汉城湖浮游动物群落结构特征及与水质关系
2018-05-14王松陈红刘清许江娟张国王在照
王松,陈红,刘清,许江娟,张国,王在照,*
1.西北农林科技大学 动物科技学院,杨凌 712100 2.西安市水产工作站,西安 710054
1 前言
浮游动物不仅是滤食性鱼类的饵料,还能够制约微生物的发生,并且对水体浮游植物群落结构变化有重要影响,对水体的物质循环、能量流动和生物资源的补充,起着非常关键的作用,是水生态系统中重要的组成部分[1–3]。浮游动物个体小,生长周期短,对水环境的变化较为敏感,其种群结构、数量变化、优势种及污染指示种、物种多样性等都能作为水体营养化的重要标志,间接地反应水体质量,因此浮游动物在水质监测调控、水污染修复治理方面的作用逐渐受到重视[4–5]。
汉城湖,原名为团结水库,坐落于西安市西北方向,全长6.27公里,水深5米左右,湖面宽20至100米,水面面积56.67 hm2,总库容约137万m3,系改革开放初期为解决西安市排水排污问题而建立,然而随着湖体水质不断恶化,2006年西安市政府决定启动环境综合治理工程,将40公里外的沣三干渠沣水注入汉城湖,年引水量达3500万m3,到2009年治理已取得显著成效,改造后的汉城湖融合了独特的汉文化元素,彰显了古都西安丰厚的历史风貌。当前的汉城湖已经成为国家水利风景区、省级水利部摄影基地、国家级水土保持科技示范园和国家4A级旅游景区,形成了集生态景区、旅游文化、防洪灌溉、文物保护等为一体的生态公园[6],经过为期十年的治理,汉城湖的水质治理效果及水生态环境备受瞩目。目前,关于汉城湖浮游动物群落结构的调查还未见报道,本研究于2015年3月至12月期间,对汉城湖浮游动物群落结构进行了调查,从浮游动物种类组成、丰度、生物量、优势种、污染指示种和多样性指数等多个角度评价了汉城湖水体营养化状况,为汉城湖水体的营养化评价及可持续发展提供了基础资料和理论依据。
2 材料与方法
2.1 采样点的设置
根据汉城湖自然环境状况与人类活动区域分布等特征,设置了5个采样点(图1),1号至5号采样地点分别位于: 封禅广场(N34°19′32″,E108°55′30″)、渔港(N34°18′36″,E108°55′29″)、大风阁(N34°18′18″,E108°55′25″)、湖心岛(N34°18′14″,E108°54′11″)和安门码头(N34°17′46″,E108°53′53″), 封禅广场为湖的上游入水口,安门码头为湖的下游出水口,采样时间为2015年3月20号、6月20号、9月20号和12月20号,均在当天上午进行并完成采样工作。
2.2 样品采集与处理
浮游动物定性采集方法:用25号浮游生物网,在水面下0.5 m至水面以“∞”状缓慢拖动5 min,将水样浓缩于网头部位,收集到50 mL标本瓶中,现场加入福尔马林溶液固定,使最终浓度为4%,带回实验室进行镜检,定性分析参照《中国淡水生物图谱》[7]和《淡水微型生物与底栖动物图谱》[8]进行。轮虫和原生动物的定量采集方法:用规格为5 L的采水器,从水面以下0.5 m处采集水样5 L,摇匀后取水1 L,加入福尔马林溶液固定保存,带回实验室静置沉淀48 h后,用虹吸方法浓缩至10 mL;枝角类和桡足类定量采集:用采水器取表层0.5 m处的水20 L,用25号网过滤浓缩,加福尔马林溶液固定。移取1 mL浓缩样至1 mL规格计数框内进行样品的定量,在10×10倍显微镜下全片计数,同一样品至少计数3次,偏差小于15%即为有效值。原生动物和轮虫的生物量(湿重)测定参照《淡水浮游生物研究方法》[9]的体积法进行,通过测量生物的体积并假定比重为1,则可计算生物体重;桡足类和枝角类的生物量(湿重)是根据体长-体重回归方程[10–11],通过测量体长(mm)计算出体重(mg)。

