深圳市内伶仃岛山蒲桃+红鳞蒲桃-小果柿群落结构及其物种多样性特征
2018-05-14李薇朱丽萍汪春燕吴荣恩刘佳凡强
李薇,朱丽萍,汪春燕,吴荣恩,刘佳,凡强,*
1.中山大学生命科学学院,有害生物防治与资源利用国家重点实验室/广东省热带亚热带植物资源重点实验室,广州 510275 2.深圳市规划国土发展研究中心,深圳 518040 3.华南农业大学华南农业博物馆,广州 510642
1 前言
广东省深圳市内伶仃岛位于珠江口,地理位置独特,生物资源较为丰富,岛屿植被主要以次生常绿阔叶林、常绿灌丛为主,并保存有一定面积的典型南亚热带季风常绿阔叶林。1988年内伶仃岛被列入国家级自然保护区,是深圳进行生态保护的重点区域,且位于深圳市基本生态控制线范围内。在历年的调查中已见岛上的植被得到了良好的生态保护,植被逐渐恢复[1],植被覆盖率达90%以上[2]。天然林群落的建群种、优势种主要有:布渣叶(Microcos paniculata)、蒲桃(Syzygium jambos)、翻白叶树(Pterospermum heterophyllum)、马尾松(Pinus massoniana)、白桂木(Artocarpus hypargyreus)、山蒲桃(Syzygium levinei)、红鳞蒲桃(S.hancei)、短花序楠(Machilus breviflora)等。
1997-1999年,广东内伶仃福田国家级自然保护区组织了针对内伶仃岛自然资源的综合考察,记述了在尖峰山的红鳞蒲桃群落[3],但未有深入研究该群落的特征。此次调查的目标群落为山蒲桃+红鳞蒲桃-小果柿(Diospyros vaccinioides)群落,其中,山蒲桃、红鳞蒲桃隶属于桃金娘科(Myrtaceae),小果柿隶属于柿科(Ebenaceae),3种均为优良乡土树种。小果柿亦为中国特有种,只见于香港、台湾及广东省,多见分布于珠江口的岛屿上[4];该种亦被世界自然保护联盟(IUCN)列入物种红色名录——据其数量和分布定义为极危(critically endangered)级别[5],面临着灭绝的威胁。另外,小果柿树形秀美,适于作盆景栽种,是园艺造景的上佳造材,因而也遭受了滥采乱挖,近年野生种群已逐渐减少。本次试图通过对山蒲桃+红鳞蒲桃-小果柿群落的调查研究,探讨内伶仃岛的原生环境,为制定适当的保护管理方案提供依据。
2 研究区域与样地设置
2.1 内伶仃岛概况
广东内伶仃岛地处珠江口伶仃洋东部,总面积4.98 km2, 地理位置为北纬 22°23¢49¢—22°25¢35¢¢,东经 113°46¢18¢—113°49¢49¢。内伶仃岛西距珠海约24 km,北距深圳蛇口约16 km岛上全年高温湿热,年均气温22.0—22.4℃,年均降雨约2000 mm,有明显的干、湿季之分,降水集中在4—9月,日照年总量约2000 h。内伶仃岛为丘陵地貌,总体地势呈中间高,四周低,主峰尖峰山海拔340.9 m。土壤母岩由花岗岩、变质砂岩构成;地带性土壤主要为赤红土、滨海沙土与耕作土。淡水资源主要由6条季节性地表径流构成。全岛共有野生维管植物133科379属569种[6]。内伶仃岛的原生植被自上个世纪60年代以来已遭破坏,目前恢复的主要是次生性的南亚热带常绿阔叶林[1,6–7]。岛上常见优势树种有蒲桃属(Syzygium)、潺槁(Litsea glutinosa)、白桂木、短序润楠、鸭脚木(Schefflera octophylla)。除常绿阔叶林外,全岛灌木林也占有相当的面积,其间覆盖了大量藤本植物,成为全岛一个主景观,主要种类有飞龙掌血(Toddalia asiatica)、龙须藤(Bauhinia championii)、刺果藤(Byttneria aspera)等[2]。
2.2 样地调查
样地设于尖峰山东南坡,经纬度为22°24¢34.70¢N、113°48¢24.05¢E, 海拔325 m, 总面积2400 m2,再划分成10 m×10 m方格,采用每木记帐调查法对样地进行调查,乔灌木起测高度1.5 m,记录种名、胸径、树高、冠幅等。每方格再设置1个2 m×2 m 小样方,记录低矮灌木、草本植物,以及乔灌木的幼苗等。
1.3 数据分析
(1)重要值。重要值=(相对显著度+相对多度+相对频度)/3,其中藤本植物在样地中的相对显著度即为相对盖度。
(2)物种多样性:采用丰富度指数、Simpson指数、Shannon-Wiener多样性指数和以Shannon-Wiener指数为基础的Pielous均匀度指数。即:

(3)优势种群的径级(DBH)结构[9]:I级幼苗:H≤33 cm;II级幼树:H>33 cm,DBH<2.5 cm;III级成树:2.5 cm≤DBH<7.5 cm;IV级壮树:7.5 cm≤DBH<22.5cm;V级成龄老树:DBH≥22.5 cm。
(4)频度分析:采用RAUNKIAER频度定律,频度在1%—20%的植物种归入A级,21%—40%为B级,41%—60%为C级,61%—80%为D级,81%—100%为E级。
