深圳大鹏半岛自然保护区钝叶假蚊母树群落特征
2018-05-14刘海军郭强张信坚孙红斌廖文波
刘海军,郭强,张信坚,孙红斌,廖文波,*
1.深圳市野生动物救护中心,深圳 518035 2.深圳市野生动植物保护管理处,深圳 518035 3.中山大学生命科学学院,有害生物控制与资源利用国家重点实验和广东省热带亚热带植物资源重点实验室,广州 510275 4.深圳市自然保护区管理中心,深圳 518035
1 前言
钝叶假蚊母树[Distyliopsis tutcheri(Hemsl.)P.K.Endr.],隶属金缕梅科(Hamamelidaceae)假蚊母树属(DistyliopsisP.K.Endr.)[1],为常绿灌木或小乔木,多分布于福建、广东、海南等地的山地常绿林中[2–3],该种也植物区系的孑遗种,常绿阔叶林群落的特征种。目前,国内关于钝叶假蚊母树的报道极少,仅见有关于其化学成分研究的报道[2]。而关于钝叶假蚊母树种群及其所在群落的特征的研究至今未见报道。根据2015年度的调查,在深圳大鹏半岛的主峰排牙山发现有较大面积的钝叶假蚊母树群落。该群落分布在山顶,受人为因素干扰少,保存较为完整。针对钝叶假蚊母树群落进行样方调查,进而对该群落的外貌、组成结构、物种多样性、地理成分性质等进行分析,旨在充分掌握钝叶假蚊母树群落的动态和演替特征,为加强保护和管理提供依据。
2 研究区、样地概况与研究方法
研究区概况
大鹏半岛自然保护区位于广东省深圳市东部,南临大亚湾和西涌湾,北接惠州地区,地理位置为114°17¢—114°22¢E,22°37¢—22°39¢N, 海拔 0—707 m。保护区地处南亚热带海洋性季风气候区,四季温和,雨量充足,日照时间长,夏季高温多雨,冬季干旱稍冷,年平均气温22.4℃,最高气温达36.6℃,最低气温为1.4℃,年平均降雨量为1948.4 mm。由于大鹏半岛北、东、南三面濒海,常年海风较大,并以东南风为主。地形以中低山地、丘陵为主,沟壑纵横,分布多个中小型人工水库。排牙山出露的岩性主要为沉积岩类,土壤多为变质岩类风化发育而成的壤土或重壤土,代表性土壤类型为赤红壤,土层厚度在1 m以内。钝叶假蚊母树群落位于排牙山主峰山顶附近, 海拔 658 m, 地理坐标 114°31¢58.062¢¢E,22°37¢7056¢N。
2.1 样地设置与调查
选择钝叶假蚊母树占优势的特征性群落,设置面积为1400 m2标准样地,依据群落调查方法[4]将样地划分为14个10 m×10 m样方,应用每木记账调查法进行调查,起测径阶≥1.5 cm,高度≥1.5 m,记录树种名称、胸径、高度、冠幅等;并在每个10 m×10 m样方内设置1个2 m×2 m的小样方,调查草本植物和乔、灌木幼苗,记录种名、株数、高度和覆盖度等。
2.2 数据分析
根据样方调查获得的相关数据,对钝叶假蚊母树群落的种类组成、群落结构、物种多样性及种群分布格局等进行分析。
(1)优势种群的重要值:根据《植物群落学实验手册》[4],计算群落中各种群的相对多度(RA)、相对频度(RF)、相对显著度(RD)、相对盖度(RC)和重要值(IV)等,其中,乔木层种群重要值IV=RD+RA+RF,灌木层种群重要值IV=RC+RA+RF。
(2)优势种群的年龄结构:以胸径级代替年龄[5],按照株高(H)及胸径(DBH)划分,采用5级立木标准划分,即:Ⅰ级为苗木,H<33 cm;Ⅱ级为小树,H≥33 cm,DBH<2.5 cm;Ⅲ级为壮树,2.5≤DBH<7.5 cm;Ⅳ级为大树,7.5≤DBH<22.5 cm;Ⅴ级为老树,DBH≥22.5 cm[4]。针对灌木或小乔木种群,再用立木径级代替5级立木标准划分种群年龄结构。共划分8个径级,每2 cm为一个径级,第1径级为0—2 cm,第2径级为2—4 cm,以此类推,最后把>14 cm 的归为第 8 径级[6–7]。