图1 汉城湖采样位点分布图Fig.1 Location of sampling sites in Hancheng Lake
水体理化指标的测定:水体透明度(SD)用萨氏盘测定;用5 L有机玻璃采水器取表层0.5 m处的水,用温度计测量水温(WT);水样经0.45 μm醋酸纤维滤膜过滤,丙酮萃取,分光光度法测叶绿素a(chl a)含量;溶解氧(DO)的测定采用碘量法(GB 7489—87);总磷(TP)用钼酸铵分光光度法(GB 11893—89);总氮(TN)用碱性过硫酸钾紫外分光光度法(GB 11894—89);化学需氧量(COD)用碱性高锰酸钾法(GB 11892—89)[12]。
2.3 数据处理与分析
Jaccard相似性系数:K=c/(a+b-c),其中a、b分别表示采样点A和B各自的物种数,c表示采样点A和B共有的物种数,K值在0—0.25为极不相似,0.25—0.50为中等不相似,0.50—0.75为中等相似,0.75—1.00 为极相似[13]。
用Y=(Ni/N)·fi来计算浮游动物的优势度,式中Ni为第i种的个体数,N为所有种类总个体数,fi为第i种出现的频率,Y≥0.02的物种为优势种。
优势种更替率:R=(a+b–2c)/(a+b–c)×100%, 其中a、b表示相邻两个季度的优势种数,c表示相邻两个季度共同优势种数[14]。
运用叶绿素a(chl a)、总磷(TP)和透明度(SD)计算综合营养状态指数(TSI),计算公式为:
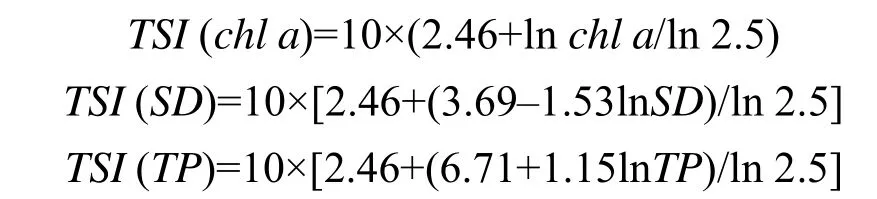
TSI=0.540×TSI(chl)+0.297×TSI(SD)+0.163×TSI(TP)
水体营养化水平评价标准为:TSI<37为贫营养型;37≤TSI<53为中营养型;TSI≥53为富营养型[15]。
E/O值为轮虫富营养种数与贫营养种数的比值,可以评价水体营养化程度:E/O<0.5为贫营养型,0.5—1.5为中营养型,1.5—5.0为富营养型,>5为超富营养型[16]。
QB/T值表示臂尾轮虫属(B)的种数与异尾轮虫属(T)的种数的比值,可以用作判断水质的类型:QB/T<1为贫营养型,1—2为中营养型,>2为富营养型[17]。
肥度指数(E)可作为评价水体富营养化的指标,其计算公式为:E=K·(x+1)/(A+Y)/(y+1),式中:K表示轮虫种数;A表示桡足类种数;Y表示枝角类种数;x表示富-中营养型指示种;y表示贫-中营养型指示种。E<0.2为贫营养型,0.2—1.0为中营养型,1.0—4.0为富营养型,>4.0为超富营养型[18]。
污生指数(S)可对水质进行综合评价,计算公式为S=∑m·h/∑h,式中:m为浮游动物种的污染带指数值(寡污型为1,β-中污型为2,α-中污型为3,多污型为4);h为种的相对数量表示值(偶见种为1,常见种为3,多见中为5)。污生指数(S)对应的水质评价标准为:1.0—1.5为寡污型,1.5—2.5为β-中污型,2.5—3.5 为α-中污型, >3.5 为多污型[19]。
浮游动物的多样性评价采用Shannon-Wiener多样性指数(H)、Margalef丰富度指数(d)和pielou均匀度指数(J)[20–22]。其计算公式如下:
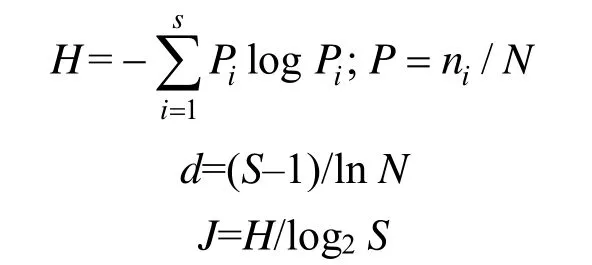
式中,ni表示第i种的个体数;N表示所有种类总个体数;S表示总种类数。多样性指数对应的水质评价标准为:(H):0—1为多污型,1—2为α-中污型,2—3为β-中污型,>3为清洁-寡污型;(d):0—1为多污型,1—2为α-中污型,2—3为β-中污型,3—4为寡污型,>4为清洁型;(J): 0—0.3为多污型,0.3—0.5为中污型,0.5—0.8为寡污型,0.8—1.0为清洁型。
