长江中游贝氏 繁殖生物学研究
2018-05-17余文娟沈建忠李春盛王克雄梅志刚
余文娟,沈建忠,龚 江,李 乾,李春盛,王克雄,梅志刚
(1.华中农业大学水产学院,武汉430070;2.湖北监利何王庙故道长江江豚省级自然保护区,湖北监利,433301;3.中国科学院水生生物研究所,武汉 430072)
长江何王庙故道(东经112°56′39″~113°01′48″,北纬29°29′30″~29°46′40″)位于湖北省监利县与湖南省华容县交界处,是1957年长江自然截弯取直形成的大“U”型通江故道,水面2 489 hm2[1]。由于故道水体无工业污染,水生植被群落丰富,整体生境稳定,是长江江豚(Neophocaenaasiaeorientalisasiaeorientalis)迁地保护的理想场所,所以2012年在此建立了江豚省级迁地自然保护区,是湖北省内继天鹅洲故道后的第二个江豚保护区,目前已有12头江豚生活其中。江豚主要以小型鱼类和大型鱼类幼鱼(5≤SL≤24 cm)为食[2],人工豢养的江豚常捕食鲫、 和似鳊等小型鱼类[3]。因此,对何王庙故道的小型鱼类,特别是对优势种类的资源进行评估和管理,于江豚保护十分重要。
贝氏 (Hemiculterbleekeri) 属鲤形目鲤科鲌亚科 属,俗称油 、白漂子,几乎遍布全国各大水系,为水体中上层小型经济鱼类,在许多水体成为优势种[4-8]。在何王庙故道,据2013年渔业资源调查,贝氏 占整个渔获物的 22.5%,是主要优势种,分析和预测其资源变化对江豚保护有重要意义。
目前,贝氏 生物学相关的研究仅见简单的生物学分析[4,9-10]以及食性分析[11]。本文通过对采自何王庙故道的长江中游贝氏 繁殖生物学的研究,旨在进一步了解其生活史对策,分析其在长江干流成为优势种的成因,同时为何王庙故道长江江豚迁地保护区渔业资源管理提供依据。
1 材料与方法
1.1 样本采集和处理
贝氏 样本采集结合长江何王庙故道小型鱼类资源调查进行,采集时间2016年4、6、8、9及12月,共采集591尾,主要来自三层刺网(网目2a=3 cm)和迷魂阵(网目2a=1 cm)。现场测量渔获物体长、全长(精确到0.1 cm)和体重(精确到0.01 g)等常规生物学数据。取背鳍下方、侧线上方鳞片5~10枚作年龄鉴定材料,根据鳞片年轮特征鉴定贝氏 的年龄。解剖、观察并记录其性别、性腺发育期等,性腺发育分期参考我国常用的六期标准划分方法来确定[12],同时测量性腺重、空壳重等生物学数据;用10%的甲醛溶液固定整个卵巢,用于卵径频率分布分析和繁殖力计算。
1.2 数据处理分析
比较不同年龄组、体长及月份的雌雄比例,同时,采用卡方检验性比是否偏离1∶1。
