长江流域10个不同水域克氏原螯虾野生群体形态差异分析
2018-05-17石林林梁宏伟李青彬李艳和
张 龙,石林林,梁宏伟,李青彬,李艳和
(1.华中农业大学水产学院,农业动物遗传育种与繁殖教育部重点实验室,水产养殖国家级实验教学示范中心,武汉 430070;2.中国水产科学研究院长江水产研究所,武汉430223)
克氏原螯虾(Procambarusclarkii),俗名小龙虾,隶属十足目(Decapoda)螯虾亚目(Astacidea)蝲蛄科(Cambaridae)原螯虾属(Procambarus)[1],因其虾肉营养价值高,味道鲜美,氨基酸种类多,蛋白质含量高及脂肪含量低[2],受到了众多消费者的喜爱[3]。克氏原螯虾对环境适应性强,耐低氧、病害少,能够生长在湖泊、沟渠、沼泽和稻田等其它水域。因此,无论是在我国南方还是北方,克氏原螯虾均可以生存[4,5]。克氏原螯虾在近几十年的繁衍和迁徙之中,受到当地气候、水环境及其他因素的影响可能已经在一些形态上产生了差异。对克氏原螯虾形态特征的研究能够快速、经济、直观地反映不同群体的形态差异,但这种方法一般适用于种间的系统分类,对种内不同群体不能有效鉴别,采用多元统计分析能够有效克服这个缺点[6]。多元回归分析常用于研究自变量与因变量之间潜在的本质关系,现已广泛应用于许多水生动物形态特征与其体重关系、群体分布以及雌雄鉴别模型等研究中[7]。本研究对来自长江上游、长江中游、长江下游以及靠近长江下游的洪泽湖和高邮湖的10个克氏原螯虾野生群体的形态运用3种多元统计方法以及T检验进行差异分析。通过对这些差异的分析,以期为克氏原螯虾的群体鉴别、种质资源以及繁殖育种等研究提供基础生物学资料。
1 材料与方法
1.1 实验材料
用于本研究的野生克氏原螯虾来自于长江流域的10个不同自然水体,样品于2016年4-8月采集,样本采集信息见表1。
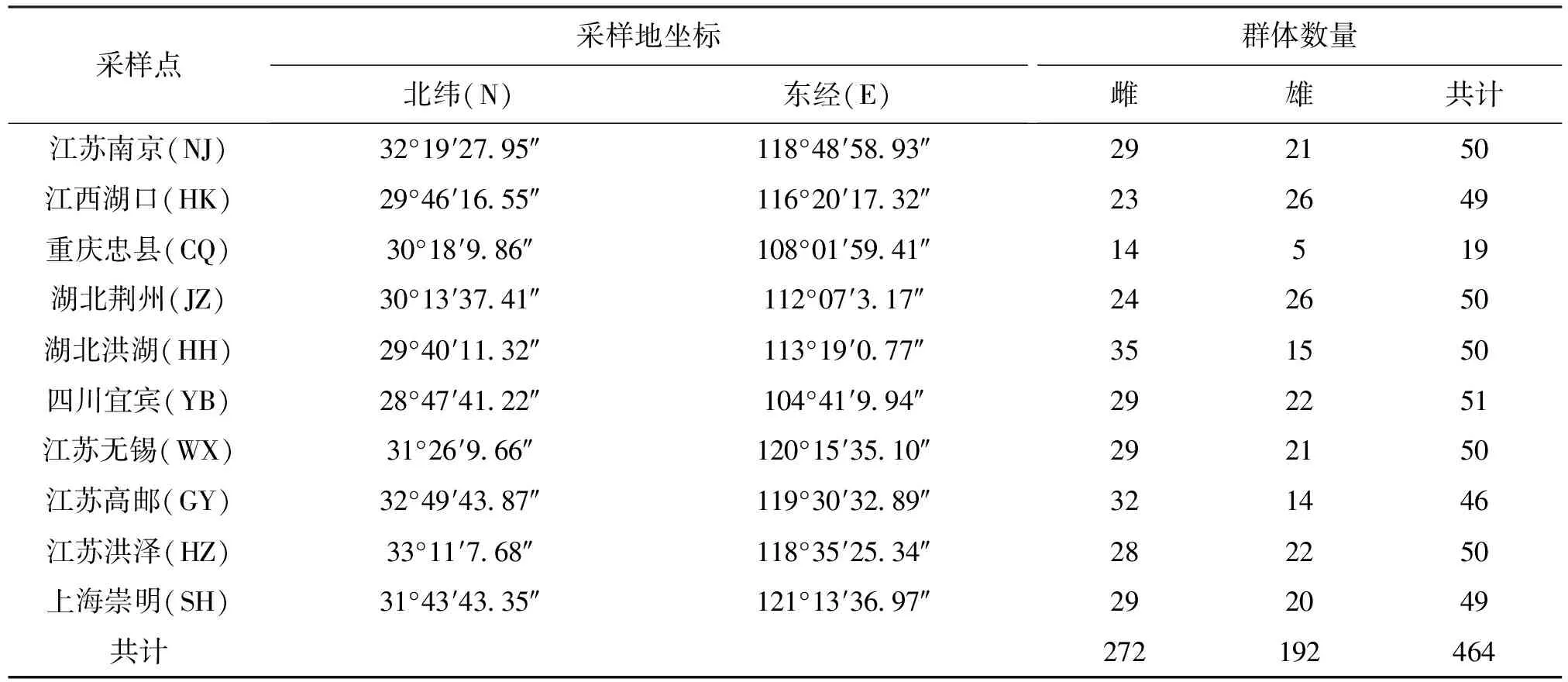
表1 采样信息表Tab.1 Sampling information
1.2 形态参数测量与数据处理