采用Cancoo 4.5软件将浮游动物数据做去趋势对应分析(detrended correspondence analysis,DCA),4个轴中梯度最大值大于4,因此采用单峰模型的典范对应分析(canonical correspondence analysis,CCA)来解释浮游动物种类与环境因子的关系。为减少机会种群对结果的干扰,选择优势度Y≥0.02的物种进行排序,为使数据更趋于正态分布,物种数据与环境数据均进行lg(x+1)转换,排序结果用物种-环境因子关系的双序图表示。
3 结果与分析
3.1 浮游动物种类组成
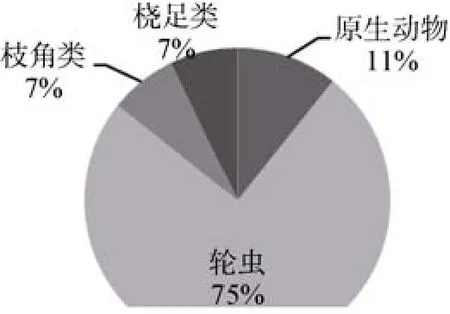
图2 汉城湖浮游动物的种类组成Fig.2 Species composition of zooplankton in the Hancheng Lake
调查期间,共检测出浮游动物28种,隶属于4门21属,其中原生动物3属3种,占总种类数的11.1%;轮虫14属21种,占77.8%;枝角类2属2种,占3.7%;桡足类2属2种,占7.4%(图2)。轮虫是汉城湖浮游动物的主要组成部分,原生动物、枝角类和桡足类的种类数均较少。从季节分布来看,春季、冬季出现的浮游动物种类数最多,分别达20种和19种,夏季、秋季浮游动物种类较少,分别为11种和13种,王氏似铃壳虫Tintinnopsis wangi、前节晶囊轮虫Asplanchna priodonta、长三肢轮虫Filinia longiseta、纤巧异尾轮虫Trichocerca tenuior、萼花臂尾轮虫Brachionus calycifloru和真翅多肢轮虫Polyarthra euryptera全年均能被检测到(表1)。
3.2 浮游动物群落结构
运用Jaccard相似性系数(K)对汉城湖浮游动物的空间分布特征进行分析,结果如表2所示,各采样点之间的浮游动物相似性系数(K)值为0.54—0.75,平均值为0.63,其中3号与5号采样点之间的K值最小,1号与2号采样点之间的K值最大,10组采样点的相似性均达到了中等相似水平,表明汉城湖各水域之间种群相似度较高,水体生态环境差异不大。
从图3和图4可知,浮游动物丰度的变化范围为17—48个·L-1,平均值为29个·L-1;
生物量的变化范围为0.012—0.058 mg·L-1,平均值为0.036 mg·L-1。轮虫平均丰度和生物量最高,分别为23个·L-1和0.023 mg·L-1,分别占平均丰度和生物量总量的80.8%和63.1%;原生动物平均丰度次之,占总平均丰度的17.2%,然而平均生物量偏低,仅为0.002 mg·L-1,占总平均生物量的6.0%;枝角类和桡足类的平均丰度较低,均仅占总平均丰度的1.0%,但桡足类的生物量较高,仅次于轮虫生物量,为0.009 mg·L-1,占总平均生物量的24.9%,枝角类生物量偏低,仅为0.002 mg·L-1。
各季节浮游动物丰度和生物量大小也有差异,平均丰度大小排序为:夏季(48个·L-1)>秋季(30个·L-1)>冬季(20个·L-1)>春季(17个·L-1);平均生物量大小排序为: 夏季(0.058 mg·L-1)>秋季(0.038 mg·L-1)>春季(0.037 mg·L-1)>冬季(0.012 mg·L-1)。就不同种类的浮游动物而言,丰度和生物量在不同季节也表现出一定差异:原生动物在春、冬季节丰度较高,所占比例分别为34.2%、53.2%,在冬季生物量比例较高,占20.3%,在其余季节生物量比例均偏低;轮虫丰度在夏、秋季节占绝对优势,占这两个季节总
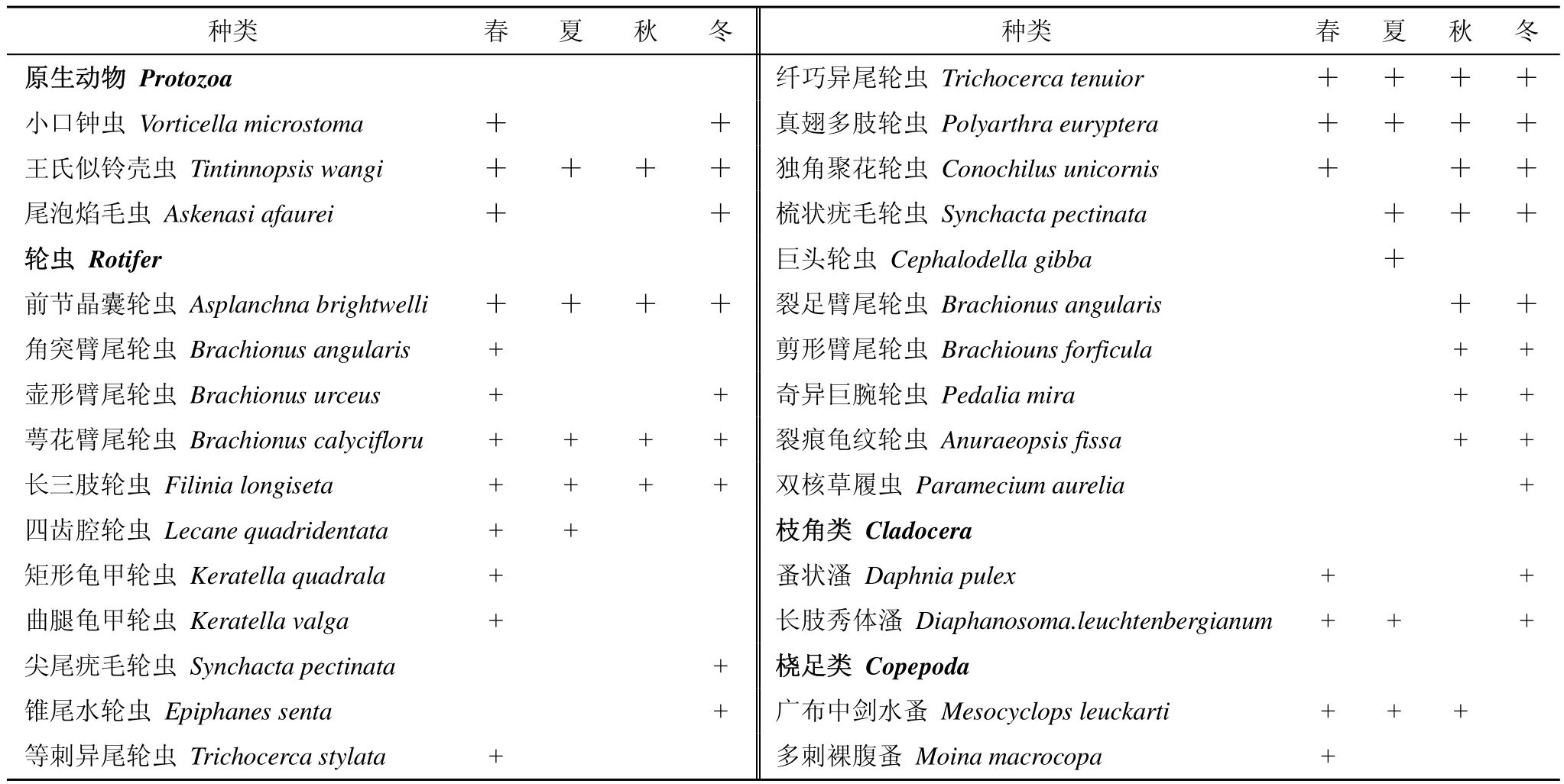
