植物WRKY转录因子的研究进展
2018-05-22梁滨董冬
梁 滨 董 冬
(山东省滨州市邹平县魏桥中学 山东滨州 256212)
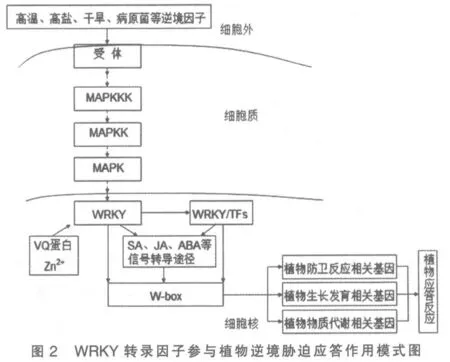
植物在生长过程中会受到生物逆境(病原菌侵入、病虫害等)和非生物逆境(高温、高盐、干旱等)的胁迫(图1),从而抑制植物的生长发育,造成各种作物质量、产量的下降,给人类生产带来极大危害。在长期进化过程中,植物为了适应这些不利的环境因素,在体内形成了一套既复杂且又高效的植物防卫反应机制。当植物受到逆境胁迫时,胞外的信号(高温、高盐、干旱、病原菌等)通过一系列信号转导途径(例如通过MAPK 激酶等进行信号转导)级联放大进入细胞核内,通过与其他防御相关蛋白之间的相互作用,进而激活或抑制下游相关防御基因的转录,从而响应植物对相关逆境胁迫的应答反应[1]。其中,转录因子(MYC、NAC、WRKY、ERF、bZIP 等)在这一系列调控中起到非常重要的作用[2]。在这一系列的转录因子中,WRKY是近年研究比较广泛的一类转录因子,研究表明,WRKY 在植物生长发育和防卫反应机制中起重要调控作用。

转录因子WRKY 是近年在植物中新发现的一类超大家族的转录调控因子[2],已发现的WRKY转录因子,在拟南芥、水稻等作物中参与免疫反应调节的WRKY 家族成员既有起正调节作用的,也有起负调节作用的,但二者的N-末端均含有高度保守的WRKYGQK 氨基酸序列[3]。不同的WRKY家族成员均通过核心结构域(WRKYGQK)结合靶基因启动子区域保守的W-BOX 调控基因转录表达,由此产生的一个突出问题是: 不同的WRKY家族成员如何实现其在植物应答逆境过程中的特异功能?最近大量报道表明,除W-BOX 侧翼序列能在一定程度上影响WRKY 蛋白特异调节靶基因外,还能与其他蛋白的互作并调节其功能以实现功能的特异性。该家族的WRKY 基因分别相继从甜马铃薯、野燕麦、皱叶欧芹、拟南芥等植物中分离获得。同时,诸多专家也研究了其相应的分子生物学功能,目前已在拟南芥中发现WRKY 基因74 个,玉米中发现119 个,水稻中发现105 个,油菜中发现46 个。这些研究都表明了WRKY 转录因子确实参与植物的多种逆境胁迫的应答,同时还参与植物生长的重要生理生化活动的调控[4-6],在植物抗逆反应过程中具有重要的作用(图2)。
1 WRKY 转录因子结构特征及分类
WRKY 转录因子是植物中广泛存在的一类超基因家族,通过其保守的氨基酸序列(WRKYGQK)对靶基因启动子区域的W-box[(T)TGACC(AT)]特异识别,并调控靶基因的转录表达。WRKY 家族在代谢调控、种子的萌发及发育衰老调控、植物防御应答、形态学建成等生命过程中都起到重要作用。
WRKY 是锌指结构的转录因子,它的DNA 结合区通常包含1 个或2 个保守的WRKY 核心结构域,根据其DNA 结合区的核心结构域的个数及锌指结构类型将WRKY 转录因子分为三大类:第1 类具有2 个WRKY 核心结构域及Cys2His2 型锌指结构;第2 类具有1 个WRKY 核心结构域及Cys2His2 型锌指结构;第3 类具有1 个WRKY 核心结构域及Cys2HisCys 型锌指结构。WRKY 转录因子是植物中最大的转录调节子家族之一(拟南芥中有74 个成员,水稻中100 多个成员,辣椒中有报道的也已经有40 多个),它能与各种蛋白质相互作用,包括MAP 激酶,MAP 激酶的激酶,14-3-3 蛋白,钙调蛋白,组蛋白去乙酰化酶,抗性蛋白和其他WRKY 转录因子,构成完整的信号网路调节许多植物生理过程。