初次性成熟体长采用L50方法评估[13]。将繁殖季节采集到的所有贝氏 雌雄鱼个体分别以体长1 cm区间分组,以性腺发育达到Ⅲ期以上的个体作为性成熟个体,计算各体长组性成熟比例,以Logistic方程拟合各体长区间性成熟百分比,以此计算出50%(SL50)初次性成熟体长。拟合公式为:
P=100/(1+e-k(SLmid-SL50)
其中P为个体长区段性成熟比例,SLmid为体长区段中值,k为参数。
繁殖期根据成熟系数(GSI)的周年变化,结合观察到的性腺发育期来确定。GSI变化选取100%性成熟个体(SL≥SL100)进行统计分析。成熟系数计算公式为:
GSI=100×性腺重/体重。
产卵类型根据分析繁殖期间成熟鱼类卵径频率分布确定[14-15]。在4月和6月随机选取49个Ⅳ期卵巢样本,每个样本挑选100~200粒沉积卵黄的卵,平铺在培养皿中,在Leica EZ4D解剖镜下拍照,然后用Image Pro Plus v6.0软件测量卵粒直径,作出卵径频率分布图。
繁殖力采用重量法计算[2]。随机选取45个性腺发育为Ⅳ期的成熟雌鱼样本,分别称取0.5 g卵巢,计数所有有卵黄沉积(Ⅲ~Ⅳ时相)的卵粒数量。
绝对繁殖力(F)∶F(粒/尾)=每克成熟卵巢的卵粒数×卵巢重;
单位体重相对繁殖力(FW)∶FW(粒/g)=绝对繁殖力/体重;
单位体长相对繁殖力(FL)∶FL(粒/cm)=绝对繁殖力/体长。
繁殖力与体长(L)、体重(W)、卵巢重(GW)和成熟系数(GSI)等生物学指标的关系,采用线性函数、指数函数、幂函数、二次多项式函数方程拟合,以相关系数R2最大者作为拟合方程,并做相关性检验。
采用Excel2013、SPSS21.0和Lab Origin Pro 8.5进行数据处理和统计分析。
2 结果分析
2.1 性比
所采591尾贝氏 样本,性别未辨个体93尾,性别可辨个体中雌性294尾,雄性204尾。群体性比为1.44∶1,χ2检验显示,与1∶1有极显著性差异(χ2=16.265,P<0.05)。
各年龄组性比见表1。性别未辨个体均为1龄个体;性别可辨个体,1龄组中雌性明显多于雄性,雌雄比与1∶1存在及其显著性差异(χ2=18.968,P<0.05);2龄组雌雄样本量接近,性比无显著性差异;3龄组雌明显多于雄性,只是样本数少,检验显示无显著差异。
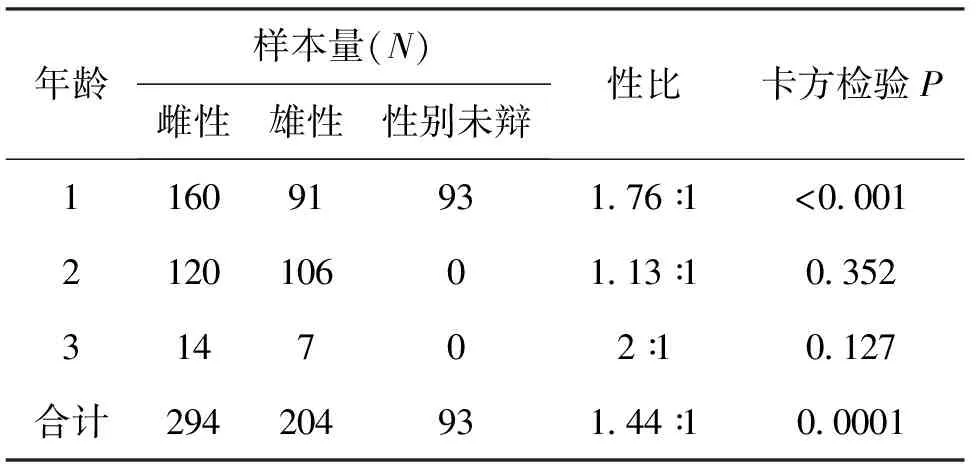
表1 贝氏 各年龄组性比Tab.1 Sex ratio among different age groups of H.bleekeri