表1 浮游动物种类组成及分布Tab.1 Zooplankton composition and distribution in each investigation

表2 各采样点之间的浮游动物相似性系数Tab.2 Similarity coefficients of zooplankton communities among 5 sampling sites in Hancheng Lake
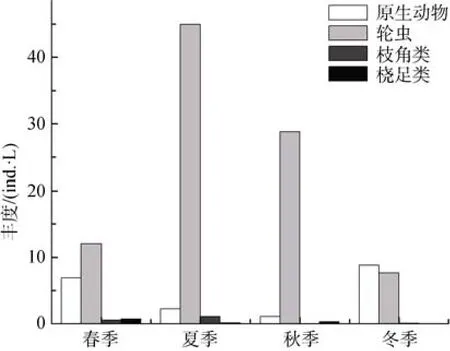
图3 汉城湖浮游动物丰度的季节变化Fig.3 Seasonal variation of zooplankton abundance in Hancheng Lake

图4 汉城湖浮游动物生物量的季节变化Fig.4 Seasonal variation of zooplankton biomass in Hancheng Lake
3.3 浮游动物优势种
丰度的 92.8%、95.6%,轮虫生物量在春季次于桡足类,占春季总生物量的29.1%,但在其他季节轮虫生物量比例均最大;枝角类和桡足类的丰度在各季节均较低,尽管如此,枝角类的生物量在夏、冬季节仅次于轮虫,分别占夏、冬季节总生物量的15.1%和38.5%,桡足类在春季的生物量显著高于其他浮游动物,占春季总生物量的56.5%,在冬季仅次于轮虫,占冬季生物量总量的15.1%。
在四次采样中,共检出了浮游动物优势种20种,轮虫优势种数最多,有16种,其次是原生动物,有3种,桡足类1种,枝角类未发现有优势种。从表3可以看出,在不同季节,优势种在种类和数量上存在一定差异,春季优势种有8种;夏季4种;秋季9种;冬季7种。真翅多肢轮虫Polyarthra euryptera全年均为优势种;王氏似铃壳虫Tintinnopsis wangi除了春季,其它季节均属于优势种;萼花臂尾轮虫Brachionus calycifloru、真翅多肢轮虫Polyarthra euryptera和前节晶囊轮虫Asplanchna brightwelli在夏季、秋季均形成优势种;小口钟虫Vorticella microstoma在春季、冬季成为优势种。通过计算浮游动物优势种更替率R值,发现汉城湖浮游动物优势种的季节更替存在一定差异,春-夏季的R值为90.9%;夏-秋季R值为55.6%;秋-冬季R值为85.7%,除夏季-秋季外,其余季节的浮游动物优势种均有明显的更替。
3.4 浮游动物与环境因子的相关性分析
汉城湖各季度理化因子平均值见表4, 通过CCA分析显示,轴1与水温(WT)(P=–0.652)、叶绿素 a(chl a)(P= –0.872)和 COD(P= –0.900)有极显著的负相关性,与透明度(SD)(P=0.667)有显著的正相关性;轴2与水温(WT)(P=–0.714)显著负相关,与总磷(TP)(P=0.853)、总氮(TN)(P=0.962)和溶解氧(DO)(P=0.944)有极显著的正相关性。尾泡焰毛虫Askenasi afaurei、尖尾疣毛轮虫Synchacta pectinata、双核草履虫Paramecium aurelia和锥尾水轮虫Epiphanes senta与总磷(TP)和溶解氧(DO)呈显著的正相关性;王氏似铃壳虫Tintinnopsis wangi与总磷(TP)显著正相关;真翅多肢轮虫Polyarthra euryptera与化学需氧量(COD)表现出较高的正相关性;萼花臂尾轮虫Brachionus calycifloru和前节晶囊轮虫Asplanchnabrightwelli与叶绿素a(chl a)呈显著正相关性;裂足臂尾轮虫Brachionus angularis、剪形臂尾轮虫Brachiouns forficula、纤巧异尾轮虫Trichocerca