植物有2 套固有的可诱导的免疫机制: 病原菌相关分子模式触发的免疫机制(PTI)及效应子触发的免疫机制(ETI)[2],而WRKY 转录因子在这2 种免疫机制中都起重要作用。拟南芥中AtWRKY48和AtWRKY7 在抗丁香假单孢杆菌中分别起正调节和负调节作用;AtWRKY51,AtWRKY70 在JA,SA,ET信号通路中起作用;AtWRKY25,AtWRKY40,AtWRKY28,AtWRKY70 同时受到生物及非生物胁迫的调控。同时WRKY 也受到机械损伤的诱导,例如烟草的NtWRKY6,NtWRKY3,参与糖代谢,以及对低温、干旱胁迫的应答。关于辣椒WRKY家族的研究还很不深入,到2014年NCBI 才公布了11 条辣椒WRKY 基因序列。
拟南芥中AtWRKY58 作为NPR1(获得性防御系统SAR 的一个负调节子)的下游基因参与NPR1 基因介导的信号传递。CaWRKY58 在辣椒对病原菌的防御反应过程中起到转录负调节作用,青枯病侵染的辣椒中,CaWRKY58 的表达先出现先下调后上调的情况,而且CaWRKY58 的表达被茉莉酸和脱落酸所抑制,同时将CaWRKY58 转化烟草,使烟草在病原菌侵染下表现出更严重的病状,同时在转化CaWRKY58 的烟草中很多重要的过敏反应相关的标记基因及一些防御相关的标记基因都出现下调,而且CaWRKY58 基因沉默后的辣椒植株表现出对病原菌的更耐受性及各种防御相关基因的上调。WRKY58 的作用可能是提供一个保障机制,以防止防卫反应的激活停留在高SA 水平。
WRKY 转录因子的结构含有DNA 结合区域、核定位信号区、转录调控区域和寡聚化位点[7]。结构特点是其DNA 结合区域中都至少含有一个WRKY 结构域,该结构域的N-末端含有高度保守的WRKYGQK 氨基酸残基和CX4-5CX22-23HXH 型或CX7CX23HXC 型的C-末端锌指结构。根据WRKY 的结构特征,可分为3 种类型:Ⅰ类为WRKY 转录因子含有2 个WRKY 结构域,锌指结构类型为C2H2(CX4-5CX22-23HX1H),其DNA 结合功能主要是通过C-末端的WRKY 结构域介导;Ⅱ类为WRKY转录因子只含1 个WRKY 结构域,锌指结构类型也为C2H2(CX4-5CX22-23HX1H),而且大部分高等植物WRKY 转录因子是属于这类型;Ⅲ类为WRKY 转录因子也仅含1 个WRKY 结构域,锌指结构为C2HC (CX7CX23HX1C),但这类型转录因子一般只存在高等植物中,在一些低等植物中是不存在的。
2 WRKY 蛋 白与W-box 结合
众多研究表明:WRKY 转录因子能特异结合启动子上的顺式作用元件W-box[(T)(T)TGAC(C/T)],其中高度保守序列TGAC 对于WRKY 的结合功能是必要的,任一核苷酸被替代,与之结合的能力都大幅度下降甚至可能完全消失。研究还发现WRKY 转录因子不仅与自身基因启动子上的顺式作用元件W-box 结合,而且与大量抗病相关基因启动子的W-box 结合参与表达调控[8],且这些W-box 多存在于抗病等相关基因的启动子区域,说明WRKY 转录因子参与植物的抗逆反应及其生长过程中生理生化等的调控。为了进一步研究W-box 顺式作用元件在基因表达调控中的作用,通常利用人工合成只含W-box 顺式作用元件的启动子研究其在植物中的防御反应[9],结果表明仅含W-box 的启动子也能介导植物的逆境胁迫和防卫反应。
3 WRKY 转录因子的生物学功能
3.1 参与生物逆境胁迫 目前大量报道表明,WRKY 转录因子在生物胁迫中的作用方面的研究既广泛又深入。植物免疫系统包括PTI (PAMPtriggered immunity),ETI (Effector- triggered immunity),MTI(MAMP-triggered immunity)及SAR(Systemic acquired resistance)等多种免疫系统,WRKY基因在其中发挥着正调控或负调控的作用,而且在参与植物的抗病反应,既存在正调控调节又有负调控途径[10]。
