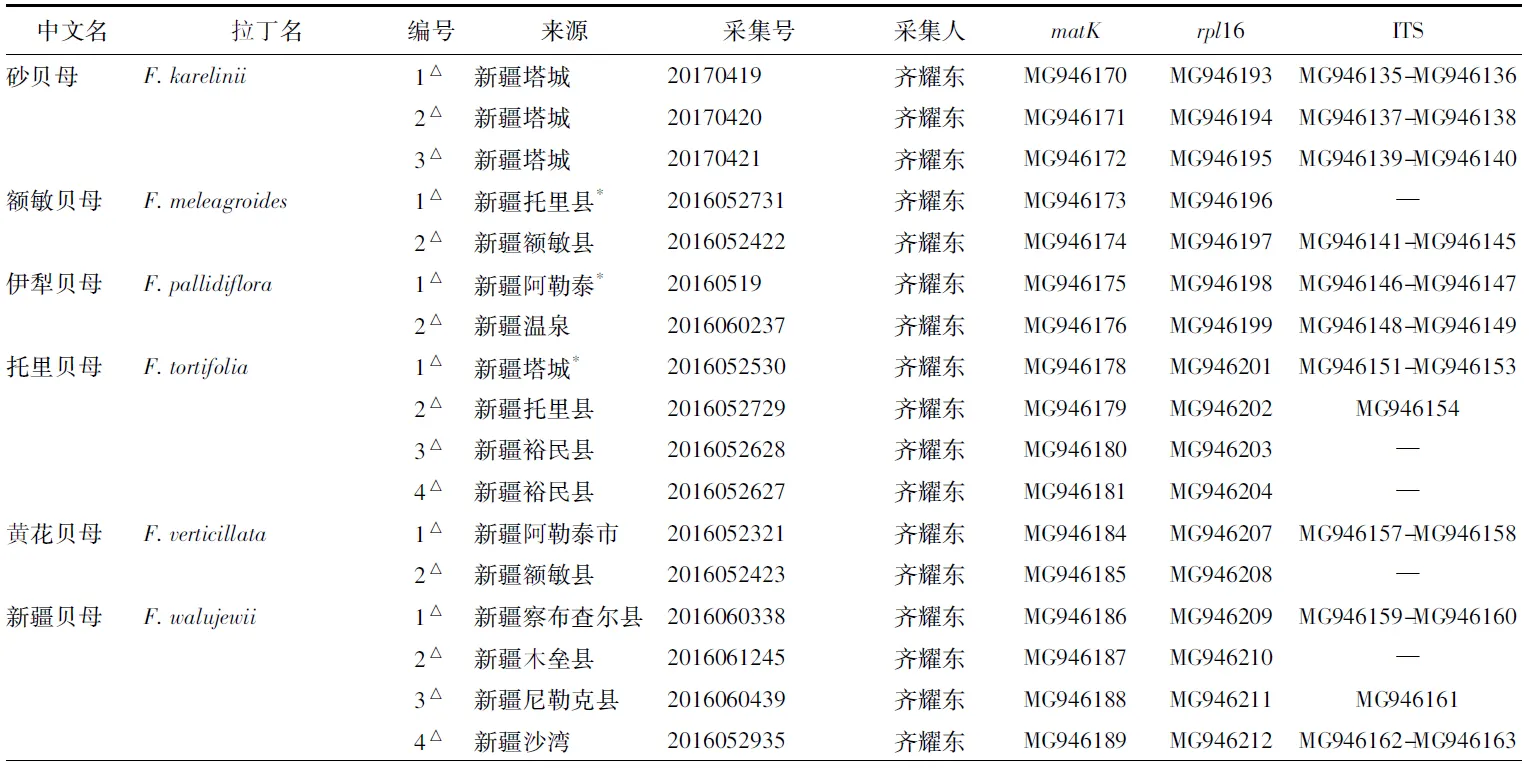
表1 样品详细信息及GenBank登录号
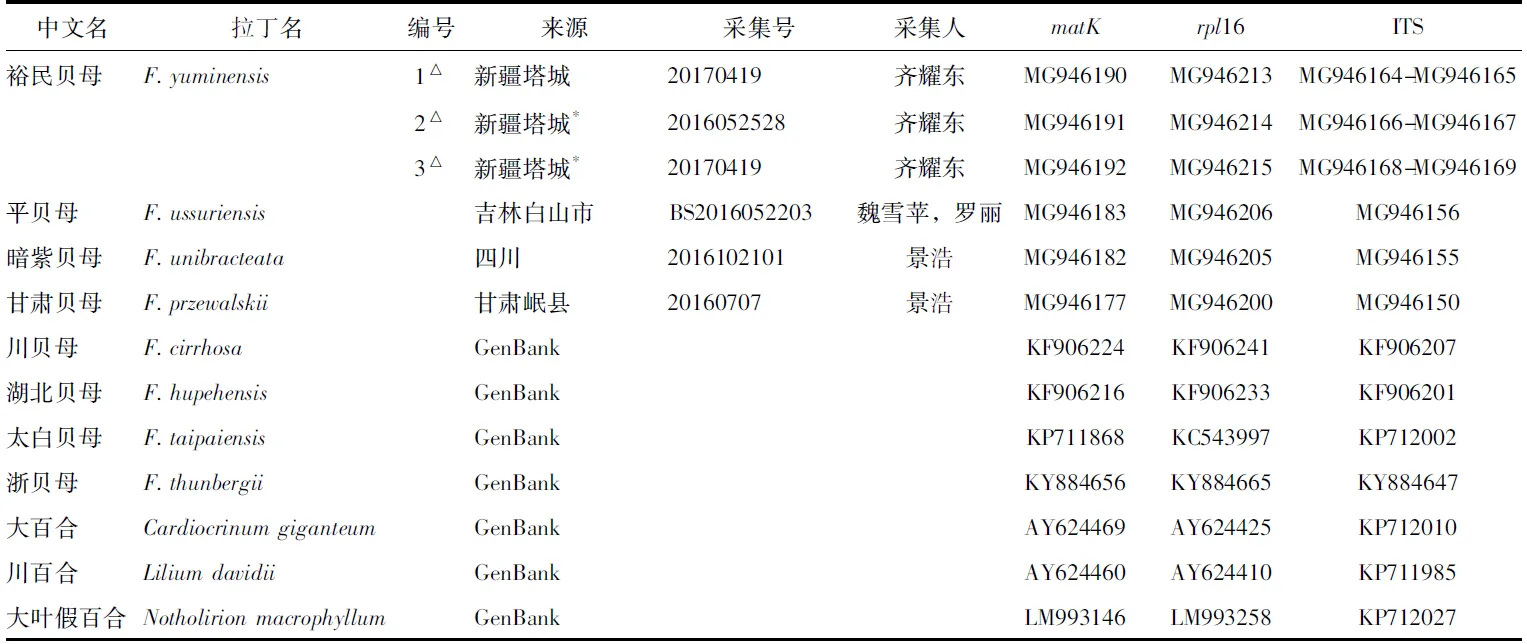
表1(续)
1.2 植物总DNA提取
根据植物基因组DNA提取试剂盒(Tiangen Biotech Co.China)说明书进行提取,提取后产物置于-20 ℃条件下保存,用于目标序列的扩增。
1.3 引物设计合成
实验选用叶绿体片段matK、rpl16,nrDNA-ITS进行扩增,引物信息见表2,引物均由生工生物工程(上海)股份有限公司合成。

表2 引物信息
1.4 PCR扩增与测序
利用表2中ITS、matK、rpl16引物在BIO-RAD PCR仪中进行样品PCR扩增,反应体系25μL,包括:2×Taq PCR Master Mix12.5 μL(北京艾德莱生物科技有限公司),2.5 μmol·L-1引物各1 μL,样品总DNA 2 μL,ddH2O 8.5 μL。
扩增程序:94 ℃预变性(ITS 3min ;matK,rpl16 5 min);94 ℃变性1 min,退火1 min(ITS 58 ℃;matK 53 ℃,rpl16 54 ℃),72 ℃延伸(ITS 2 min;matK 2.5 min,rpl16 2 min)(34个循环),72 ℃延伸(ITS 5 min;matK 7 min,rpl16 5 min),12 ℃保存。
1.5 序列测序
PCR产物经纯化后直接测序,其中ITS有双峰的序列,克隆测序得到的多条序列,均用于后续分析。测序由北京美吉桑格生物医药科技有限公司完成。
1.6 序列分析
测序序列用CodonCode Aligner v.3.7(Codon Code Corporation,Dedham,MA,USA)进行拼接,拼接后的序列结合GenBank下载序列在BioEdit v.7.0.9.0[34]中进行手工排序。