tenuior和梳状疣毛轮虫Synchacta pectinata与叶绿素a(chl a)和水温(WT)均呈显著正相关性;小口钟虫Vorticella microstoma、长三肢轮虫Filinia longiseta、角突臂尾轮虫Brachionusangularis、壶形臂尾轮虫Brachionus urceus、矩形龟甲轮虫Keratella quadrala、曲腿龟甲轮虫Keratella valga和广布中剑水蚤Mesocyclops leuckarti与各环境因子的相关性不显著。
3.5 水体质量评价
通过计算综合营养状态指数(TSI)可以得出,春季至冬季,汉城湖水体的TSI值分别为49.9、59.3、62.7和53.5,对照水体营养化评价标准,除春季为中营养水平以外,其余季节均为富营养化水平。
浮游动物的丰度和生物量可作为判断水体营养化程度两种指标,浮游动物丰度<1000 ind·L-1为贫营养型,1000—3000 ind·L-1为中营养型, >3000 ind·L-1为富营养型[19];浮游动物生物量<1为贫营养型,1—3为中营养型,>3为富营养型[18],汉城湖全年浮游动物丰度和生物量均较低,因此以丰度和生物量作为水体营养化水平指标时,汉城湖水体为贫营养型。
汉城湖浮游动物贫-中营养型种类有:裂痕龟纹轮虫Anuraeopsis fissa、矩形龟甲轮虫Keratella quadrala、纤巧异尾轮虫Trichocerca tenuior和独角聚花轮虫Conochilus unicornis,富-中营养型种类有:角突臂尾轮虫Brachionus angularis、壶形臂尾轮虫Brachionus urceus、萼花臂尾轮虫Brachionus calycifloru、裂足臂尾轮虫Brachionus angularis、剪形臂尾轮虫Brachiouns forficula、锥尾水轮虫Epiphanes senta、巨头轮虫Cephalodella gibba、奇异巨腕轮虫Pedalia mira、长三肢轮虫Filinia longiseta、多刺裸腹溞Moina macrocopa、真翅多肢轮虫Polyarthra euryptera和小口钟虫Vorticella microstoma。春季至冬季,汉城湖浮游动物E/O指数分别为2.3、4、3和2.7,均值为3;QB/T指数分别是3、1、3和4,均值为2.75;肥度指数(E)指数分别为11、10、25和29,均值为19。E/O指数、QB/T指数和肥度指数(E)指数均值对应的水体营养化类型分别为富营养型、富营养型和超富营养型。
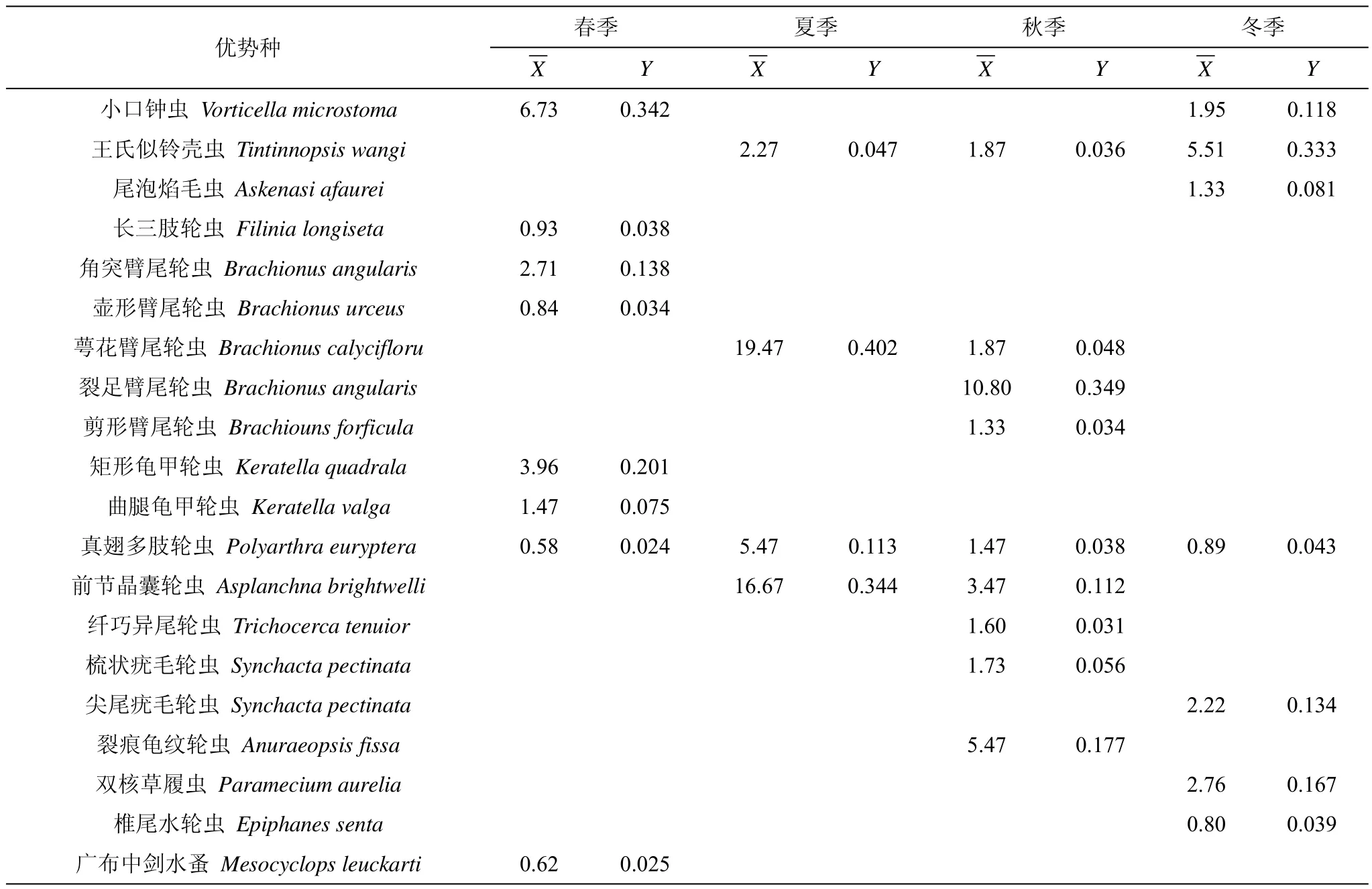
表3 浮游动物优势种及平均丰度Tab.3 Dominant species and mean abundance of zooplankton
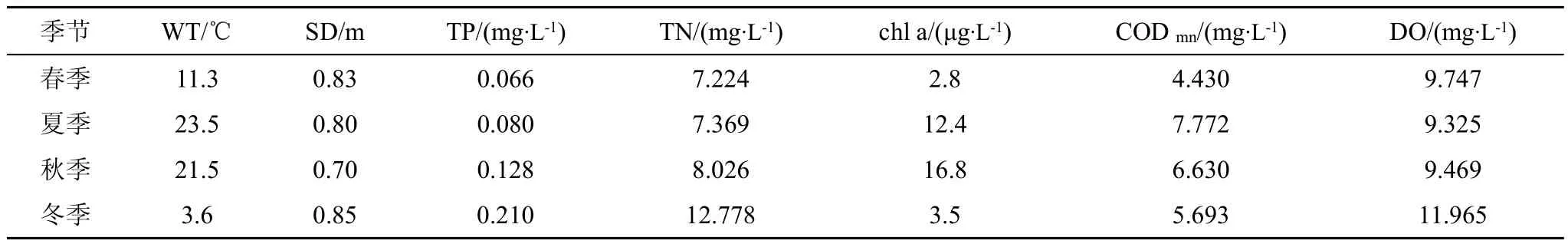
表4 汉城湖水质理化指标检测情况Tab.4 The survey result of chemical and physical index of Hancheng Lake in 4 seasons

图5 四个季节不同采样点的浮游动物物种多样性Fig.5 Diversity of zooplankton community at each sampling site in 4 seasons
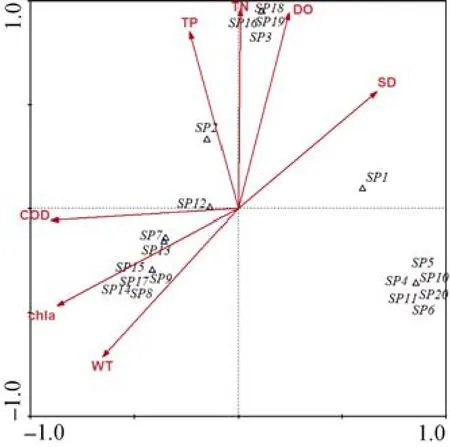
图6 浮游动物与环境因子关系的CCA排序图Fig.6 CCA diagram of zooplankton dominant species and environmental factors