参照王克行[8]的测量方法,用游标卡尺测量每尾虾的全长、头胸甲长、头胸甲宽、第一腹节长、第一腹节宽和腹节总长(图1),精确到0.1 mm;用天平称取体质量,精确到0.01 g。数据分析采用SPSS 22.0软件包处理,运用聚类分析、主成分分析、判别分析和T检验对所得实验数据进行分析。为消除虾体大小对参数值的影响,多元分析中采用以全长为基数或取两个形态参数的比值。本研究共选取了13项形态比例参数,具体见表2。

图1 克氏原螯虾各形态测量指标Fig.1 The morphological measurement indexes of P.clarkiia.全长;b.头胸甲长;c.腹节总长;d.头胸甲宽;e.第一腹节长;f.第一腹节宽

表2 13项克氏原螯虾形态特征比例参数Tab.2 13 Morphological characteristics of P.clarkii
注:a.全长;b.头胸甲长;c.腹节总长;d.头胸甲宽;e.第一腹节长;f.第一腹节宽;W.体重
1.3 分析方法
1.3.1 聚类分析
首先运用EXCEL分别求出各个群体13项形态指标的平均值,然后采用欧式系统聚类法对这些平均值进行聚类分析。
1.3.2 主成分分析
主成分分析是一种将多个指标做小数指标的统计方法。先对数据进行校正处理,然后用SPSS 22.0软件对10个不同水域克氏原螯虾野生群体的形态参数进行主成分分析,得出各个成分的特征值、贡献率以及主成分载荷值。
1.3.3 判别分析
运用逐步判别分析法进行分析,对判别贡献率较大的指标建立判别方程,判别准确率的计算参照李思发等[9]的方法,计算公式如下:
判别准确率=某克氏原螯虾群体判别正确的尾数/该群体的实际尾数×100%

式中:Ai为第i个群体中判别正确的尾数,Bi为第i个群体的实际尾数,k为群体数。
1.3.4 差异系数(CV)检验
按如下参考公式进行:
CV=(M1-M2)/(S1+S2)
式中:M1和M2表示2个群体某指标的平均值,S1和S2为标准差。若CV值小于1.28,则表示2个群体在这项指标上的差异未达到亚种水平。
1.3.5 T检验