以大百合Cardiocrinumgiganteum、大叶假百合Notholirionmacrophyllum、川百合Liliumdavidi为外类群,用最大似然法(Maximum likelihood,ML)、最大简约法(Maximum parsimony,MP)、贝叶斯分析法(Bayesian inference,BI)分别构建叶绿体联合、ITS片段系统发育树。ML分析在RAxML网络版[35]中完成;MP分析利用PAUP v.4.0[36]完成;BI分析利用MrBayes v.3.2[37]完成。利用jModelTest v.2.1.7[38]评估ITS、叶绿体联合片段在ML、BI分析中的最适核苷酸替代模型分别为GTR+G、TIM1+G。ML分析自展值(bootstrap value,BSML)通过快速自展式搜索(rapid bootstrap search)100次得到。MP分析中,目标核苷酸性状等同加权,gap作缺失处理,用每步增加一棵树的方法,任意重复1000次进行启发式搜索,TBR分枝交换法获取系统树,得到的系统树分支的可靠性评价使用自展分析,1000 次重复取样,1000次随机序列加入,最后得到MP自展值(BSMP)。BI分析采用马尔科夫链蒙特卡洛(Markov Chain Monte Carlo,MCMC)方法,以随机树为起始树,运行100 000 0代,每1000代取样1次。舍弃10%的老化样本后,对其余所有的树计算其多数一致树,最后获得各分枝的后验概率(posterior probability,PPBI)。
2 结果
2.1 序列信息
实验共测得ITS序列35条,matK、rpl16各23条。贝母属属下物种ITS序列长度为503~634 bp,排列长度为636 bp,变异位点为67个,信息位点48个;matK序列长度和排序长度均为1235 bp,变异位点为48个,信息位点39个;rpl16序列长度为904~953 bp,排列长度为983 bp,变异位点为38个,信息位点30个。
2.2 系统发育树重建
2.2.1matK、rpl16联合分析matK、rpl16联合数据构建的3种系统发育树拓扑结构一致,本文仅展示了BI系统发育树,见图1。
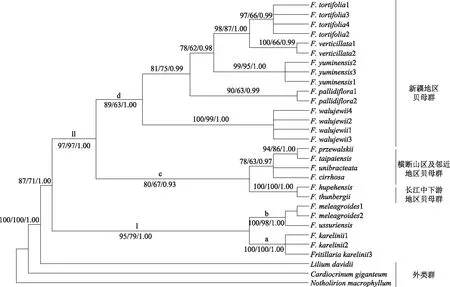