(3)各种群的频度分析:采用Raunkiaer频度级[8],将频度划分为5个等级,即:1%—20%为A级,21%—40%为B级,41%—60%为C级,61%—80%为D级,81%—100%为E级。
(4)群落物种多样性:根据Simpson多样性指数(D)、Shannon-Wiener多样性指数(H),以及Pielou均匀度指数(Jsw)进行测度[9]。计算公式如下:

其中,Pi=Ni/N,N为乔木层、灌木层或草本层各层的个体总数;Ni为第i个植物种的个体数;S则为乔木层、灌木层或草本层各层的物种总数。
3 研究结果
3.1 群落种类组成分析
根据样方调查统计,该群落共有维管植物106种,隶属于47科75属。包括蕨类植物有4科5属9种,其中有国家二级保护植物一种,即金毛狗(Cibotium barometz);被子植物43科70属97种,未发现裸子植物(表1)。群落组成的主要优势科有:樟科(Lauraceae),共6属14种;冬青科(Aquifoliaceae)1属7种;壳斗科(Fagaceae)3属6种,山矾科(Symplocaceae)1属5种,山茶科(Theaceae)有5属5种;有15科的物种数为2—4种,其余28科均为1属1种。
根据吴征镒种子植物属的分布区类型划分原则[10]进行统计,扣除去世界分布属后,该群落以热带分布区类型占绝对优势,共有56属,占非世界属总数的81.16%;温带性属共13属,占非世界属总数的18.84%。此外,该群落保存有较丰富的古老和原始科属,如壳斗科、木兰科(Magnoliaceae)、金缕梅科、山茶科等,原始的木本属有锥属(Castanopsis)、青冈属、木荷属(Schima)、含笑属(Michelia)、假蚊母树属(Distyliopsis)、柃木属(Schima)等。
3.2 群落垂直结构分析
该群落为南亚热带常绿阔叶矮林,以常绿阔叶树种占据绝对优势,落叶树种极少,主要有罗浮柿(Diospyros morrisiana)、山乌桕(Sapium discolor)和少叶黄杞(Engelhardtia fenzlii)等。因此,群落林冠层在夏季整体呈深绿色,秋冬季则有部分黄绿色斑块镶嵌其中。
根据群落的高度级频率分布(图1),该群落乔木层分为两个亚层,其中,第一亚层高10—18 m,约有20种66株,以鹿角锥(Castanopsis lamontii)、粗脉桂(Cinnamomum validinerve)、钝叶假蚊母树为主,其他树种主要有纤花冬青(Ilex graciliflora)、硬壳柯(Lithocarpus hancei)、密花树(Rapanea neriifolia)、深山含笑(Michelia maudiae)、烟斗柯(Lithocarpus corneus)、樟叶泡花树等;第二亚层高度5—10 m,共有39种380株,以钝叶假蚊母树、密花树、锈叶新木姜子(Neolitsea cambodiana)、黄丹木姜子(Litsea elongata)为主,其他树种主要有樟叶泡花树、鸭公树(Neolitsea chuii)、粗脉桂、密花山矾(Symplocos congesta)、大头茶(Gordonia axillaris)、大花枇杷(Eriobotrya cavaleriei)等。
灌木层高1—5 m,主要树种有细枝柃(Eurya loquaiana)、鱼骨木(Canthium dicoccum)、绒毛山胡椒(Lindera nacusua)、棱果花(Barthea barthei)等,还有较多的乔木幼苗,如钝叶假蚊母树、锈叶新木姜子、大花枇杷、黄丹木姜子、密花树等。该层植物种类丰富,共有种群59种534株,占林木层种株数的54.49%。整体上看,该群落灌木层和乔木第二亚层物种较为丰富,共有植物914株,占林木层总株数的93.32%。乔木第一亚层物种较少,且植株高度多在10—12 m左右。