将10个群体的克氏原螯虾分成雌雄两组,然后对两组克氏原螯虾的13项形态指标进行T检验,判断雌雄在某性状指标的差异程度,若P>0.05则表示差异不显著,反之则视为差异显著。
2 结果与分析
2.1 聚类分析
对10个不同自然水域野生克氏原螯虾的13项形态指标平均值的聚类分析结果显示雌雄克氏原螯虾均分为2类(图2,图3),均为重庆忠县、湖北洪湖、江西湖口、湖北荆州聚为一类,其它群体聚为一类。在雌性群体中,江苏高邮和江苏洪泽欧式距离最短,首先聚为一支,后再与上海崇明、江苏南京、四川宜宾和江苏无锡聚为一支;雄性群体中,同样也是江苏高邮和江苏洪泽首先聚为一支,再同江苏南京、四川宜宾、上海崇明和江苏无锡聚为一支。

图2 10个不同水域克氏原螯虾雌性群体聚类分析Fig.2 Cluster analysis of P.clarkii female populations from 10 different watersheds
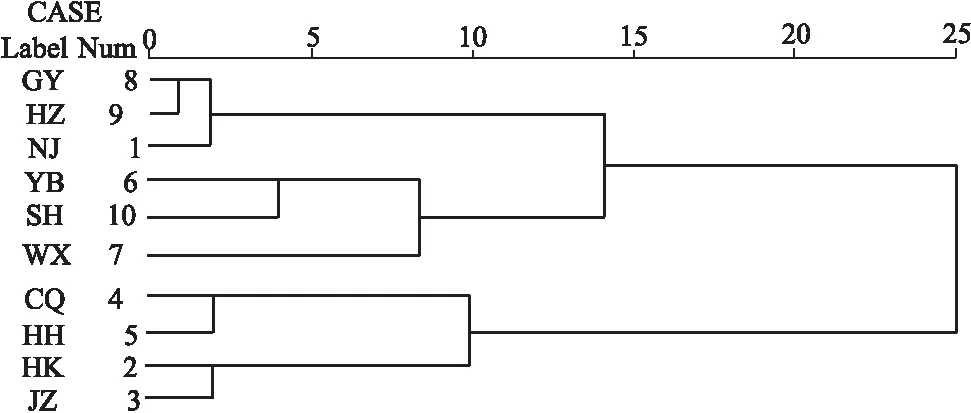
图3 10个不同水域克氏原螯虾雄性群体聚类分析Fig.3 Cluster analysis of P.clarkii male populations from 10 different watersheds
2.2 主成分分析
对10个不同水域克氏原螯虾野生群体进行主成分分析,选取了前四个主成分用于分析,累积贡献率为84.003%。其中主成分1为28.393%、主成分2为23.968%、主成分3为18.981%、主成分4为12.661%,较好地反映了克氏原螯虾群体的形态特征。第一主成分的特征根为3.691,方差贡献率为28.393%,反映了克氏原螯虾腹节总长的长度特征,称为腹长因子;第二主成分的特征根为3.116,方差贡献率为23.968%,反映了克氏原螯虾第一腹节的长度特征;第三主成分主要反映的是腹节宽度的特征,称为腹宽因子;第四主成分主要反映了克氏原螯虾头胸甲的特征,称为头胸甲因子。在10个群体中,江苏高邮群体第一主成分贡献率最高(39.776%),最低为上海崇明群体(30.127%);4个主成分累积贡献率最高的是重庆忠县群体(91.736%),最低的是上海崇明群体(84.997%)(表3)。