图1 采用BI构建的mat K、rpl 16数据系统发育树
结果显示:在系统发育树中贝母属物种构成具有较高支持的一支(BSML=87/BSMP=71/PPBI=1.00),砂贝母和额敏贝母、平贝母构成Ⅰ支,砂贝母的不同个体构成亚支a(BSML=100/BSMP=100/PPBI=1.00)、平贝母和额敏贝母构成亚支b(BSML=100/BSMP=98/PPBI=1.00),二者均为单系。分支Ⅱ分成两个亚支c(BSML=80/BSMP=67/PPBI=0.93)、d(BSML=89/BSMP=63/PPBI=1.00)。亚支c包含长江中下游(湖北贝母、浙贝母)和横断山区及附近地区贝母群(川贝母、甘肃贝母、太白贝母、暗紫贝母),二者分别构成单系;亚支d为新疆地区贝母群(新疆贝母、伊犁贝母、裕民贝母、托里贝母、黄花贝母)。在亚支d中,同物种内不同个体分别聚为单系,新疆贝母位于基部,为该地区其他物种的姐妹群;而伊犁贝母与裕民贝母、黄花贝母、托里贝母组成姐妹群。
2.2.2 ITS序列分析 ITS数据构建的ML、MP、BI系统发育树拓扑结构基本一致,本文仅展示了BI系统发育树,见图2。结果显示:基于ITS数据构建的系统发育树与基于叶绿体数据构建的系统发育树拓扑结构略有差异:ITS数据显示贝母属物种构成单系,但支持率较低(BSML<50/BSMP=79/PPBI<0.70);在亚支d中,新疆地区贝母群同物种内不同个体不聚成单系;且黄花贝母、托里贝母、裕民贝母、伊犁贝母和新疆贝母种间和种内个体相互嵌入。
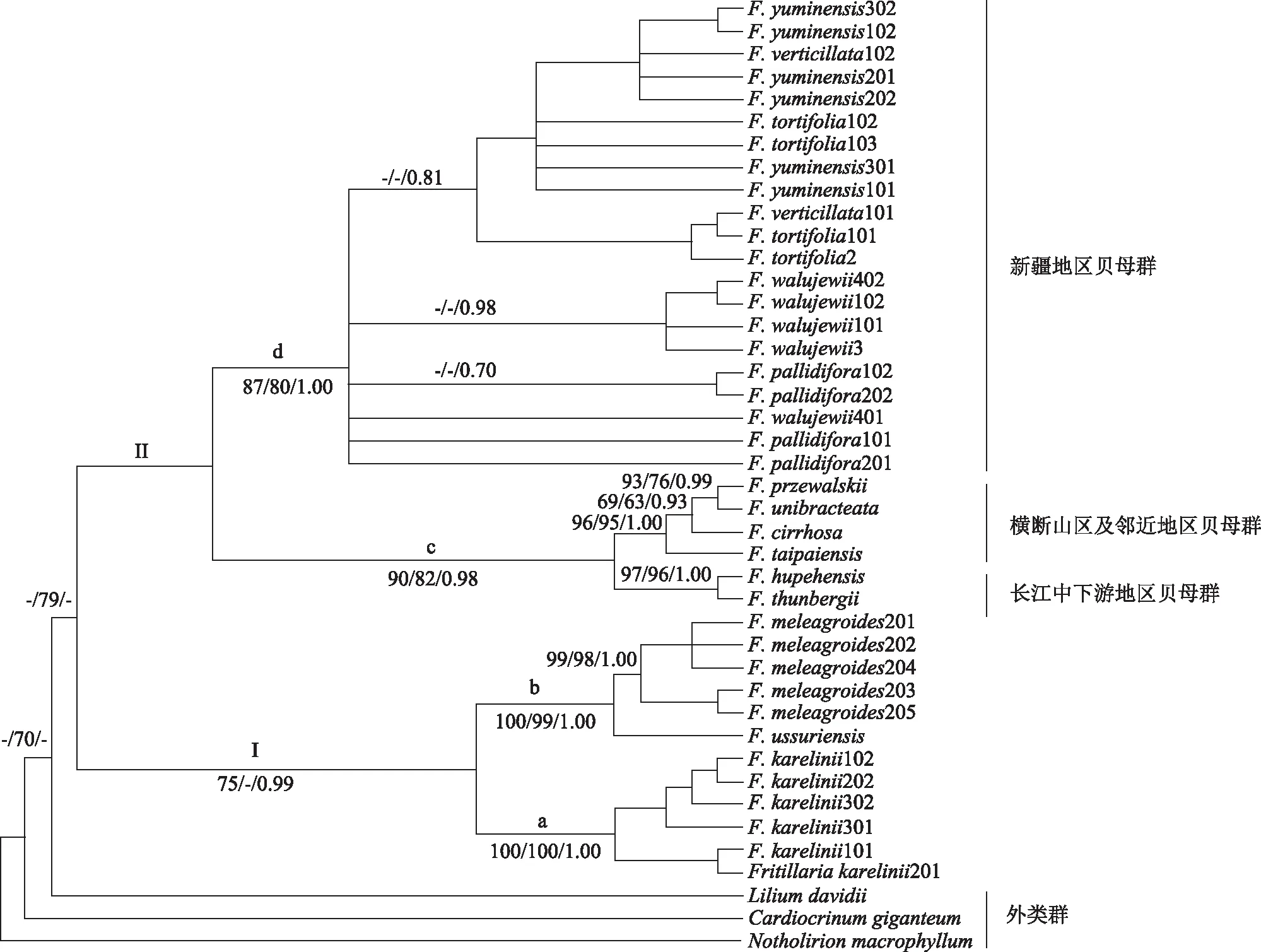
注:“-”示BSML/BSMP <50,PPBI <0.70。图2 采用BI构建的mat K、rpl 16 ITS联合数据系统发育树
3 讨论
3.1 新疆地区贝母属物种亲缘关系
目前对于新疆地区贝母属物种亲缘关系的研究较少,仅王果平等人[21]、詹羽娇等人[22]利用ISSR分析做过较为全面的研究,但二者的结论有所差异:王果平等人认为该地区该物种分为四类,第一类为新疆贝母、伊犁贝母、裕民贝母,第二类为塔城贝母(裕民贝母变种)、大白花贝母(黄花贝母变种)、托里贝母、小白花贝母(黄花贝母变种)、黄花贝母,第三类额敏贝母,第四类砂贝母;同一类的物种亲缘关系较近。