群落草本层物种也较丰富,种群有68种共计478株,以流苏贝母兰(Coelogyne fimbriata)、紫花短筒苣苔(Boeica guileana)、华山姜(Alpinia chinensis)居多,还有较多的钝叶假蚊母树幼苗以及少些黄丹木姜子、光叶海桐(Pittosporum glabratum)、映山红(Rhododendron simsii)等乔灌木的幼苗。蕨类植物主要有中华复叶耳蕨(Arachniodes chinensis)、毛蕨(Cyclosorus interruptus)、乌毛蕨(Blechnum orientale)等。此外,还有国家二级保护植物金毛狗2株。藤本植物主要有石柑子(Pothos chinensis)、粉背菝葜(Smilax hypoglauca)、香花崖豆藤(Millettia dielsiana)、暗色菝葜(Smilax lanceifoliavar.opaca)、菝葜(Smilax china)、毛蒟(Piper hongkongense)等。

表1 深圳大鹏半岛钝叶假蚊母树群落种类组成Tab.1 Species composition in Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen
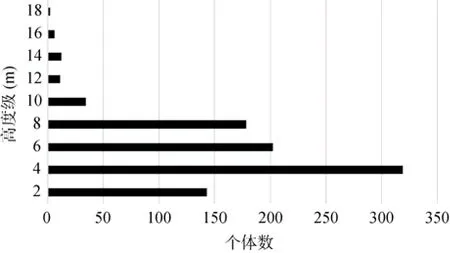
图1 深圳大鹏半岛钝叶假蚊母树群落高度级频率分布图Fig.1 The heightclassfrequency distribution of Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen
3.3 优势种群和重要值分析
对群落乔木层、灌木层、草本层物种的重要值进行计算,结果见表3-5。乔木层重要值大于1.00的植物共19种,其中以钝叶假蚊母树的重要值最高,为15.20,鹿角锥次之,重要值为10.13,密花树的重要值也达到9.62。灌木层重要值大于1.00的植物共21种,其中细枝柃的重要值最高,为19.02,鱼骨木次之,为11.57,棱果花的重要值也达到11.20。草本层重要值大于1.00的植物共21种,其中以流苏贝母兰的重要值最高,为16.88,紫花短筒苣苔次之,重要值为9.69,草本层分布有较多的钝叶假蚊母树小苗,其重要值为7.11。

表3 深圳钝叶假蚊母树群落乔木层主要物种的重要值Tab.3 Important value(IV)of tree layer dominant population in Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen

表4 深圳钝叶假蚊母树群落灌木层主要物种的重要值Tab.4 Important value(IV)of shrub layer dominant population in Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen

表5 深圳钝叶假蚊母树群落草本层主要物种的重要值Tab.5 Important value(IV)of herb layer dominant population in Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen
3.4 群落频度分析
按照Raunkiaer频度定律分析方法,对深圳大鹏半岛钝叶假蚊母树群落进行物种频度分析,并与Raunkiaer标准频度级进行比较,结果如图2所示。与Raunkiaer标准频度级相比,深圳钝叶假蚊母树群落的A级频度级与B级频度级所占比例更高,分别为60.34%和25%。整体而言,该群落频度分布为A>B>C<D>E,与Raunkiaer标准频度定律A>B>C≥D<E相似,但A、B频度级物种所占比例偏高,C、D、E频度级物种所占比例偏低,表明该群落偶见种较多。
