广西野生鲤与建鲤的遗传多样性分析
2018-05-17陈子桂王培培覃俊奇袁宗伟曹寿雄
陈子桂,王培培,覃俊奇,廖 愚,袁宗伟,曹寿雄
(广西壮族自治区水产引育种中心,南宁 530031)
鲤(Cyprinuscarpio)属鲤形目鲤科鲤属,鲤是广西水产养殖的重要产业。上个世纪90年代,广西引进建鲤品种,由于经过广西当地多代的养殖,一些繁育场忽略保种,没有进行适当的选育或提纯复壮,加上近年来受生态破坏、水质污染等因素影响,鲤种群的种质资源严重受损,遗传多样性丧失,种质严重退化,表现为生长速度慢,抗逆抗病性差,性成熟过早等经济性状退化[1-3]。因此,进一步了解广西封闭水体鲤遗传结构,合理开发利用和保护广西鲤种质资源,对建鲤的继续选育和种质提纯工作具有十分重要的意义。
微卫星分子标记(SSR)因共显性具有遗传、可重复性好、高度多态性的特点,在鱼类种质分析中已经得到广泛应用。Reid等[4]利用SSR分析了虹鳟群体的遗传多样性,Wang等[5]对亚州海鲈进行全同胞子代种质鉴定,确定QTL的数量和位置,确定负责生长相关性状的个体基因,鲁翠云等[6]利用SSR技术进行镜鲤家系构建,进一步避免了家系间的近交,孙效文等[7]对镜鲤的遗传结构分析并确定了与性状相关的等位基因,邢新梅等[8]对镜鲤的体形性状的关联性进行分析,其它研究人员对鲤遗传结构也做了诸多研究[9-14]。
该研究利用微卫星分子标记技术,对相对封闭水域的广西本地野生鲤和引进广西多年的建鲤进行遗传结构分析研究,以期把广西本地野生鲤和当地建鲤优良性状聚合,为合理开发利用广西野生鲤种质资源和对建鲤的继续选育提供数据支持。
1 材料与方法
1.1 实验材料
该研究所用的试验材料由广西水产引育种中心2015年从广西天等县较封闭的水域收集的本地野生鲤群体和2004年从中国水产科学研究院淡水渔业研究中心引进的并经过第三代群体选育的建鲤,实验材料于2016年2月分别从以上两个群体进行采样,每个群体取30尾鱼作为样本。
1.2 实验方法
(1)选取广西本地鲤 (H1号)和建鲤(H2号)各30尾鱼,抽取尾部静脉血样30 μL/尾,置于1 mL的EP管加入ACD抗凝溶液摇匀,血液基因组DNA提取试剂盒抽提DNA,纯化后的DNA保存液用琼脂糖凝胶电泳测定完整性,稀释至50 ng/μL备用。微卫星引物为中国水产科学研究院黑龙江水产研究所开发的镜鲤微卫星标记的引物,详见表1。(2)使用TAKARA Mighty Amp®DNA Polymerase Ver.2 (R071Q)进行PCR扩增,反应体系如下:25 μL 2×Mighty Amp Buffer Ver.2、1 μL PrimerF 10 μmol/L、1μL Primer R 10 μmol/L、1 μL Mighty Amp、DNA Polymerase、1 μL DNA、21 μL ddH2O。PCR反应程序为 94 ℃预变性4 min,94 ℃变性30 s,退火30 s,72 ℃延伸30 s,30个循环,最后72 ℃延伸5 min。(3) PCR扩增产物经8.0%非变性聚丙烯酰胺凝胶电泳后,硝酸银染色,用混合显色液显色,系统成像仪拍照。(4)根据50 bp DNA Step ladder分子标记的清晰条带和鲤PCR扩增片段大小,分析各位点的等位基因。
1.3 数据处理
由群体遗传学分析软件Popgene 1.32分析数据,分析计算等位基因数(A),有效等位基因数(Ne),期望杂合度(He),平均多态信息含量(PIC)等信息,用populations计算群体间遗传距离(genetic distanceD)。

表1 10对引物的序列和退火温度Tab.1 Sequences and annealing temperatures of 10 pairs of primers
2 结果与分析
实验结果中10对微卫星的电泳图全部显示出了清晰的条带,从表2分析表明,10个位点均检测出了等位基因,有效等位基因在1.167 5~1.520 3,平均为1.360 2;等位基因控制着数量性状,数量多少决定遗传多态性高低,等位基因的丰度越高,群体的生产性状越具有多样性,因而能更多适应生态自然环境的自然选择。观测杂合度在0.397~0.762之间,平均为0.562 6,期望杂合度0.301~0.629之间,平均0.463 3,说明它们的遗传变异度高,遗传多样性丰富,多态性信息含量(PIC)在0.289 7~0.685 0之间,平均多态信息含量0.438 8,为中度多态性。表明建鲤在广西经多年人工群选,由于经过一定的人为选择性,建鲤群体的杂合度比广西本地野生鲤群体降低了,建鲤的遗传多样性数值低于广西本地鲤,建鲤经过广西本地多年的适应性养殖,其性状趋于稳定,遗传多样性降低,可进一步选育的空间有所下降。