表3 10个不同水域克氏原螯虾群体的特征值和累积贡献率Tab.3 Eigen values and cumulative contribution rates of P.clarkii from 10 different watersheds
2.3 判别分析
通过逐步判别分析方法,从13项性状比值中筛选出对于区分10个不同水域克氏原螯虾雌性群体有显著贡献的5个变量X1、X2、X4、X7和X11。当雌性群体判别函数有这5个变量时,显著性检验结果Wilks’λ=0.102,P<0.000 1,说明利用这5个变量建立的雌性群体判别函数的判别效果良好,具有极显著的意义。对于雄性群体则筛选出了4个变量X1、X4、X9和X11,当雄性群体判别函数有这4个变量时,显著性检验结果Wilks’λ=0.114,P<0.000 1,说明利用这4个变量建立的雄性群体判别效果良好,具有极显著的意义。利用建立的判别方程式(略)对10个不同水域克氏原螯虾雌雄群体进行判别。将每尾雌性个体的5项性状的数值和每尾雄性个体的4项性状的数值分别带入雌雄群体的判别方程,函数值最大的判别方程所对应的群体就是该个体的所属群体。雌性群体中,判别率最高的是南京群体(93.1%),最低的是洪泽群体(32.1%),综合判别率为64.7%;雄性群体中,判别率最高的是宜宾群体(100%),最低的是高邮群体(7.1%),综合判别率为65.6%(表4)。

表4 克氏原螯虾雌雄群体的判别结果Tab.4 Discrimination results for male and female P.clarkii populations %

续表4
2.4 T检验