詹羽娇等人则分为另外四类,第一类新疆贝母、伊犁贝母,第二类大白花贝母、托里贝母、黄花贝母,第三类额敏贝母、裕民贝母,第四类砂贝母。裕民贝母的归类存在明显争议。本研究利用ITS、叶绿体数据构建的系统发育树支持新疆地区贝母属物种主要分为三群:伊犁贝母、新疆贝母与托里贝母、黄花贝母、裕民贝母为一群(新疆地区贝母群);额敏贝母与东北地区的平贝母构成一群;砂贝母为单独一支。这与Day等人[25]利用matK、rpl16、rbcL构建的贝母属8个亚属代表物种系统发育树拓扑结构一致。形态上[4,9],新疆贝母群物种植株高大、花丝均无小乳突,区别于额敏贝母、平贝母和砂贝母植株矮小、花丝具小乳突特征;额敏贝母与平贝母均为紫花又能很好地与砂贝母粉花区分;此外砂贝母其独有特征,如全株被乳突状毛,花粉外壁纹饰拟脑纹型[14]。这些形态特征很好地支持了其在系统树上的位置,表明了托里贝母、黄花贝母、裕民贝母与伊犁贝母、新疆贝母亲缘关系较近;额敏贝母、砂贝母与伊犁贝母、新疆贝母亲缘关系较远。
3.2 三个片段的在贝母属系统发育分析中适用性
叶绿体基因构建的系统发育树上,新疆地区贝母群很好地聚为一支,5个物种的多个个体各自构成单系;而ITS构建的系统发育树上,同样支持新疆地区贝母群聚为一支,但物种间没有分辨率,且同一物种不同个体或同一个体不同拷贝相互穿插,物种间无法有效区分。研究表明,叶绿体和核基因系统发育树存在冲突可能的原因包括杂交、基因渗入,不完全谱系分选、基因重复等[39-40]。本研究的这种不一致一方面可能是新疆地区贝母群物种间存在上述现象导致,另一方面可能与贝母属物种基因组巨大、复杂[41]、ITS存在多个拷贝和协同进化有关。Rønsted等人[23]在贝母属和百合属的研究中也存在类似情况。因此,贝母属属下分类与鉴定研究中,叶绿体基因比ITS更为适合,深入理解贝母属的亲缘关系和物种形成还需要更多的核基因数据。
3.3 新疆地区贝母属物种作为贝母药材使用合理性