表2 微卫星各位点及两个群体的遗传参数Tab.2 Microsatellite loci and genetic parameters of two populations
注:A为平均等位基因;Ne为有效等位基因;Ho为观测杂合度值;He为期望杂合度值;PIC为多态信息含量
多态信息含量是检验群体遗传多样性的指标[15],本地野生鲤的有效等位基因数1.562,大于建鲤的1.388,多态信息含量0.685,大于建鲤的0.424,广西本地野生鲤的遗传多样性高于建鲤,建鲤经过多年的群体选育,由于人为群体选择,遗传多样性低于野生鲤,而本地野生鲤由于生存在相对封闭的水域,即使与早先引进的建鲤存在一定的基因交流,由于没有受到人工选择的干扰,也能保持较高的遗传多样性,结果详见表2。
群体间遗传分化系数(GST)被认为是度量群体间基因分化相对大小的一个较好指标[16],该研究通过分析H1号和H2号群体10个位点的基因分化系数(GST)和基因流(Nm),存在显著遗传分化的位点有3个,极显著遗传分化的位点有1个,H1号和H2号群体平均基因分化系数为0.106 9,表明有10.69%的遗传分化来自两个不同群体之间,89.31%的遗传分化来自整个鲤内部个体之间,基因流平均为6.216 9(表3)。表明2个鲤群体间之间有明显的遗传分化,基因流Nm和基因分化系数GST是负相关的。
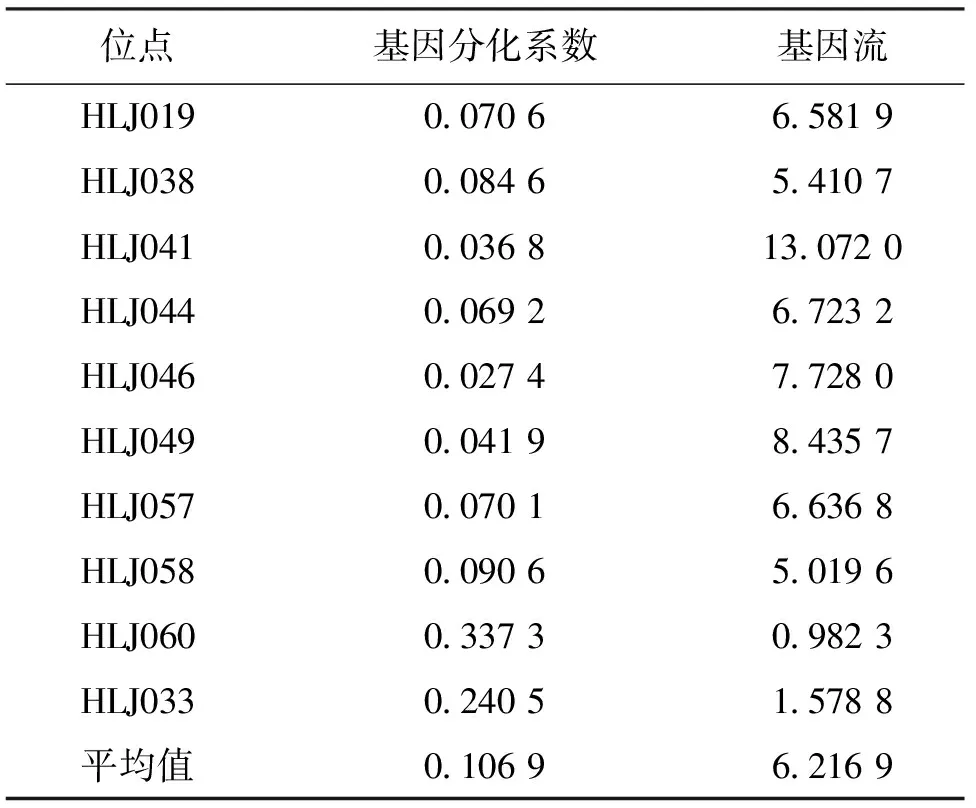
表3 10个微卫星位点的基因分化系数GST和基因流NmTab.3 Gene differentiation coefficient (GST) and gene flow (Nm) of 10 microsatellite loci
从实验结果来分析,广西本地鲤与建鲤的遗传距离为0.146 3,相似性系数为0.828 4(表4)。表明广西本地野生鲤与人工选育品种建鲤之间存在一定的遗传异质性,在选择育种过程中,为提高选择效应,获得最大的遗传增益,在建立选育基础群体的时候可以进行优良性状聚合。

表4 两个鲤群体的遗传距离和遗传相似系数Tab.4 Genetic distance and genetic similarity coefficient of two populations
注:0.146 3为遗传距离;0.828 4为遗传相似性系数
3 讨论
3.1 鲤种群遗传多样性
等位基因是位于一对同源染色体的相同位置上控制同一性状的不同形态的基因,数量越多,表明遗传多态性高,控制不同性状的基因座位越丰富,该试验中平均等位基因数5.3个,有效等位基因数平均为1.360 2,观测杂合度平均为0.562 6,期望杂合度平均0.463 3,观测杂合度较为接近期望杂合度,表明控制相对性状的等位基因在两个鲤群体均匀分布,两个群体的基因频率符合哈迪-温伯格规律,群体间保持了足够的有效群体数量。多态信息含量(PIC)是DNA变异程度高低的指标,根据Botstein等[17]提出的衡量标准:0.25 遗传距离通常被用来衡量群体间遗传关系,反映不同群体间的血缘关系的远近,该研究根据Nei等[19]的方法对广西本地野生鲤与建鲤的亲缘关系进行分析,根据Thorp在1982年研究提出同种群体间遗传在0.03~0.2之间,遗传距离数值越大,表明鲤两个群体控制相同性状的基因频率相差较大,来自相同祖先的机率越小,亲缘关系就越远。该实验中广西本地野生鲤与建鲤的遗传距离为0.146 3,相似性系数为0.828 4,研究结果表明,虽然建鲤已在广西推广养殖多年,但由于该研究在广西相对封闭的水域取样,与其它开放性河流不同,野生鲤与建鲤的基因交流相对较少,两者具有明显的遗传异质性,亲缘关系相对较远。 由表3结果表明两个鲤群体基因分化系数在0~1之间,平均为0.106 9,为中度遗传分化水平,等位基因频率在两个鲤群体中分布有所差异,既有起源相同的遗传性状,又具有明显遗传分化,由于取样的地理局限性,无法完全排除两个群体之间的基因交流,基因流大于1,部分性状趋于相同,但并不一定由基因交流影响,广西西南地区斑块化水生环境,选择压力可能起了主导作用。 