3.5 乔木层优势种群的年龄结构分析
种群的年龄结构是种群内不同年龄个体数量的分布情况[11]。种群年龄结构能反映种群在时间和空间上的变动规律,体现种群动态及其所在群落的演替趋势[12],也能一定程度上反映种群与环境间的相互关系,说明种群在群落中的作用和地位[4]。在深圳大鹏半岛钝叶假蚊母树群落中选取乔木层重要值较大的钝叶假蚊母树、鹿角锥、密花树、粗脉桂和锈叶新木姜子等5种优势种群进行种群年龄结构分析(图3)。针对钝叶假蚊母树、密花树和锈叶新木姜子等三种灌木或小乔木种群,再用立木径级代替5级立木标准划分种群年龄结构(图4)。

图2 深圳大鹏半岛钝叶假蚊母树群落的频度级与Raunkiaer标准频度级对比分析Fig.2 Comparison analysisbetween frequency of Raunkiaer and Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen
由图4可以看出,乔木层中重要值最高的钝叶假蚊母树种群以Ⅲ级壮树最多,占40.82%,Ⅱ级小树次之,占25.71%,Ⅰ级苗木较少,为17.55%,无Ⅴ级老树。若按照5级立木标准划分年龄结构,可认为群落中的钝叶假蚊母树种群处于较为稳定、成熟的状态。但由于钝叶假蚊母树属于小乔木树种,样地内钝叶假蚊母树的胸径在0—14 cm的范围内,因此采用5级立木标准划分年龄结构会有所偏差。若按照径级结构划分,由图4可以看出,胸径在0—2 cm范围内的钝叶假蚊母树最多,占38.78%,整体径级结构呈现出“金字塔”型结构。表明群落中的钝叶假蚊母树种群处于发展阶段。

图3 深圳大鹏半岛钝叶假蚊母树群落乔木层主要优势种年龄结构示意图Fig.3 Age forum of tree layer dominant species in Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen
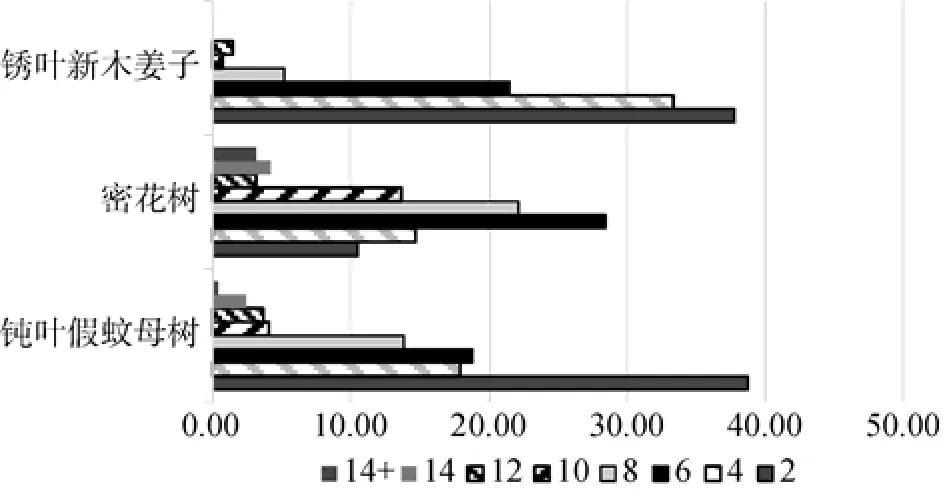
图4 深圳大鹏半岛钝叶假蚊母树群落乔木层主要优势种径级结构Fig.4 The DBH class structure of tree layer dominant speciesin Distyliopsistutchericommunity in Dapeng peninsula Nature Reserve,Shenzhen
鹿角锥种群以Ⅳ级大树居多,占41.18%,Ⅱ级小树次之,占23.53%,Ⅴ级老树占14.71%,Ⅰ级苗木仅有2.94%。表明群落中的鹿角锥种群处于成熟阶段。而密花树种群则以Ⅲ级壮树最多,占58.95%,Ⅳ级大树次之,为28.42%。若按照径级结构划分(图4),群落中密花树的胸径在0—17 cm的范围内,并且胸径在4—6 cm范围内的密花树最多,占28.42%,胸径6—8 cm的密花树数量次之,占22.11%,胸径>8 cm的密花树较少。因此可认为,群落中密花树种群处于趋向稳定、成熟阶段。粗脉桂的种群年龄结构总体上与密花树相似,呈现出“倒金字塔”型,与密花树种群相比,粗脉桂种群的Ⅲ级壮树比例较少,且有少量的Ⅴ级老树,可认为群落中的粗脉桂种群处于成熟并趋向衰退阶段。锈叶新木姜子种群以Ⅲ级壮树最多,占48.89%,Ⅱ级小树次之,占45.93%,Ⅰ级苗木和Ⅳ级大树均少于5%,无Ⅴ级老树。结合该种群的径级结构分布,群落中锈叶新木姜子的胸径在0—12 cm的范围内,并且胸径在2—4 cm范围内的锈叶新木姜子最多,占37.78%,胸径>6 cm的锈叶新木姜子占比不足10%,因此可认为,该种群处于发展阶段。
3.6 物种多样性分析