各体长组性比变化见表2。体长6.9 cm以下均为性别未辨个体,7.0 cm以上体长组,除8.0~8.9 cm体长组雄性个体多于雌性外,其他体长组均为雌性多于雄性。检验显示,9.0~9.9 cm(χ2=5.081,P<0.05)以及10.0~10.9 cm体长组(χ2=22.617,P<0.01)雌雄比与1∶1有显著性差异,其他体长组则无显著性差异(P>0.05)。
各月性比存在明显差异(表3),χ2检验显示,4月份雌雄比6.43∶1,与1∶1存在极显著差异(P<0.001);8月份则雄性多于雌性,雌雄比0.46∶1,与1∶1存在显著性差异(P<0.05);其他各月性比与1∶1无显著性差异(P>0.05)。
2.2 初次性成熟大小
在何王庙故道采集到的样本中,最小观测性成熟个体体长、体重分别为:雌性7.5 cm,体重6.65 g;雄性8.2 cm,体重8.13 g。均为1龄鱼。
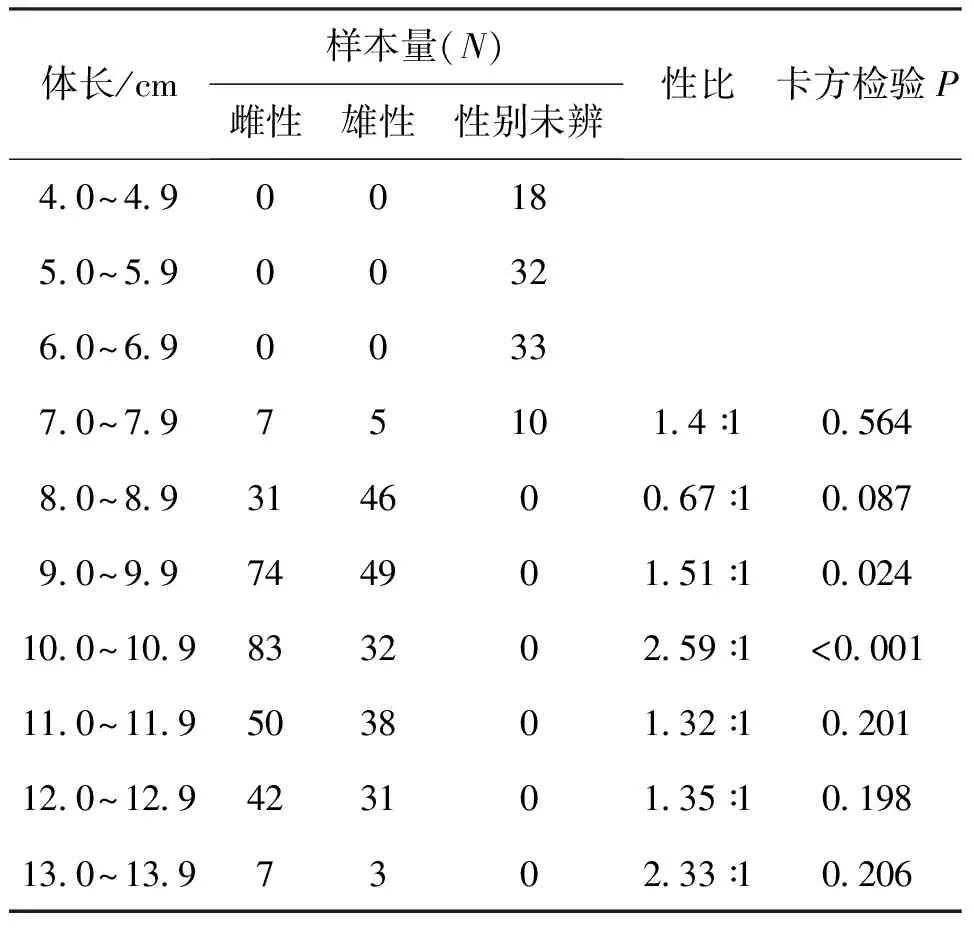
表2 贝氏 各体长组性比Tab.2 Sex ratio among different body length groups of H.bleekeri
用logistic方程拟合的贝氏 体长与性成熟的比例关系(如图1)如下:
从方程中可以得出,SL♀50=8.08 cm,SL♂50=8.47 cm,对应的雌、雄初次性成熟体重和年龄分别为7.76 g、0.8龄和7.63 g、0.9 龄。
2.3 繁殖期
图2为2016年贝氏 成熟群体成熟系数GSI(均值±SD)的变化图。4月采集到的贝氏 性成熟个体性腺绝大多数发育到Ⅳ期,少量处于Ⅴ期。雌性和雄性的GSI均达到峰值,雌性均值为11.26,雄性均值3.07。6月份仍发现有Ⅴ期性腺,但GSI有所下降,雌性GSI均值降为8.88,雄性GSI均值则降至0.83。8月份雌性GSI均值快速下降至2.46,雄性GSI均值下降至0.62,无Ⅳ和Ⅴ期卵巢,至9月下旬,雌性、雄性性腺100%为Ⅵ-Ⅱ期,雌性GSI均值下降至0.43,雄性GSI均值下降至0.46,说明繁殖在8月就已经结束。由此推测,贝氏 繁殖期主要为4-6月,可能延至7月份。

图1 体长和性成熟比例的logistics回归曲线Fig.1 The logistics curve fitted by standard length and percentage of sexual maturity of H.bleekeri

图2 贝氏 性成熟系数(GSI)变化Fig.2 Variation of gonadosomatic index (GSI) in H.bleekeri
2.4 产卵类型