运用T检验对所采集的雌雄克氏原螯虾(雌性272尾,雄性192尾)的13项形态参数进行分析,结果表明雌雄克氏原螯虾在X1、X2和X9这3个形态参数上有显著性差异(P<0.05),在X3、X4和X13这3个形态参数上的差异达到了极显著(P<0.01)(表5)。从雌雄各形态参数的平均值看,6个有显著性差异的参数中,雄性X1、X2、X3和X9的平均值比雌性的大;而雌性的X4和X13这两个形态的平均值比雄性大。因此雌雄之间的形态学差异主要在体重、头胸甲长、头胸甲宽、第一腹节长和腹节总长上,且雄性体重、头胸甲长和头胸甲宽的平均值比雌性大,雌性的腹节总长平均值比雄性的大。
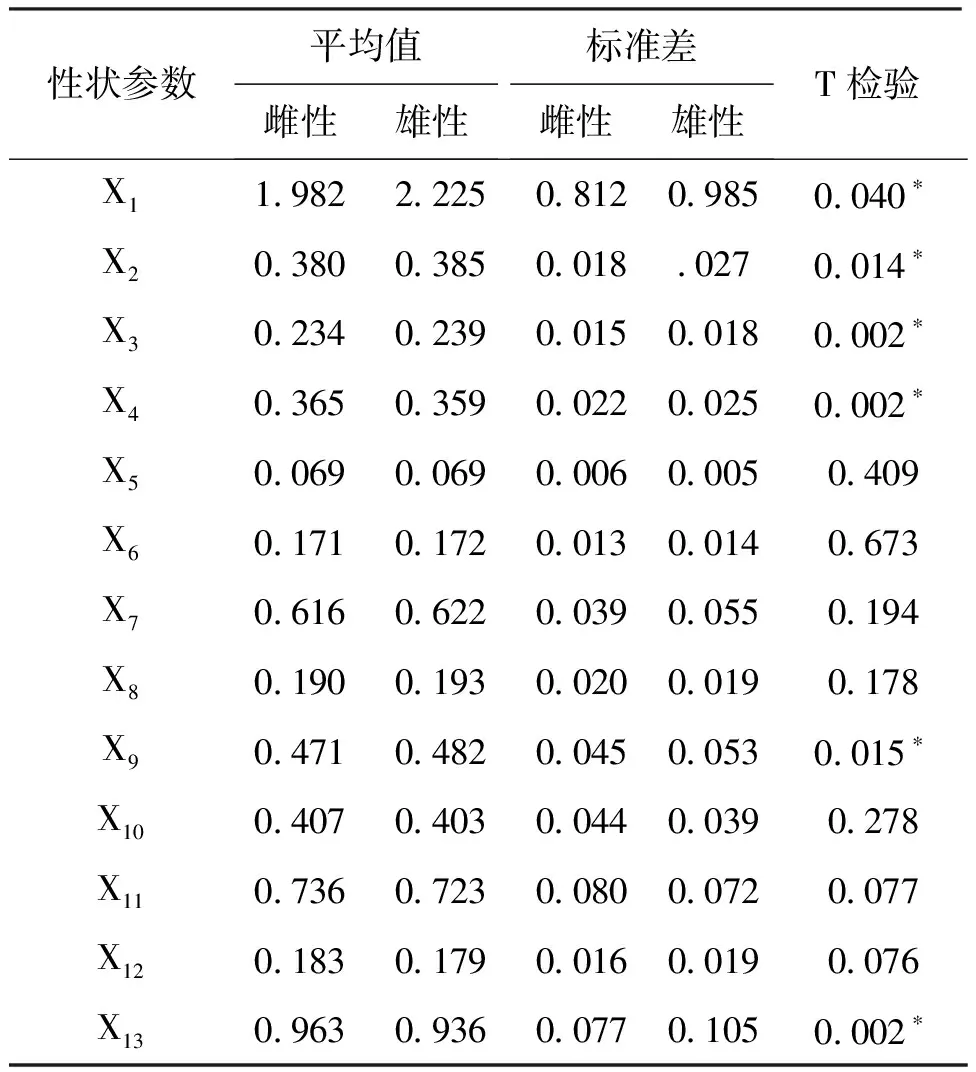
表5 雌雄两组群体的13项形态特征比例参数统计表Tab.5 Differences in the proportional parameters of 13 morphological characteristics between male and male group
注:*表示P<0.050,存在显著性差异。
2.5 差异系数(CV)检验分析
差异系数检验的结果(表6)显示,在13项形态指标中,有6项指标的差异系数值大于临界值1.28,但尚未占到13项指标的绝大多数。所以,10个群体克氏原螯虾群体间形态差异并未达到亚种以上分类水平。
3 讨论
3.1 克氏原螯虾不同群体间的形态差异分析
聚类分析可对不同群体进行初步分类,聚类的距离可以反映不同群体亲缘关系的远近[10]。本研究聚类分析结果显示10个不同水域克氏原螯虾的雌雄群体均为江苏高邮与江苏洪泽最先聚为一支,说明这两个群体的亲缘关系最近;重庆忠县、江西湖口、湖北荆州和湖北洪湖的雌雄群体与其它群体最后才聚为一支,表明这几个位于长江中上游地区的克氏原螯虾群体与江苏南京、江苏无锡和上海崇明等处于长江下游地区的克氏原螯虾群体亲缘关系较远。韩晓磊等[10]对不同地区克氏原螯虾形态差异分析表明江苏群体与湖北群体形态差异较远;郑友等[11]对6个水域克氏原螯虾形态差异分析结果显示鄱阳湖群体与洪泽湖群体亲缘关系较远,2个群体存在显著差异,其聚类分析结果均与本研究结果一致。值得注意的是,本次聚类分析显示四川宜宾的雌雄群体均与江苏无锡、上海崇明等长江下游水域聚为一类,这个结果与地理分布不相符合。