根据不同浮游动物所指示的水质类型,将污染指示种划分为4个等级:寡污型;β-中污型;α-中污型和多污型,通过参考相关资料[23–24]共鉴定出浮游动物污染指示种17种,其中寡污型指示种3种,β-中污型指示种6种,α-中污型指示种8种,汉城湖春季至冬季污生指数值(S)分别为:2.6、2.3、2.9和2.3,平均值为2.52,污生指数值(S)均值显示汉城湖水质整体为α-中污染型。
浮游动物多样性指数不仅能反应群落结构稳定性、评价物种资源丰富度、衡量种群演替进程,也可用作评价水质优劣的标准[25],因此运用浮游动物多样性指数来反映汉城湖水体营养化程度。从图(3)可以看出,汉城湖多样性指数(H)最大值为2.70,最小值为1.95,夏季为α-中污型,其余季节均为β-中污型;丰富度指数(d)最大值为2.36,最小值为1.15,除夏季为α-中污型,其余季节均为β-中污型;均匀度指数(J)最大值为0.90,最小值为0.75,除了冬季属于清洁型,其余季节均为寡污型。汉城湖全年浮游动物多样性指数H、d和J的均值分别为2.46、1.81、0.79,其对应的水质类型分别为β-中污型、α-中污型和寡污型,多样性指数的综合结果显示,汉城湖水质整体为中污染型。
4 讨论
4.1 浮游动物群落结构分析
汉城湖浮游动物群落结构特征为个体较小的轮虫、原生动物在种类和丰度上占优势,而个体较大的枝角类、桡足类在种类和丰度上均较少,浮游动物结构明显趋于小型化,这种现象在很多不同的水体中均有发现[26–28]。枝角类与轮虫之间存在竞争关系,当枝角类的种类和丰度偏低时,轮虫的抑制作用被减弱,导致轮虫大量繁殖[29–30]。汉城湖枝角类的种类、数量较低,因此对轮虫可能会形成较小的竞争压力。也有很多研究认为,鱼类捕食压力是造成这一现象的重要原因[3,31–33]。为净化水质,营造良好的水体景观环境,近年来,汉城湖每年会进行增殖放流活动,大量的鲢、鳙、锦鲤和鲫鱼被投放到汉城湖中,研究表明,鱼类在同等能耗下进行捕食时,会优先选择个体较大的浮游动物[34–35],鲢、鳙作为虑食性鱼类,其投放密度的增加能有效地降低枝角类、桡足类等个体较大的浮游动物生物量,而对轮虫等小型浮游动物的生物量不但不会降低,反而会增加,使浮游动物种群趋于小型化[36–37],因此推断,汉城湖水体浮游动物小型化现象,可能是由于以大型浮游动物为食物的鲢、鳙鱼大量投放造成的。另外,夏、秋季节是藻类大量繁殖生长的季节,有学者指出,藻类的大量存在会导致一些大型枝角类尤其溞属因滤食器官堵塞而难以生存[38],汉城湖春、冬季节均发现有大型枝角类—蚤状溞Daphnia pulex存在,而夏、秋季节均未检测到,推测可能与汉城湖浮游植物的爆发有关。
4.2 浮游动物与环境因子关系
通过浮游动物丰度与环境因子相关性分析发现,汉城湖浮游动物主要与水温、总氮、溶解氧、透明度以及叶绿素a有关,少数浮游动物与化学需氧量、总磷有一定关系。水温作为影响浮游动物生长、发育、繁殖和群落演替极为重要的非生物因子,能间接地影响浮游动物种群结构[39–40],研究显示,轮虫的数量高峰一般出现在20℃以上的水温[27],汉城湖在夏、秋季节水温均高于20℃,轮虫的丰度和生物量较大,说明水温可能是影响汉城湖浮游动物种群结构的主要因素之一。透明度能在一定程度上反映浮游植物的丰度,浮游植物越多,透明度越低,反之越高,汉城湖水体透明度与多种浮游动物显著负相关,也与叶绿素a也极显著负相关,邱小琮[41]调查宁夏沙湖发现,水体的透明度对浮游动物的影响主要是通过叶绿素a含量间接影响,并且呈负相关性。汉城湖水体溶解氧与多种浮游动物表现显著的正相关性,表明溶解氧是影响浮游动物分布的重要环境因素之一,这与杜明敏[42]、吴利[43]等人的调查结果相一致。