4月和6月贝氏 Ⅳ期卵巢样本沉积卵黄的卵径频率分布如图3。 这两个月的卵径分布范围和平均卵径十分相似,4月卵径范围524.6~1 801.4 μm,均值935.3 μm;6月卵径范围508.2~1 812.1 μm,均值928.0 μm。虽然卵径分布频率呈“单峰型”,但沉积卵黄的卵径分布范围较大,由此推测贝氏 很有可能是分批产卵鱼类,而不是一次性产卵鱼类。
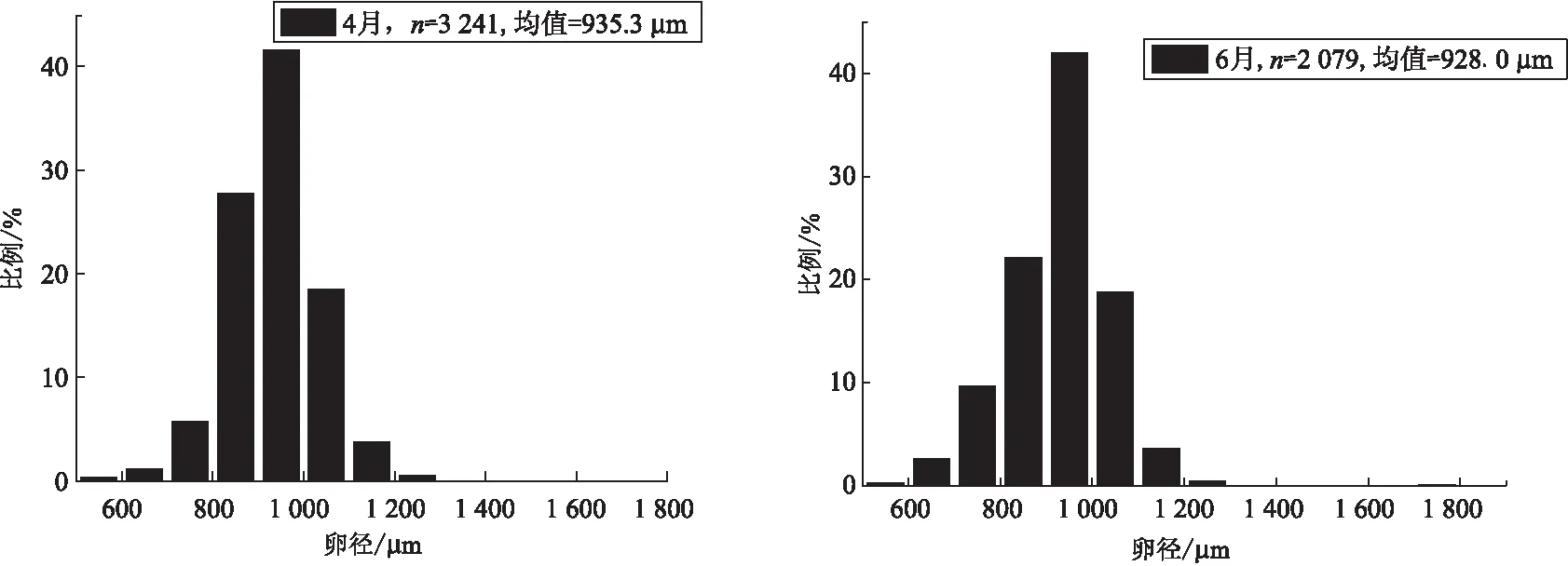
图3 贝氏 4月(左)和6月(右)Ⅳ 期卵巢卵径分布Fig.3 Distribution of oocytes diameter in stage Ⅳ ovaries of H.bleekeri sampled in April (left)and June (right)
2.5 产卵群体组成
贝氏 繁殖群体年龄结构简单,只有3个龄组,主要由1-2龄个体组成,其中1龄个体占51.8%,2龄个体占44.9%,3龄个体仅为3.3%。根据贝氏 的产卵群体(P)由补充群体(K)和剩余群体(D)两部分组成,且以初次性成熟的1龄补充群体为主的特点,认为贝氏 繁殖群体属于第Ⅱ类型(P=K+D,K>D)。
2.6 繁殖力
2.6.1 个体繁殖力及变动范围
随机取样的卵巢发育处于Ⅳ期的45尾贝氏 样本,体长范围为8.1-12.6 cm,体重范围为6.36~28.98 g,成熟系数(GSI)为9.07%~21.29%,性腺重(GW)为0.75~4.55 g。绝对繁殖力F为1 703~9 601粒/尾,平均为(5 876.6±1 837.8)粒/尾;相对繁殖力FW为204.7~406.5粒/g,平均(322.3±49.7)粒/g,相对繁殖力FL为212.0~827.6粒/cm,平均(552.4±136.1)粒/cm。
按体长分组(表4),F、FW均随体长的增加而增加,而FL、GSI随体长变化不明显。按年龄分组(表5),2龄组F、FL及FW均高于1龄组。

表4 贝氏 各体长组的繁殖力Tab.4 Fecundity in different standard length groups of H.bleekeri
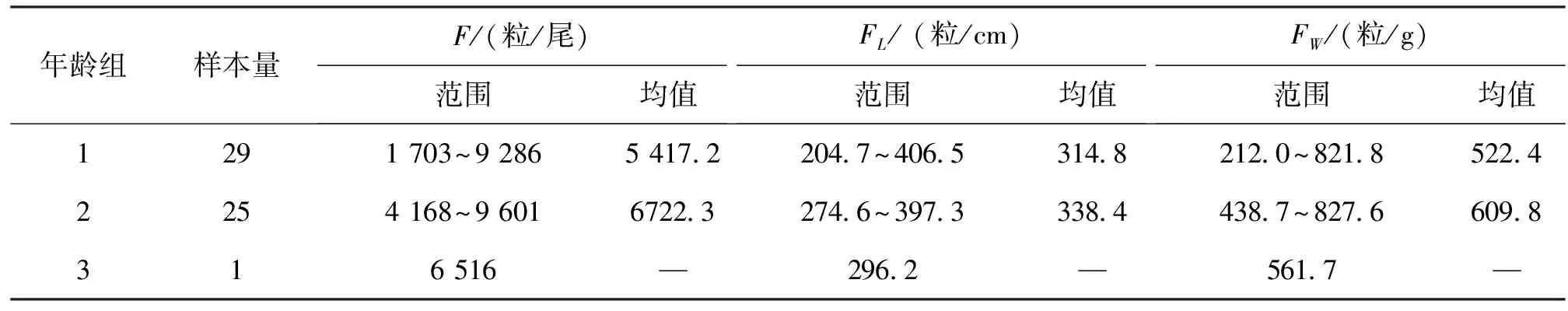
表5 贝氏 各年龄组的繁殖力Tab.5 Fecundity in different age group of H.bleekeri
2.6.2 个体繁殖力与生物学指标的关系
绝对繁殖力(F)与体重(W)、性腺重(GW)呈线性关系,与体长(SL)呈幂函数关系,而与成熟系数(GSI)不相关;单位体重相对繁殖力(FW)与成熟系数呈幂函数相关,与体长、体重和性腺重不相关;单位体长相对繁殖力(FL)与体重呈幂函数相关,与性腺重呈线性相关,而与体长、体重不相关。相关公式如下:
F=20.7SL2.397 8(R2=0.628 4,n=45)