克氏原螯虾已成为我国重要的水产养殖对象,长江中下游地区是克氏原螯虾人工养殖的主产区,苗种及成虾可能会因为销售等因素而存在于地理上较远的水域[12]。宜宾市河流多、密度大、水量丰富,共有大小溪流600多条,但因其河流多源于崇山峻岭,故水流湍急,加之长江所建水电站较多,因此,笔者推测克氏原螯虾虽迁徙能力较强,但也难以从长江末段的无锡、上海等水域迁徙到宜宾。并且近年来随着克氏原螯虾市场的火爆,克氏原螯虾养殖面积不断扩大,这让长江中下游地区的苗种和成虾通过销售等其它非自然因素方式进入宜宾成为可能。此外,宜宾市的河流纵横交错形成网状,易于克氏原螯虾在宜宾各河流分支的扩散。
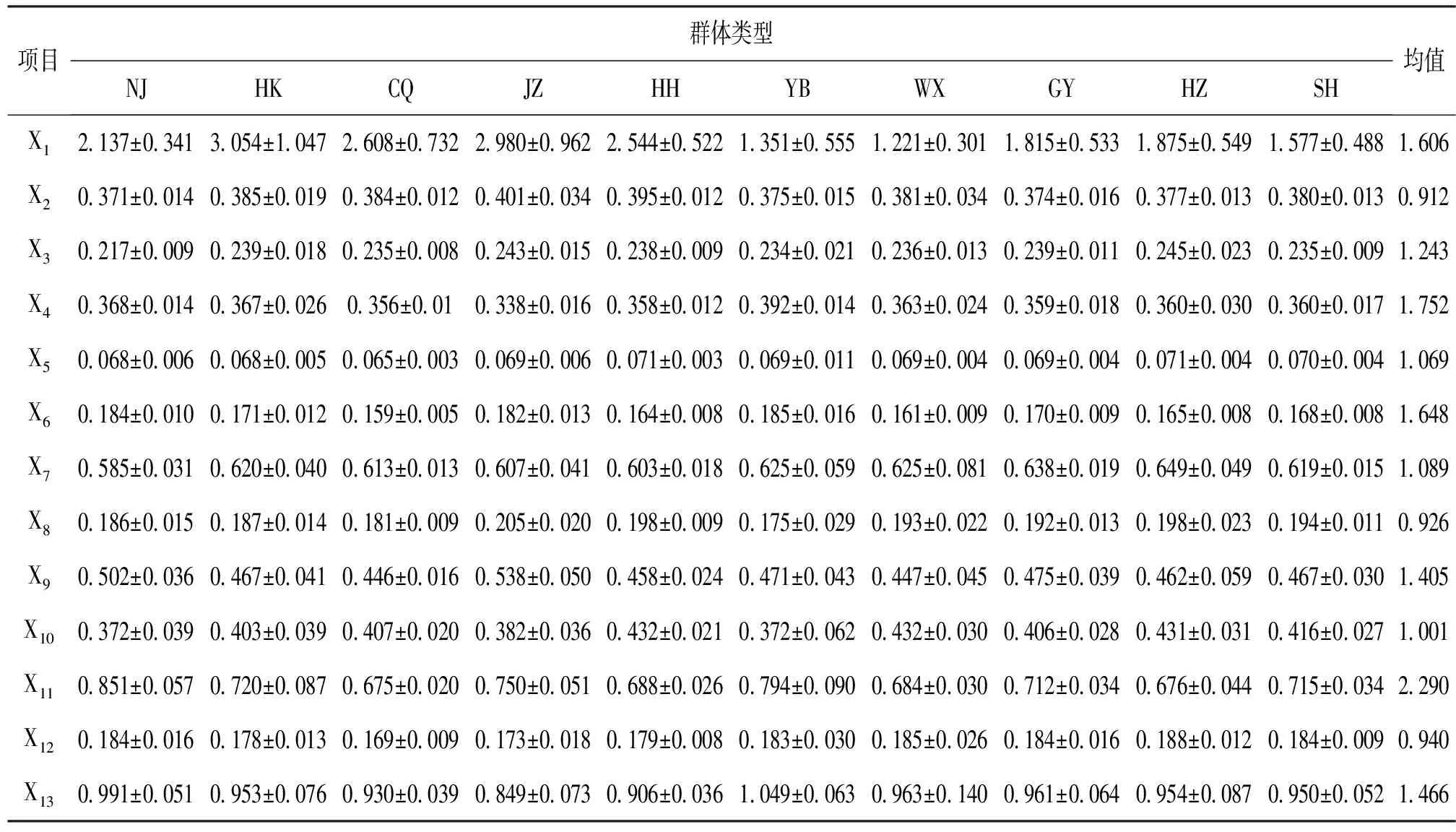
表6 克氏原螯虾群体间差异系数Tab.6 The coefficient of variation among P.clarkii populations
注:差异系数是指差别最大的2个群体间的CV值。
主成分分析能够将多个变量通过线性变换成几个少数主成分变量,并且要求这几种主要变量的相关性要尽可能低,是一种将少数主要变量来代替原来多变量的方法[13]。该方法现已经广泛应用于水产动物的形态分析,并取得了良好的效果[14]。韩晓磊等[15]对克氏原螯虾和东北螯虾的形态学特征进行主成分分析,结果表明第一主成分反映了克氏原螯虾和东北螯虾身体宽度的形态变化;安丽等[16]对67尾翘嘴红鲌的10项比例性状进行主成分分析后,将翘嘴红鲌的形态指标归纳为“肥瘦特征”、“躯干特征”、“眼间距特征”、“尾部特征”和“头部特征”,较为完整地描述了翘嘴红鲌的形态特征;梁宏伟等[17]利用主成分分析获得了长丰鲢和长江鲢15个指标85%以上的信息,较好地反映了长丰鲢和长江鲢的形态性状信息。本研究将13项性状比值简化为4个主成分综合指标,累积贡献率达84.003%,较好地反映了克氏原螯虾的形态信息。其中前两个主成分的累积贡献率为52.361%,反映了克氏原螯虾的腹节特征,其包括的忠县群体和高邮群体的信息比其它群体更为丰富,所以这10个群体的形态差异主要集中在腹节因子上。