群落的物种多样性是一个群落结构与功能复杂性的度量[13],多样性指数和均匀度指数作为反映物种多样性的定量数值,对衡量群落演替、探讨群落的最优物种结构等方面具有重要意义,并且可以作为自然资源保护与开发的数量指标[14]。
目前国内关于钝叶假蚊母树群落的研究很少,关于钝叶假蚊母树种群及其所在群落特征的研究至今未见报道。因此,本研究选取地理位置相近的广东南岭大东山浙江润楠群落[15]进行比较研究。由表6可知,在物种多样性方面,深圳大鹏半岛钝叶假蚊母树群落的乔木层与灌木层在Simpson指数、Shannon-wiener指数和Pielou指数方面均稍低于南岭大东山浙江润楠群落,但其草本层的Shannon-wiener指数和Pielou指数则明显高于南岭大东山浙江润楠群落。
而在钝叶假蚊母树群落不同层次物种多样性变化方面,由表6可知,Simpson指数和Pielou指数表现为灌木层>乔木层>草本层,而Shannon-wiener指数则表现为灌木层>草本层>乔木层。总体上看,灌木层物种多样性和均匀度最高。
总体而言,与地理位置相近的其他群落相比,深圳大鹏半岛钝叶假蚊母树群落物种多样性较高,属于受人为干扰少,保存完好的南亚热带常绿阔叶林,群落各层次整体上符合自然生态系统各层次的物种多样性灌木层>乔木层>草本层的格局[16]。

表6 深圳大鹏半岛钝叶假蚊母树群落与其他植物群落的物种多样性指数比较Tab.6 Comparison of biodiversity index between the Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve and other communities

接上表
3.7 生活型分析
根据C.Raunkiaer的生活型分类系统[17],对深圳钝叶假蚊母树群落进行生活型分析,并与广东南昆山地区的森林群落进行对比,结果如下图所示(见图5),由图可知,该群落以高位芽植物种群为主,占群落植物总种数的70.08%,其中又以小高位芽植物最多,占40.17%;地上芽植物和地下芽植物较少,分别占5.13%、2.56%;地面芽植物和藤本植物较多,分别占14.53%、7.69%,无一年生草本和非自养植物。相较而言,深圳大鹏半岛钝叶假蚊母树群落的高位芽植物比例低于广东南昆山地区的森林群落(83.8%),特别是广东南昆山森林群落中高位芽植物比例(36.7%)明显高于深圳大鹏半岛钝叶假蚊母树群落(14.53%)。一方面表明其热带性气候、雨热条件充沛,中高位芽植物减少、小高位芽及矮高位芽植物增多,另一方面也与其位于山顶,高位芽植物种群出现矮化现象有关。另外,群落的藤本、地面芽植物比例较高,这与南亚热带海洋性季风气候区湿润多雨的气候相适应。
4 讨论
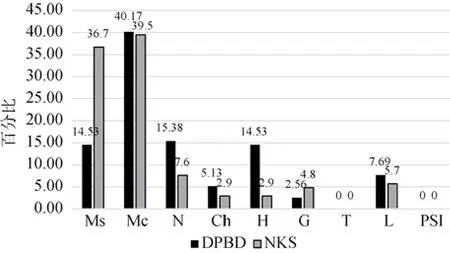
图5 深圳大鹏半岛钝叶假蚊母树群落生活型谱与邻近地区常绿阔叶林的比较Fig.5 Comparison oflife-form spectrum between Distyliopsis tutcheri community in Dapeng peninsula Nature Reserve,Shenzhen and other evergreen broad-leaved forest communities in the adjacent region
深圳大鹏半岛自然保护区钝叶假蚊母树群落有47科75属106种,林下草本和层间藤本植物较为丰富,物种多样性较高,是保存完好的南亚热带常绿落叶阔叶林群落。群落垂直结构分层明显,乔木下层物种丰富,乔木上层物种较少,这与群落主要优势种多为灌木或小乔木树种有关:一方面由于灌木层和乔木第二亚层包含了乔木第一亚层树种的幼苗;另一方面则表明该群落以灌木树种为主,占较大优势,这与该群落所处的海拔位置相关,钝叶假蚊母树群落位于排牙山顶附近,而排牙山三面濒海,常年海风较大,不利于高大乔木的生长,因此,较为低矮的灌木获得了竞争优势。群落地理成分分析结果表明,群落中物种组成以热带成分占绝对优势(81.16%),符合典型的亚热带山地性质,温带性属以东亚—北美间断分布属最多,体现出钝叶假蚊母树群落的物种组成具有一定的古老性。