F=164.8+312.56W(R2=0.775 1,n=45)
F=196.1+2 199.96GW(R2=0.916,n=45)
FW=51.84GSI0.689 1(R2=0.797 4,n=45)
FL=82.02W0.659 7(R2=0.604 1,n=45)
FL=123.01+163.41GW(R2=0.960 8,n=45)
3 讨论
3.1 性比
种群性比反映种群结构特点和变化,既有种的特性,又受环境因子的影响。而产卵群体的性比主要与鱼类繁殖习性相关[16]。本研究中,贝氏 总体雌雄性比为1.44∶1,与长江其他小型鱼类(表8),如半 (Hemiculterellasauvagi)[17](1.46∶1)、银 (Squalidusargentatus)[18](1.94∶1)、似鳊(Pseudobramasimoni)[19](2.12∶1)等相似,均为雌性多于雄性,而与 (Hemiculterleucisculus)(1.04∶1)[20]、张氏 (Hemiculterchangi)(1.07∶1)[21]群体雌雄数量相当不同。但与银鮈和似鳊有所不同的是,在4-6月繁殖季节,贝氏 产卵群体雌雄比2.5∶1,而银鮈和似鳊产卵群体性比均接近1∶1;贝氏 非繁殖季节雌雄比为0.91∶1,接近1∶1,而银鮈和似鳊非繁殖季节性比均是雌性显著多于雄性[20-22],恰恰相反。因为采样点在长江中游的监利何王庙故道,而并非在其繁殖发生所在的长江中游干流,所以,本实验研究的繁殖群体性比能不能代表在长江中游干流的实际繁殖群体,尚有待于进一步研究验证。
3.2 初次性成熟体长
初次性成熟大小是鱼类繁殖的属性之一,同一物种的初次性成熟大小和环境之间密切相关,因此了解它的变化状况有助于理解鱼类的环境适应性,也有助于资源的有效管理[23]。我国以往研究多用最小观测性成熟大小来衡量鱼类性成熟大小[9-10,17,20-21]。但van der Velde等[13]认为最小观测体长往往是种群特例,不能代表整个种群,因此用最小观测体长评估种群初次性成熟体长是不准确的,而用L50法估算的初次性成熟体长更能代表整个种群性成熟状况。因此建议在今后的研究中使用L50法估算小型鱼类的初次性成熟体长。
性成熟年龄和个体大小对一个物种来说具有稳定性[24],但会受到生长速度等因素影响。吴清江等[9]观察到的黑龙江流域贝氏 最小观测成熟体长,与李树国等[10]观察到的呼伦湖蒙古油 最小观测性成熟体长几乎一致,而从何王庙故道采集的长江中游贝氏 雌性、雄性初次性成熟体长比这两个北方水域的最小观测性成熟体长都要小(表6)。黑龙江流域和呼伦湖蒙古油 最小观测性成熟体长比长江流域贝氏 大,主要原因是贝氏 在北方水体中由于水温低,生长相对慢,性成熟相对较晚,因此初次性成熟个体较大;而长江流域水温高,生长快,性成熟早,1龄鱼均可达到性成熟,因此初次性成熟个体较小。