研究发现,拟南芥的大部分WRKY 基因受SA(水杨酸)和病原菌的诱导,并且这些WRKY 基因自身的启动子中富含W-box,说明这类转录因子在植物抗病反应中形成一个复杂的调控网络。很多WRKY 转录因子被报道能调控抗病基因(PR)的表达。拟南芥AtWRKY70 在SA(水杨酸)和JA(茉莉酸)途径中扮演了重要的调控角色,其功能的缺失将使植物易感染灰霉病菌、叶斑病菌和真菌性白粉病菌等[11]。拟南芥AtWRKY18,AtWRKY40和AtWRKY60 结构非常相似,参与植物抗病并且在功能上存在冗余[12],此外,AtWRKY18 还作为正调控因子参与系统获得性抗性,而AtWRKY40没有参与其中[13]。
另一方面,也有许多WRKY 基因参与植物防御的负调控。在拟南芥中,AtWRKY7 功能缺失体将导致细菌性叶斑病菌抗性的增强,且在AtWRKY7 和AtWRKY11 双突变中得到进一步的提高,说明两者在抗病过程中起到负调控作用[14]。AtWRKY25,AtWRKY38 和AtWRKY62 在植物防御中也起到负调控作用,其过表达植株都显著抑制 了PR 基因的表达[15-16]。
此外,还有许多非模式植物中WRKY 基因参与了抗病反应,如皱叶欧芹中PcWRKY1,2,3,4,5,红辣椒中CaWRKY1,40,大麦中的HvWRKY1,2,10,19,28 等都在抗病调控方面起到重要的作用。
3.2 参与非生物逆境胁迫 近年越来越多的研究证实了WRKY 基因在高温、高盐、机械损伤、干旱等非生物逆境胁迫下,其表达量会发生特异地变化,表明其在非生物逆境中扮演重要的角色,对于了解植物适应复杂多变的环境的分子机制具有重要的指导意义。qRT-PCR 研究发现,拟南芥AtWRKY25,28,40,70 均受到高盐、高温及渗透胁迫的诱导[17],AtWRKY25,26,33,39 则能应答热激反 应[18-19],AtWRKY57 能通过激活与ABA 相关基因的表达增强拟南芥耐干旱的能力[20]。此外还发现拟南芥AtWRKY7 中含有一个钙调节蛋白结合域,能特异结合钙调蛋白并介导Ca2+离子信号转导[21]。这些研究有力地说明了WRKY 转录因子在抵御高温、高盐、干旱和热等复杂环境中发挥着重要的作用。
3.3 参与植物的生长发育及物质代谢 研究表明,WRKY 家族还参与了调控植物种子形成、休眠和萌发及叶片衰老等生长发育及物质代谢等一系列生理活动。例如拟南芥AtWRKY10 参与种子的发育过程,AtWRKY6 不仅参与植物的防御应答而且还参与植物的衰老调控,拟南芥AtWRKY44 参与调控种皮和表毛皮的发育,其功能突变体后使得表毛皮出现未分支且数量显著下降,同样地,水稻OsWRKY78 也参与调控种子大小[5]。此外,棉花GaWRKY1 参与调控棉子酚的合成。这些结果都表明WRKY 基因在植物生长发育、物质代谢等生理过程中发挥了极其重要的作用。
4 展望
其实,在应用基因工程培育作物抗逆新品种方面,人们已经从植物基因组的角度进行了深入的研究,这些研究成果在分子水平上对植物的相关表达调控机制的研究具有重要的指导意义。而启动子是基因表达调控的重要元件,是基因在转录水平的表达模式及调控机制中的一个关键元素。因此,研究WRKY 转录因子等相关抗病基因启动子的调控特性,分离并分析其顺式作用元件的分布及逆境应答对于阐明植物抗病性极其重要,可为作物抗病育种开辟出一条新途径。大量报道表明,除了W-BOX 侧翼序列能在一定程度上影响WRKY 蛋白特异调节靶基因外,还能与其他蛋白的互作并调节其功能以实现功能的特异性。而迄今仅分离鉴定了部分的WRKY 蛋白的互作蛋白,大多数WRKY 家族成员的互作蛋白仍未见报道。因此,分离相关的互作蛋白,同时对其互作验证及功能和作用机制的研究将成为今后的研究热点。