优势种群年龄结构和重要值分析表明,该群落乔木层以钝叶假蚊母树重要值最高,且草本层也有较多钝叶假蚊母树幼苗分布,表明了该群落纯度高、分布集中、原始性强,此外,该群落的乔木层优势种较多,但优势度较不明显,各主要优势种互相竞争,群落较为稳定。Raunkiaer频度级分析表明大鹏半岛钝叶假蚊母树群落频度级分布规律为A>B>C<D>E,与Raunkiaer标准频度定律A>B>C≥D<E相比,该群落D级树种所占比例高于E频度级。D级为频度在61%—80%的物种,主要有大花枇杷、华山姜、鹿角锥、深山含笑等7种,而E级植物仅4种,其中钝叶假蚊母树和密花树的频度均为100%。结合该群落的种群重要值分析,钝叶假蚊母树和密花树在乔木层中占有较高重要值,优势度明显,而且从优势种群的年龄结构分析来看,频度级为D级的鹿角锥种群为成熟种群,而频度级为E级的钝叶假蚊母树种群和密花树种群均为增长种群,推测在与D频度级树种的竞争中,有较大竞争优势的E级树种钝叶假蚊母树和密花树抑制了D级物种的分布扩张。整体上看,该群落处于郁闭未稳定的亚顶级状态,群落结构趋于稳定,但群落内部仍有一定的动态变化过程。
物种多样性方面,钝叶假蚊母树群落物种多样性各项指数与广东南岭大东山浙江润楠群落相近,但其草本层物种相对较丰富。物种多样性各项指数首先与森林类型有关[18],南岭大东山浙江润楠群落属于典型的中亚热带常绿阔叶林,结构较为复杂,且处于演替的中期阶段,因此南岭大东山浙江润楠群落的物种多样性较高,但与深圳钝叶假蚊母树群落的各项指数相差不大,说明钝叶假蚊母树群落发展较好。而深圳钝叶假蚊母树群落的草本层在Shannon-wiener指数和Pielou指数相对较高,而Simpson指数较低,说明该群落草本层物种相对丰富,且分布均匀,但优势种较少。这也表明,相比大东山浙江润楠群落,钝叶假蚊母树群落处于演替的较早期阶段。而从群落各层次比较上看,该群落灌木层物种多样性最高,一方面是由于群落主要优势种多为灌木树种,乔木树种较少;另一方面,灌木层不仅有灌木树种,还包括乔木树种的幼树。由于Shannon-Wiener指数与丰富度关系较为密切[19],因此在灌木树种占优势的群落,乔木层物种丰富度低,Shannon-wiener指数较低。草本层的Simpson指数和Pielou指数均为最低,但与乔木层和灌木层的差别不大。群落各层次的物种多样性是相互依赖的[20],并且森林的特性及动态特点有关[21],在受干扰较小的常绿阔叶林中,由于群落郁闭度较高,林下光照不足,会导致草本层物种丰富度的降低。而由于群落位于排牙山顶,不利于高大乔木的生长,因此乔木树种较少。结合从生活型分析来看,群落以高位芽种群为主,其中又以小高位芽种群最多,缺少大高位芽植物,有相对较多的地面芽植物和藤本植物,表明该群落将向更稳定的中期阶段发展。
总体而言,深圳大鹏半岛钝叶假蚊母树群落分布较为集中,受非自然因素干扰较少,是保存完好的南亚热带常绿阔叶林资源,对进一步开展钝叶假蚊母树群落相关研究有十分重要的学术价值和生态学价值,建议保护区加强对该群落的管理和保护。
致谢:本项目得到了深圳市野生珍稀濒危重点保护植物调查项目(71020140)、深圳市级自然保护区监测项目(71021056)、广东省野生动植物保护管理项目资助(42090393)。此外,深圳市野生动植物保护管理处胡平,中山大学植物学专业本科生刘佳、朱晓枭,研究生赵万义、阴倩怡、许可旺、关开朗等参加了野外工作,特此鸣谢!