植物的亲缘关系、化学成分与疗效间存在潜在关联[26,42-44],物种间亲缘关系、化学成分的比较研究有助于发现植物的药用价值,开发新的药用植物资源[45]。贝母为多基原药材,植物亲缘关系研究、化学成分比较对其临床用药安全、有效及新基原资源开发具有重要意义。研究表明,贝母类药材镇咳平喘的活性成分为总生物碱[46],单体成分贝母甲素verticine、贝母乙素verticinone、西贝素imperialine、西贝素苷imperialine-β-Dglucoside和贝母辛peimisine等具有良好的镇咳平喘的作用[47-48],西贝素及西贝素苷还作为伊贝母药典检测成分[10]。结合本研究亲缘关系探究结果,该研究有以下观点。
新疆地区贝母群 本研究表明裕民贝母、托里贝母、黄花贝母与伊贝母的基原植物伊犁贝母和新疆贝母具有较近的亲缘关系。已有的化学成分研究表明新疆贝母群的5个物种均含有贝母乙素、西贝素、西贝素苷3种生物碱类成分[49-52]。因此,裕民贝母、托里贝母、黄花贝母具有作伊贝母药材使用的潜在可能。
额敏贝母 在系统发育树上,额敏贝母与东北分布的平贝母互为姐妹支,而与新疆地区其它贝母属物种亲缘关系较远。其化学成分研究相对较少,仅张鹏葛等[53-54]报道额敏贝母含有生物碱类化合物,但其含量均低于新疆地区其它贝母属植物,单体化合物尚未报道,因此从化学成分上难以判断是否可作为贝母类药材使用。
砂贝母 在系统发育树上,砂贝母也与新疆地区其它贝母属物种亲缘关系较远,与额敏贝母、平贝母共同构成中国贝母类群的基部分支。化学成分的研究显示砂贝母的生物碱含量仅高于额敏贝母,低于新疆地区其它贝母属物种[53-54],目前也仅报道西贝素一个单体成分[55],从化学成分上也难以判断是否可作为贝母类药材使用。
4 结论
通过系统发育树的重建发现,新疆地区分布的黄花贝母、托里贝母、裕民贝母、伊犁贝母、新疆贝母亲缘关系较近;额敏贝母与东北地区分布的平贝母亲缘关系较近,砂贝母与之互为姐妹群,位于基部。伊贝母药材两基原植物亲缘关系相近的3个物种,含有与之相同的有效成分,有作为伊贝母药材使用的潜在可能,作为贝母类药材使用较为合理;额敏贝母和砂贝母与伊贝母药材两基原亲缘关系较远,作为伊贝母使用值得商榷;额敏贝母与平贝母互为姐妹关系,但其化学成分研究相对较少,能否作为平贝母药材使用有待进一步研究。