判别分析是用来判别对象所属分类的一种统计分析方法,它主要通过观测已知样品的数据来构建一个判别函数,通过构建的函数来对未知样品进行分类[18]。陈杰等[19]对不同鳊鲂属鱼类进行判别分析,建立了6个判别函数,判别准确率在82%至100%之间。张帅[20]对138尾盖子红七彩神仙进行判别分析,建立了雌雄判别方程,雄鱼的准确率为71.4%,雌鱼的准确率为87.7%。本研究结果中,雌性群体中的南京群体判别率达到93.1%,表明对于南京群体的判别效果较好。雄性群体中,宜宾群体、南京群体和无锡群体的判别效果较好,判别准确率分别是100%,95.2%,85.7%,表明可以对这3个群体的雄性克氏原螯虾进行有效的判定。但雌雄的综合判别率较低,雌性为64.7%,雄性为65.6%。这可能是由于采样群体在形态上的差异并不显著,判别分析无法根据形态差异对群体做出正确率较高的分类。
现在的分类学家一般将亚种视为分类的最小单位,但有学者认为在亚种的水平下还可以进行不同种群的分类,并提出了差异系数的概念[21]。从Mayr等[22]将差异系数(CV)值1.28视为界定亚种水平的标准来看,本研究大部分形态比例参数的差异系数值小于1.28,说明这10个不同水域群体的克氏原螯虾群体的形态差异并未达到亚种水平。Avise等[23]认为亚种间的形态差异需同分子差异保持一致,虽然形态差异分析不能够准确地判定群体之间差异,但可为后续分子研究提供参考。
3.2 克氏原螯虾雌雄形态差异分析
T检验结果显示,雌雄克氏原螯虾的主要差异集中在体重、头胸甲长、头胸甲宽和腹节总长上,且雄性螯虾的平均体重比雌性要重,头胸甲平均长度比雌性长,头胸甲平均宽度也比雌性宽,但腹节总长平均值比雌性要小。而全长相同的情况下雄性体重一般大于雌性,可能与雄性克氏原螯虾的两只大螯通常比同体型的雌性的两只螯要粗壮以及雄性的头胸甲长长于雌性等有关。全长相同的情况下雄性的腹节总长短于雌性可能与克氏原螯虾雌虾抱卵、孵化幼苗的习性有关。
参考文献:
[1]邓沼泽.小龙虾的市场经济价值论述[J].现代商贸工业,2010,(13):155.