参考文献
[1]张宏达.中国金缕梅科植物订正[J].中山大学学报(自然科学版),1973(1):68–71.
[2]梅兴国,陈钧鸿.钝叶水丝梨化学成分研究[J].植物科学学报,1989,7(2):179–187.
[3]邢福武,陈红锋,王发国等.南岭植物物种多样性编目[M].武汉:华中科技大学出版社,2011,114–115.
[4]王伯荪,余世孝,彭少麟.植物群落学实验手册[M].广州:广东高等教育出版社,1996:1–56.
[5]JIN Z.A Study of Population Structure and Distribution Pattern ofHeptacodiummiconioidesin the Tiantai Mountain,Zhejiang[J].Chinese Journal of Ecology,1997.
[6]张志祥,刘鹏,刘春生,等.浙江九龙山南方铁杉(Tsuga tchekiangensis)群落结构及优势种群更新类型[J].生态学报,2008,28(9):4547–4558.
[7]达良俊,杨永川,宋永昌.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报,2004,28(3):376–384.
[8]ROBERT P.Mclntosh.Raunkiaer’s “Law of Frequency”[J].Ecology,1962,43(3):533–535.
[9]孙儒泳,李博,诸葛阳,等.普通生态学[M].北京:高等教育出版社,1996.136–137.
[10]吴征镒.中国种子植物属的分布区类型[J].植物分类与资源学报,1991(S4).
[11]张育新,马克明,祁建,等.北京东灵山海拔梯度上辽东栎种群结构和空间分布[J].生态学报,2009,29(6):2789–2796.
[12]MASAKI T,OTA T,SUGITA H,et al.Structure and dynamics of tree populations within unsuccessful conifer plantations near the Shirakami Mountains,a snowy region of Japan[J].Forest Ecology&Management,2004,194(1):389–401.
[13]谢晋阳,陈灵芝.暖温带落叶阔叶林的物种多样性特征[J].生态学报,1994,(4):337–344.
[14]彭少麟,王伯荪.鼎湖山森林群落分析Ⅰ.物种多样性[J].生态科学,1983(1):11–17.
[15]胡晓敏,董安强,王发国,等.广东南岭大东山浙江润楠群落物种多样性与区系地理成分分析[J].植物科学学报,2011,29(3):265–271.
[16]贺金生,陈伟烈,李凌浩.中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J].植物生态学报,1998(4):303–311.
[17]朱忠保.森林生态学[M]北京:中国林业出版社,1991.33–40.
[18]黄忠良,孔国辉,何道泉.鼎湖山植物群落多样性的研究[J].生态学报,2000,20(2):193–198.
[19]马克平,黄健辉,于顺利,等.北京东灵山地区植物群落多样性的研究[M].生态学报,1995,16(3):225–234.
[20]BRADFIELD G E,SCAGEL A.Correlations among vegetation strata and environmental variables in subalpine spruce-fir forests, southeastern British Columbia[J].Vegetatio,1984,55(2):105–114.
[21] AUCLAIR A N, GOFF F G. Diversity relations of upland forests in the western Great Lakes area[J]. American Naturalist, 1971, 105(946): 499–527.