汉城湖水体化学需氧量、总磷和总氮与多种浮游动物呈正相关关系,有研究表明,小型浮游动物多以浮游藻类为主,水中营养盐(N、P)和有机物(COD)对浮游动物群落结构的影响,主要是通过直接作用于浮游植物间接对浮游动物产生影响[40,44]。叶绿素a作为水体初级生产力的标志,反映了水体浮游植物的状况,通过调查武汉东湖和广东省的城市湖泊时发现,浮游植物丰度与叶绿素a含量极为相关[45–46],当叶绿素a水平在 5—19 μg/L水平时,蓝藻与叶绿素a显著正相关[47]。汉城湖夏秋季节叶绿素a水平较高,多种轮虫表现出与叶绿素a具有明显的正相关性,因此推测,轮虫可能是通过利用蓝藻进行大量繁殖,进而形成优势种群。汉城湖的广布中剑水蚤Mesocyclops leuckarti与其他环境因子相关性均不明显,可能与桡足类受环境因子的影响较小有关[32]。另外,由于水体环境与浮游动物的关系错综复杂,多种浮游动物如小口钟虫Vorticella microstoma等与调查的环境因子没有表现出显著的相关性,这有待于我们进一步深入考察和研究,以得出更好的解释。
4.3 浮游动物种群结构与水质评价
以往水质检测常用到的方法是理化分析法,本研究也采用了综合营养状态指数(TSI)方法对汉城湖的水质进行了评估,结果显示汉城湖整体处于中-富营养水平,但其结果仅代表采样时河流的瞬时状况,不能反映河流过去一段时间所受的污染。而浮游动物游泳能力弱或完全没有游泳能力,常悬浮于水体表面以水中的有机碎屑等为食,对于生活在一定地段的浮游动物,集合了自身整个生活周期的环境因素,更能反应一段时间的水质情况,因此我们采用了生物学的方法,将浮游动物丰度、生物量、E/O指数、QB/T指数、E指数、污生指数(S)和多样性指数等多种能反映水体营养化水平的指标结合起来,比较全面地分析了汉城湖在2015年的水体营养化状况,各项分析结果显示,汉城湖水体整体处于中营养化至富营养化水平之间,与理化因子检测显示的结果是一致的。
参考文献
[1]刘建康,黄祥飞,林婉莲,等.东湖生态学研究[M].北京:科学出版社,1995.
[2]李超伦,王克.植食性浮游桡足类摄食生态学研究进展[J].生态学报,2002,22(4):593–596.
[3]高子涵,张健,皮杰,等.湖南省大通湖浮游动物群落结构及其与环境因子关系[J].生态学杂志,2016,35(3):733–740.
[4]吴利,冯伟松,张堂林,等.湖北省西凉湖浮游动物群落周年动态变化及其与环境因子的关系[J].湖泊科学,2011,23(4):619–625.
[5]刘歆璞,王丽卿,张宁,等.青草沙水库后生浮游动物群落结构及其与环境因子的关系[J].生态学杂志,2013,32(5):1238–1248.
[6]蒋建军.试论西安市汉城湖对汉文化与水文化的传承与创新[J].水利发展研究,2015,15(11):81–84.
[7]韩茂森,束蕴芳.中国淡水生物图谱[M].北京:海洋出版社,1995.
[8]周凤霞.淡水微型生物与底栖动物图谱[M].北京:化学工业出版社,2011.
[9]章宗涉,黄详飞.淡水浮游生物研究方法[M].北京:科学出版社,1991.
[10]陈雪梅.淡水桡足类生物量的测算[J].水生生物学报,1981(3):397–408.
[11]赵文.水生生物学[M].北京:中国农业出版社,2005.
[12]国家环境保护总局.水和废水监测分析方法(第四版)[M].北京:中国环境科学出版社,2002:88–284.
[13]高原,赖子尼,李捷,等.连江浮游动物多样性的空间分布[J].生物多样性,2013,21(6):699–708.
[14]李强,田华,姜民,等.淀山湖浮游动物群落结构特征及其影响因子[J].水生态学杂志,2015,36(4):69–77.