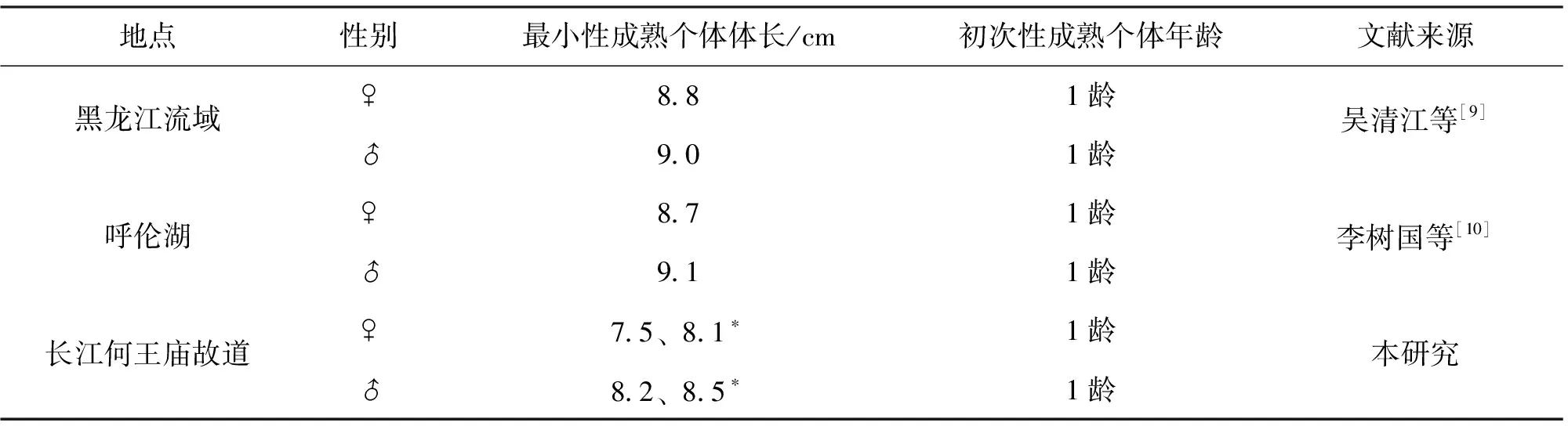
表6 不同地区贝氏 性成熟个体比较Tab.6 The minimum mature individuals of H.bleekeri in different waters
注: *上标表示初次性成熟体长(SL50)

表7 长江小型鱼类繁殖生物学特征比较Tab.7 Reproductive biological indices among different small-sized fishes in the Yangtzi River
3.3 繁殖期和繁殖习性
鱼类繁殖期一般总是在一年中能使它们繁殖后代的时间延至最长,并使它们的后代的早期发育获得最佳环境条件,特别是幼鱼是否存在丰富的开口食饵,以确保繁殖后代获得最大的成活率的时期,而温度、饵料等是影响鱼类繁殖时间的最主要的生态环境因子[16]。学者们认为依赖流水条件产卵的鱼类,其繁殖季节往往同所在水体每年涨水期一致,洪水可扩大繁殖栖息地、增加可利用饵料、避免种群拥挤并减少捕食压力[25-26]。而长江流域属于典型的亚热带季风气候,每年大部分降水集中在4-9月份,且这期间水温回升,光照时间延长,饵料生物大量繁殖[16]。因此长江流域绝大多数繁殖期集中在4-9月份,小型鱼类也是如此,如似鳊[19](5-8月份)、银[18](5-9月份)、[20](5-8月)和张氏 (5-9月)[21](表7)。贝氏 在长江干流的繁殖期,据刘媛媛[27]对长江口繁殖盛期稚鱼优势种的分析结果显示,6月份贝氏 稚鱼为优势种,由此可推测贝氏 在长江口繁殖高峰期在6月之前;而王红丽等[28]在三峡库区丰都江段调查时发现,6月后流速较高时段出现的卵汛中贝氏 的数量最多(占86.24%),由此推测贝氏 在三峡库区丰都江段繁殖期主要在6-7月。本研究中,根据成熟系数变化结合性腺发育期观察推测贝氏 在长江的繁殖期为4-6月,可能延至7月,与上述两者的观察结果基本一致。
鱼类繁殖期开始和持续时间长短常与鱼类的产卵类型相关,同时也受到水温、水文等环境因子的影响。根据贝氏 成熟卵巢中有卵黄沉积的卵径分布频率为单峰型的特点,判断可能为一次性产卵鱼类,但鉴于其卵径分布范围大(0.5~1.8 mm)的特点,推测长江中游的贝氏 可能为分批产卵鱼类,从繁殖期较长的实际情况判断,其为分批产卵类型的可能性更大,具体为何种产卵类型,需要对其卵巢发育进行连续的组织切片观察分析后,才能做出客观的判断。