[2]王 蕊.克氏原螯虾的营养保健功能及相关食品的研究与开发[J].水产科技情报,2008,(1):24-27.
[3]龚世园,吕建林,孙瑞杰,等.克氏原螯虾繁殖生物学研究[J].淡水渔业,2008,38(6):23-25,30.
[4]陆剑锋,赖年悦,成永旭.淡水小龙虾资源的综合利用及其开发价值[J].农产品加工(学刊),2006,(10):47-52,63.
[5]李艳和.克氏原螯虾在我国的入侵遗传学研究[D].武汉:华中农业大学,2013.
[6]郑 友.不同地区野生克氏原螯虾形态差异及氨基酸的分析[D].南昌:南昌大学,2013.
[7]黄有辉.日本沼虾不同地理种群形态学及多样性研究[D].上海:华东师范大学,2016.
[8]王克行.虾蟹类增养殖学[M].北京:中国农业出版社,1997.
[9]李思发,李晨虹,李家乐.尼罗罗非鱼品系间形态差异分析[J].动物学报,1998,(4):75-82.
[10]韩晓磊,马 强,李小蕊,等.不同地区克氏原螯虾群体的形态差异分析[J].水生态学杂志,2011,(3):82-87.
[11]郑 友,胡火根,唐建清,等.不同水域克氏原螯虾群体的形态差异分析[J].南昌大学学报(理科版),2014,(1):96-102.
[12]张 萌,白 俊,金 辉,等.不同地理群体的克氏原螯虾形态差异多元分析[J].南昌大学学报(理科版),2016,(2):188-196.
[13]傅德印.主成分分析中的统计检验问题[J].统计教育,2007,(9):4-7.
[14]梁宏伟,曹力欢,李 翔,等.三个不同品系中华鳖形态差异分析[J].淡水渔业,2017,47(4):91-96.
[15]韩晓磊,马 强,李小蕊,等.中国螯虾科虾类(克氏原螯虾和东北螯虾)形态差异及判别分析[J].中国农学通报,2011,27(11):61-65.
[16]安 丽,孟庆磊,张龙岗,等.翘嘴红鲌雌雄个体的形态差异分析[J].淡水渔业,2017,47(2):36-41.
[17]梁宏伟,李 忠,罗相忠,等.长丰鲢与长江鲢形态差异与判别分析[J].水生生物学报,2015,39(5):1059-1064.
[18]王 静,夏结来,叶冬青.判别分析方法在医学应用中的进展[J].数理统计与管理,2008,(2):369-376.
[19]陈 杰,李福贵,黄创新,等.不同鳊鲂属鱼类群体的形态差异分析[J].上海:上海海洋大学学报,2014,(3):388-394.
[20]张 帅.七彩神仙形态指标体系分析及其雌雄鉴别模型[D].上海海洋大学,2013.
[21]曾 聪,阎里清,高泽霞,等.梁子湖、鄱阳湖和淤泥湖团头鲂的形态学比较[J].华中农业大学学报,2012,(1):88-94.
[22]Mayr E,Linsley E G,Usinger R L.Methods and principles of systematic zoology[M].New York and London:Mc Graw Hill,1953.
[23]Avise J C,Ball R M.Principles of genealogical concordance in species concepts and biological taxonomy [J].Oxford Surv Evol Biol,1990,(7):45-67.