参考文献: [1]杨宁生,葛常水.我国水产种种质资源信息系统建设[J].中国农业科技导报,2003,5(3):47-51. [2]檀学文,杜专雄.我国渔业可持续发展问题研究[J].经济研究参考,2006,35:42-49. [3]乔德亮.水产动物种质资源保护策略和措施[J].当代水产,2004,29(2):39-40. [4]Reid D P,Szanto A,Glebe B,et a1.QTL for body weight and conditionfactorin Atlantic salmon (Salmosalar):comparative analysis with rainbow trout (Oncorhynchusmykiss) and Arctic eharr (Salvelinusalpinus)[J].Heredity,2004,94(2):166-172. [5]Wang C M,Lo L C,Zhu Z Y,et a1.A genome scan for quantitative trait loci affecting growth-related traits in an F1 family of Asian seabass (Lates calcarifer)[J].BMC Genom,2006,7(1):274-278. [6]鲁翠云,曹顶臣,孙效文,等.微卫星分子标记辅助镜鲤家系构建[J].中国水产科学,2008,15(6):893-901. [7]孙效文,鲁翠云,匡友谊.镜鲤两个繁殖群体的遗传结构和几种性状的基因型分析[J].水产学报,2007,31(3):273-279. [8]邢新梅,张 研,徐 鹏,等.镜鲤微卫星标记与体形性状的关联分析[J].中国水产科学,2011,18(5):965-982. [9]贾志虎,匡友谊,×××等.鲫微卫星标记与几个生长性状的相关性分析[J].水产学杂志,2012,25(2):1-6. [10]郑先虎,匡友谊,孙效文,等.镜鲤体长、体高、体厚性状QTL定位分析[J].遗传,2011,33(12):1366-1373. [11]顾 颖,李 超,鲁翠云.建鲤微卫星DNA亲权鉴定[J].遗传,2012,34(11):1447-1455. [12]马 磊,张晓峰,张天奇,等.微卫星标记与镜鲤部分生长性状的相关分析[J].湖南农业大学学报(自然科学版),2010,36(4):453-458. [13]张义凤,张 研,鲁翠云,等.鲤鱼微卫星标记与体重、体长和体高性状的相关分析[J].遗传,2008,30(5):613-519. [14]Sun X W,Liang L Q.A genetic linkage map of common carp and mapping of a locus associated with cold tolerance[J].Aquaculture,2004,238(4):165-172. [15]张 研,梁立群,常玉梅,等.鲤鱼体长性状的QTL定位及其遗传效应分析[J].遗传,2007,29(10):1243-1248. [16]陈文华,李建林,徐 跑,等.五个家系吉富罗非鱼的遗传多样性分析[J].生物技术通报,2009,(8):83-87. [17]Botstein D,White R L,Skolnick M,et al.Construction of a genetic linkage map in man using restriction fragment length polymorphism[J].Amer J Human Gen,1980,32:314-331. [18]关建义,张 芹,屈长义,等.野生和人工选育黄河鲤遗传多样性的ISRR[J].河南师范大学学报(自然版),2010,38(4):128-131. [19]Nei M,Maruyama T,Chakraborty R.The bottleneck effect and genetic variability in populations[J].Evolution,1975,29:1-10.3.2 亲缘关系与遗传分化