李树国等[10]认为呼伦湖的贝氏 是一次性产卵鱼类,繁殖季节6月中旬至7月下旬;黑龙江流域贝氏 尚不清楚是何种产卵类型,其在6月产卵[9]。相对北方水体而言,贝氏 在长江流域产卵较早,而且繁殖相对延长。从产卵性质来看,贝氏 在黑龙江和长江中均产漂流性卵[9,28],而在呼伦湖产粘性卵[10],与其适应长期生活的繁殖环境有关。与长江流域其他小型鱼类相比(表7),贝氏 和半[17]繁殖期相似,而比银[18]、似鳊[19]、[20]、张氏[21]稍早。与其他鱼类错开繁殖高峰期,可以减轻种间仔稚鱼对食物的竞争而提高后代成活率,有利于种群补充和发展。
3.4 繁殖力
繁殖力是估算鱼类种群繁殖潜力的基础,鱼类种群往往随着环境条件的改变而做出适应性变化,通常受到饵料丰度、水温和其他环境因素的影响[29-30]。
与呼伦湖贝氏 相比,长江贝氏 的绝对繁殖力F(1 703~9 601粒/尾)、相对繁殖力FL(212.0~827.6粒/cm)、相对繁殖力FW(204.7~406.5粒/cm),均低于呼伦湖贝氏 的F(2 021~15 470粒/尾)、FL(230~1 137粒/cm)、FW(273~700粒/g)[10]。长江中游贝氏 与呼伦湖贝氏 繁殖力的差异可能是不同种群在不同环境压力下的反应,是为了保障种群繁衍的一种繁殖策略。
鱼类在繁殖投入有限的情况下或采取减小卵径、提高繁殖力的繁殖策略,或采取增大卵径、降低繁殖力的繁殖策略。与长江流域其它小型鱼类如半[17]、[20]、张氏[21]、银[18]及似鳊[19]等相比(表7),贝氏 和半[17]采取的繁殖策略最为相似,两者成熟卵径较大,但繁殖力较低,而银[18]、似鳊[19]和[20]则采取高繁殖力、小卵径的繁殖对策。卵径较大,所带卵黄营养多、孵出鱼苗个体较大,有利于开口鱼苗摄食和早期生长,提高成活率[24]。
面对过度捕捞等压力,小型鱼类因其性成熟早、年龄结构简单、世代更新快而取代中大型鱼类成为天然水体中优势种类[31]。贝氏 性成熟早、个体小、繁殖群体年龄结构简单、繁殖期较长且与长江其它小型鱼类繁殖高峰期错开,虽然繁殖力相对稍低但成熟卵体积较大,卵径较大能在开口饵料和早期生长方面提高后代成活率,从而使其成为长江干流及其附属湖泊的主要优势种类。何王庙故道与长江相通,可以获得来自长江中游干流贝氏 等优势小型鱼类的种群补充,对保障长江江豚饵料鱼饵料资源十分有利。
参考文献:
[1]王斌梁,张 琪,王 玺,等.长江中游故道藻相特征及水质分析[J].环境科学与技术,2014,37(6):166-170.
[2]陈佩薰,刘沛霖,刘仁俊,等.长江中游(武汉—岳阳江段)豚类的分布、生态、行为和保护[J].海洋与湖沼,1980,11(1):73-84.