[1] Rix E M.Fritillaria:A Revised Classification Together with an Updated List of Species[M].UK:Publication of the Fritillaria Group of the Alpine Garden Society,2001.
[2] Hill,L.Fritillaria:A List of Published Names.v4.0.[EB/OL]< www.fritillariaicones.com/info/names/frit.names.pdf >(last accessed 26.11.13).
[3] WCSP,2014.World Checklist of Selected Plant Families.Facilitated by the Royal Botanic Gardens,Kew[EB/OL].
[4] Chen S Q,Mordak H V.Fritillaria Linnaeus.In:Wu Z Y & Peter P H(Eds.)Flora of China[M].Beijing:Science Press,Missouri Botanical Garden Press,St.Louis,2000:127-133.
[5] 罗毅波,陈心启.中国长江中下游地区贝母属的修订[J].植物分类学报,1995,33(6):592-596.
[6] 罗毅波,陈心启.中国横断山区及其邻近地区贝母属的研究(一)——川贝母及其近缘种的初步研究[J].植物分类学报,1996,34(3):304-312.
[7] 罗毅波,陈心启.中国横断山及其邻近地区贝母属的研究(二)[J].植物分类学报,1996,34(5):547-553.
[8] 罗毅波,陈心启.新疆贝母属的订正[J].植物分类学报,1996,34(1):77-85.
[9] 新疆植物志编辑委员会.新疆植物志:第3卷[M].新疆科学技术出版社,1996:505-517.