[15]袁聪,陶诗雨,张莹莹,等.安康水库表层浮游藻类群落结构及其与环境因子的关系[J].应用生态学报,2015,26(7):2167–2176.
[16]陈立婧,顾静,彭自然,等.上海崇明岛明珠湖轮虫群落结构[J].应用生态学报,2009,20(12):3057–3062.
[17]张觉民,何志辉.内陆水域渔业自然资源调查手册[M].北京:农业出版社,1991.
[18]鞠永富,于洪贤,于婷,等.西泉眼水库夏季浮游动物群落结构特征及水质评价[J].生态学报,2016,36(16):5126–5132.
[19]王凤娟,胡子全,汤洁,等.用浮游动物评价巢湖东湖区的水质和营养类型[J].生态科学,2006,25(6):550–553.
[20]WHITTAKER,R.H.Evolution and measurement of species diversity[J].Taxon,1972,21(2/3):213–251.
[21]MARGALEF D R.Information theory in ecology[J].General-Systems,1958,3:36–71.
[22]PIELOU E C.Species-diversity andpattern-diversity in the study of ecological succession[J].Journal of Theoretical Biology,1966,10(2):370–383.
[23]王凤娟.巢湖东半湖浮游生物与水质状况及营养类型评价[D].合肥:安徽农业大学,2007.
[24]陈立婧,顾静,彭自然,等.滆湖轮虫群落结构与水质生态学评价[J].动物学杂志,2008,43(3):7–16.
[25]ACHENBACH L,LAMPERT W.Effects of elevated temperatures on threshold food concentrations and possible competitive abilities of differently sized cladoceran species[J].Oikos,1997,79(3):469–476.
[26]姜胜,黄长江,陈善文,等.2000—2001年柘林湾浮游动物的群落结构及时空分布[J].生态学报,2002,22(6):828–840.
[27]陈光荣,钟萍,张修峰,等.惠州西湖浮游动物及其与水质的关系[J].湖泊科学,2008,20(3):351–356.
[28]黄简易,朱艺峰,王银,等.象山港浮游动物群落结构时空变化的定量驱动分析[J].生态科学,2014,33(4):713–722.
[29]NOGUEIRA M G.Phytoplankton composition,dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir(Paranapanema River),São Paulo,Brazil[J].Hydrobiologia,2000,455(2/3):115–128.
[30]杨亮杰,吕光汉,竺俊全,等.横山水库浮游动物群落结构特征及水质评价[J].水生生物学报,2014,38(4):720–728.
[31]杨宇峰,黄祥飞.鲢鳙对浮游动物群落结构的影响[J].湖泊科学,1991,4(3):78–86.
[32]张世羊,周巧红,成水平,等.复合养殖系统中浮游动物种类丰度及其影响因素[J].中国环境科学,2009,29(7):745–750.
[33]陈炳辉,刘正文.滤食杂食性鱼类放养对浮游动物群落结构的影响[J].生态科学,2012,31(2):161–166.
[34]HALL D J,THRELKELD S T,AND CW B,et al.The Size-Efficiency Hypothesis and the Size Structure of Zooplankton Communities[J].Annual Review of Ecology and Systematics,1976,7(1):177–208.
[35]YANG Y F,HUANG X F,LIU J K,et al.Effects of fish stocking on the zooplankton community structure in a shallow lake in China[J].Fisheries Management and Ecology,2005,12(12):81–89.
[36]KAJAK Z.Thepossibilities of fish,especially silver carp to overcome water bloom in temperature water bodies[J].Symp Biol Hung,1979,19:77–86.
[37]赵玉宝.鲤鱼种和鲢鳙对池塘浮游生物的影响[J].生态学报,1993,03(4):348–355.
[38]郭飞飞,张云,赵广,等.金沙河水库浮游动物群落结构及其与环境因子的关系[J].生态学杂志,2016,35(8):2208–2216.
[39]SARMA1 S S S,NANDINI S,GULATI R D.Life history strategies of cladocerans:comparisons of tropical and temperate taxa[M].Aquatic Biodiversity II.Springer Netherlands,2005:315–333.
[40]PINTOCOELHO R,PINELALLOUL B,Méthot G,et al.Crustacean zooplankton in lakes and reservoirs of temperate and tropical regions:variation with trophic status[J].Canadian Journal of Fisheries and Aquatic Sciences,2005,62(2):348–361.
[41]邱小琮,赵红雪,孙晓雪.沙湖浮游动物与水环境因子关系的多元分析[J].生态学杂志,2012,31(4):896–901.
[42]杜明敏,刘镇盛,王春生,等.中国近海浮游动物群落结构及季节变化[J].生态学报,2013,33(17):5407–5418.
[43]吴利,李源玲,陈延松.淮河干流浮游动物群落结构特征[J].湖泊科学,2015,27(5):932–940
[44]DRUVIETISI,KLAVINSM,URTANEL,etal.Evaluation of plankton communities in small highly humic bog lakes in Latvia[J].Environment International,1998,24(5):595–602.
[45]蔡庆华.武汉东湖浮游生物间相互关系的多元分析[J].中国科学院大学学报,1995,12(1):97–102.
[46]陈光荣,雷泽湘,谭镇,等.环境因子对广东城市湖泊后生浮游动物的影响[J].水生态学杂志,2010,03(4):31–35.
[47]郭劲松,陈园,李哲,等.三峡小江回水区叶绿素a季节变化及其同主要藻类的相互关系[J].环境科学,2011,32(4):976–981.