[3]于道平,蒋文华.半自然水域中长江江豚食性与摄食行为的初步观察[J].兽类学报,2003,23(3):200 -202.
[4]李德尚,吴雄飞,谢宗墉,等.杂鱼在养鱼水库中的作用及其种群控制[J].生态学报,1988,8(4):16-22.
[5]叶佳林.太湖梅梁湾沿岸带鱼类组成和摄食生态研究[D].武汉:华中农业大学,2006.
[6]陈校辉,边文冀,赵 钦,等.长江江苏段鱼类种类组成和优势种研究[J].长江流域资源环境,2007,16(5):571-577.
[7]李辉华,郭弘艺,唐文乔,等.长江下游靖江段沿岸贝氏 渔获量的时间格局及ARIMA模型预测[J].中国水产科学,2009,16 (3):357-364.
[8]毛志刚,谷孝鸿,曾庆飞.呼伦湖鱼类群落结构及其渔业资源变化[J].湖泊科学,2016,28(2):387-394.
[9]吴清江,易伯鲁. 条属鱼类和黑龙江流域 条属鱼类的初步生态调查[J].水生生物学报,1959(2):157-167.
[10]李树国,张全诚,高庆全,等.呼伦湖蒙古油 繁殖生物学的研究[J].淡水渔业,2008,38 (5):51-54.
[11]边书京.梁子湖五种小型鲤科鱼类食性和消化器官的初步研究[D].武汉:华中农业大学,1999.
[12]黄海水产研究所.海洋水产资源调查手册[M].上海:上海科学技术出版社,1981.
[13]van der Velde T D,Griffiths S P,Fry G C.Reproductive biology of the commercially and recreationally important cobiaRachycentroncanadumin northeastern Australia[J].Fish Sci,2010,76(76):33-43.
[14]王剑伟.稀有鮈鲫的繁殖生物学[J].水生生物学报,1992,16(2):165-174.
[15]Mackay I,Mann K H.Fecundity of two cyprinid fishes in the River Thames,Reading,England[J].Fish Res Bd Can,1969,26:2095-2105.
[16]殷名称.鱼类生态学[M].北京:中国农业出版社,1995.
[17]王 俊,刘 飞,张 雄,等.赤水河半鳖繁殖生物学研究[C].中国海洋湖沼学会,中国动物学会鱼类学分会·2012年学术研讨会.2012.
[18]王海生.长江天鹅洲故道银鮈的生物学研究[D].武汉:华中农业大学,2013.
[19]孙广文.长江天鹅洲故道似鳊的年龄、生长、死亡率和繁殖[D].武汉:华中农业大学,2013.
[20]杨泽均,何晓红,赖朝文,等. 的性状、年龄与生长及性腺发育[J].四川动物,1992,11(1):42-44.
[21]孙宝柱,李 晋,但胜国,等.张氏 的繁殖生物学特性[J].水生生物学报,2010,34(5):998-1003.
[22]李修峰,黄道明,谢文星,等.汉江中游产漂流性卵鱼类产卵场的现状[J].大连海洋大学学报,2006,21(2):105-111.
[23]沈建忠.中华鳑鮍Rhodeussinensis繁殖习性的初步观察[J].华中农业大学学报,2000,19(5):494-496.
[24]Wootton R J.Ecology of teleost fishes[M].London and New York:Chapman & Hall,1990.
[25]Welcomme R L.Fisheries ecology of flood plain rivers[M].London and New York:Longman.1979.
[26]Humphries P,King A J,Koehn J D.Fish,flows and flood plains:Links between freshwater fishes and their environment in the Murray-Darling River System,Australia[J].Environ Biol Fish,1999,56(1):129-151.
[27]刘媛媛.长江口鱼类繁殖盛期仔稚鱼种类组成及优势种分布研究[D].上海:上海海洋大学,2016.
[28]王红丽,黎明政,高 欣,等.三峡库区丰都江段鱼类早期资源现状[J].水生生物学报,2015,39(5):954-964.
[29]Lepage C,Cury P.Population viability and spatial fish reproductive strategies in constant and changing environments:an individual-based modelling approach[J].Can J Fish Aquat Sci,2011,54(10):2235-2246.
[30]Hossain M Y,Rahman M M,Miranda R,et al.Size at first sexual maturity,fecundity,length-weight and length-length relationships ofPuntiussophore(Cyprinidae) in Bangladeshi waters[J].J Appl Ichthyol,2012,28(5):818-822.
[31]刘其根,沈建忠,陈马康,等.天然经济鱼类小型化问题的研究进展[J].上海海洋大学学报,2005,14(1):79-83.