[10] 国家药典委员会.中华人民共和国药典:一部[S].北京:中国医药科技出版社,2015:36-38,97-98,141-142,292,348-349.
[11] 刘勇民.维吾尔药志[M].上册.新疆:新疆科技卫生出版社,1999:59-66.
[12] 杨洪军,唐仕欢,黄璐琦,等.基于亲缘关系的中药药性研究[J].中国中药杂志,2008,33(24):2983-2985.
[13] 李萍,蒋鑫,刘惠娟,等.贝母属植物叶表面显微观察研究[J].中国药科大学学报,1993,24(4):205-211.
[14] 李萍,濮祖茂,徐珞珊,等.中国贝母属花粉形态的研究[J].云南植物研究,1991,13(1):41-46,113-115.
[15] 张鹏葛,盛萍,任慧梅,等.新疆贝母属8种药用植物叶片的解剖学结构[J].江苏农业科学,2016,44(5):267-270.
[16] 张鹏葛,盛萍,任慧梅.新疆贝母属8种药用植物鳞茎的解剖学、组织化学和植物化学研究[J].中国药学杂志,2015,50(23):2028-2034.
[17] 翟诗虹,刘国钧,李懋学,等.新疆贝母属的核型研究[J].植物分类学报,1985,23(4):264-269.
[18] 陆静梅,赵毓棠.中国三种贝母染色体数目及核型研究[J].东北师大学报(自然科学版),1990(4):89-92.
[19] Z H Cai,P Li,T T Dong,et al.Molecular diversity of 5S-rRNA spacer domain in Fritillaria species revealed by PCR analysis.[J].Planta medica,1999,65(4):360-364.
[20] C Z Wang,P Li,J Y Ding,et a1.Simultaneous identification of Bulbus Fritillariae cirrhosae using PCR-RFL Panalysis[J].Phytomedicine,2007,14:628-632.
[21] 王果平,樊丛照,李晓瑾,等.基于ISSR的新疆贝母属植物遗传多样性研究[J].中草药,2013,44(7):887-890.
[22] 詹羽姣,盛萍,姚蓝,等.新疆贝母属8种药用贝母遗传多样性ISSR分析[J].中国野生植物资源,2015(4):1-6.
[23] Rønsted N,Law S,Thornton H,et al.Molecular phylogenetic evidence for the monophyly of Fritillaria and Lilium(Liliaceae;Liliales)and the infrageneric classification of Fritillaria[J].Molecular Phylogenetics and Evolution,2005,35(5):509-527.
[24] 赖宏武,齐耀东,刘海涛,等.贝母类药材湖北贝母Fritillariahupehensis系统位置的探讨——来自ITS,rpl16,matK序列的证据[J].中国中药杂志,2014,39(17):3269-3273.
[25] Day P D,Berger M,Hill L,et al.Evolutionary relationships in the medicinally important genus Fritillaria L.(Liliaceae)[J].Molecular Phylogenetics and Evolution,2014,80(1):11-19.
[26] 肖培根.植物亲缘关系、化学成分和疗效间的联系性[J].中国药学杂志,1978,13(1):1-5.
[27] 郑曼莉,夏永梅.利用ITS(nrDNA)和matK(cpDNA)探讨姜族植物的系统发育[J].云南大学学报:自然科学版,2010,32(1):426-432.
[28] 辛天怡,姚辉,罗焜,等.羌活药材ITS/ITS2条形码鉴定及其稳定性与准确性研究[J].药学学报,2012,(8):1098-1105.
[29] 丑欢欢,唐红.基于ITS和matK基因对牡丹组序列分析及其亲缘关系的研究[J].植物研究,2017,37(4):603-612.
[30] Molvray M,Kores P J,Chase M W.Polyphyly of mycoheterotrophic orchids and functional influences on floral and molecular characters[M].Monocots:systematics and evolution.Collingwood,Australia:CSIRO,2000:441.
[31] Cué noud Philippe,Savolainen Vincent,Chatrou Lars W,et al.Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL,atpB,and matK DNA sequences[J].American journal of botany,2002,89(1):132-144.
[32] Baum D A,Small R L,Wendel J F.Biogeography and floral evolution of baobabs(Adansonia,Bombacaceae)as inferred from multiple data sets.[J].Systematic biology,1998,47(2):181-207.
[33] Y Sun,Skinner D Z,Liang G H,et al.Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA[J].TAG.Theoretical and applied genetics.Theoretische und angewandte Genetik,1994,89(1):26-32.
[34] Hall T A,BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[C].Nucleic acids symposium series,1999,41:95-98.
[35] Stamatakis A,P Hoover,J Rougemont,A rapid bootstrap algorithm for the RAxML Web servers[J].Systematic Biology,2008,57(5):758-771.
[36] Swofford D L,PAUP*.Phylogenetic Analysis Using Parsimony(*and Other Methods).version 4.0b10[J].Sinauer Associates,Sunderland,MA.,2003.
[37] Fredrik R,Maxim T,Paul V D M,et al.MrBayes 3.2:Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space[J].Systematic Biology,2012,61(3):539-542.
[38] Darriba D,Taboada G L,Doallo R,et al.jModelTest 2:more models,new heuristics and parallel computing.[J].Nature Methods,2012,9(8):772.
[39] Baldwin B G,Sanderson M J,Porter J M,et al.Erratum:The ITS region of nuclear ribosomal DNA:A valuable source of evidence on angiosperm phylogeny(Annals of the Missouri Botanical Garden(1995)82(247-277))[J].Annals of the Missouri Botanical Garden,1996,82(2):247.
[40] Doyle J J.Gene trees and species trees:molecular systematics as one-character taxonomy.[J].Systematic Botany,1992,17(1):144-163.
[41] Bennett M D,Leitch I J.2012.Plant DNA C-values database(release 6.0,December 2012)[EB/OL].http://www.kew.org/cvalues/.
[42] 肖培根,陈碧珠,王立为,等.大黄属的植物亲缘关系、化学成分与疗效间联系性的初步研究[J].药学学报,1980,15(1):33-39.
[43] 肖培根.中国毛茛科植物群的亲缘关系、化学成分和疗效间相关性的初步探索[J].植物分类学报,1980,18(2):142-153.
[44] 肖培根,徐克学,宋晓明.大黄属植物的外形、成分与泻下作用问联系性的多元分析[J].中国中西医结合杂志,1982,(4):231-234,197.
[45] 陈四保,彭勇,陈士林,等.药用植物亲缘学[J].世界科学技术—中医药现代化,2005,7(6):97-103.
[46] 李萍,季晖,徐国钧,等.贝母类中药的镇咳祛痰作用研究[J].中国药科大学学报,1993,24(6):360-362.
[47] 周颖,季晖,李萍,等.五种贝母甾体生物碱对豚鼠离体气管条M受体的拮抗作用[J].中国药科大学学报,2003,34(1):58-60.
[48] 赵益,朱卫丰,刘红宁,等.贝母辛平喘作用及机制研究[J].中草药,2009,40(4):597-601.
[49] 张鹏葛.新疆贝母属8种药用贝母组织化学及活性成分研究[D].乌鲁木齐:新疆医科大学,2016:24-48.
[50] 徐东铭,黄思喜.伊贝母生物碱的研究[J].Acta Botanica Sinica,1990,32(10):789-793.
[51] 路熹.新疆贝母化学成分研究[D].天津:天津理工大学,2015:39-47.
[52] 曹新伟.川贝母的化学成分研究与贝母属药用植物质量评价[D].北京:北京协和医学院,2008:118-135.
[53] 张鹏葛,盛萍,任慧梅.新疆贝母属8种药用贝母地上部位与鳞茎生物碱含量差异研究[J].中国野生植物资源,2016,35(1):12-15.
[54] 张鹏葛,盛萍,任慧梅,等.新疆贝母属8种药用植物地上部位叶的组织化学研究[J].安徽农业科学,2015(7):28-31.
[55] 潘鑫复,朱子清.砂贝母中的植物碱[J].科学通报,1980,25(4):186-186.