3 结果与分析
3.1 群落外貌与组成结构
(1)外貌与分层结构。选定的山蒲桃+红鳞蒲桃-小果柿群落位于主峰尖峰山,接近山顶,面向东南部的迎风坡处,群落外貌葱郁浓密,青绿色,郁闭度达80%—92%,林冠层起伏明显,稍连续,冠层植株分枝较多。乔木层可分2个亚层,第一亚层高12—17 m,优势种主要山蒲桃、红鳞蒲桃、红枝蒲桃等,其它常见种有短序润楠、鸭脚木、笔管榕(Ficus superba);第二亚层分层极不明显,成层现象不显著,为3—11 m,种群结构复杂,物种组成丰富,林内植株分枝较密,优势种主要有、白背槭(Acer decandrum)、小果柿、布渣叶、芳槁润楠(Machilus suaveolens)等,其它常见种有假苹婆(Sterculia lanceolata)、山杜荆(Vitex quinata)、翻白叶树(Pterospermum heterophyllum)、刺葵(Phoenix hanceana)等。灌木层种类分层较明显,主要有小果柿、豺皮樟(Litsea rotundifolia)、九节(Psychotria rubra)、轮叶木姜子(Litsea verticillata)、大叶紫珠(Callicarpa macrophylla)、大叶桂樱(Laurocerasuszippeliana)、紫玉盘(Uvaria microcarpa)、白楸(Mallotus paniculatus)等,多为南亚热带常绿阔叶林下常见种群。草本层种类较少,林下层郁闭度大,达90%—97%,因缺少阳光而覆盖率低,主要有华山姜(Alpinia chinensis)、半边旗(Pteris semipinnata)、三叉蕨(Tectaria subtriphylla)等。层间植物丰富,大型藤本长势茂密,攀附、绞杀现象明显,常严重遮蔽林层空间,主要种类有广州槌果藤(Capparis cantoniensis)、刺果藤、白藤(Calamus tetradactylus)、罗浮买麻藤(Gnetum lofuense)、飞龙掌血等。
(2)群落物种组成。在2400 m2的样方中,记录维管植物共102种,隶属80属46科,其中蕨类植物6科7属8种,裸子植物仅1科1属1种,被子植物39科71属93种,与全岛相比较科属种所占比例分别为34.59%、21.11%、17.93%。主要优势科有:樟科(Lauraceae)(7属/9种,下同)、大戟科(Euphorbiaceae)(6/7)、番荔枝科(Annonaceae)(3/5)、芸香科(Rutaceae)(6/6)、卫矛科(Celastraceae)(2/4)、茜草科(Rubiaceae)(3/3)、马鞭草科(Verbenaceae)(2/5)、棕榈科(Palmae)(2/3)、桃金娘科(1/3)等。各数量优势科在植被群落中亦占有一定位置,或为优势种群所属科。如樟科、芸香科、茜草科、卫矛科等亦是华夏植物区系的表征科[2],而棕榈科更为热带与亚热带植被的指示性科,群落具强烈的南亚热带植被特征。层间藤本植物丰富,一是出现于群落内成为常绿阔叶林重要热带性特征,二是由于内伶仃岛海拔不高,常风较大,使林缘、灌丛中形成大面积的藤本群落,亦成为热带、亚热带森林的一个主要外貌结构特征[10]。
(3)群落种类组成的地理成分性质。蕨类植物共有5科,均为世界广布科[11],如鳞毛蕨科(Dryopteridaceae)、海金沙科(Lygodiaceae)、凤尾蕨科(Pteridaceae)等,说明蕨类植物并未出现特征性的种群。种子植物共出现40科,其中,世界广布科9科,从蕨类植物广布科、种子植物广布科看,尖峰山的该群落具有明显的次生性。整体上,种子植物的热带性科[12]占绝对优势,占非世界科总数的93.55%,如棕榈科、樟科、番荔枝科、山茶科(Theaceae)等。温带性科仅有2科,为冬青科(Aquifoliaceae)、槭树科(Aceraceae)。

表1 山蒲桃+红鳞蒲桃-小果柿群落植物种类组成及生态型Tab.1 Species composition and ecologic types of Sygyzium levinei+S.hancei-Diospyros vaccinioides community
从属级水平分析,参照中国种子植物属的分布区类型[13]进行统计,热带成分共61属,占非世界属总数的88.40%,泛热带分布属数最多,如厚壳桂属(Cryptocarya)、苹婆属(Sterculia)、九节属(Psychotria)、柿属、刺果藤属(Byttneria);其次为旧世界热带分布,如蒲桃属、八角枫属(Alangium)、紫玉盘属(Uvaria)、大沙叶属(Pavetta)等;热带亚洲至大洋洲分布的瓜馥木属(Fissistigma)、海芋属(Alocasia)、银柴属(Aporusa)等;热带亚洲分布的如润楠属(Machilus)、山茶属(Camellia)、省藤属(Calamus)、破布叶属(Microcos)等。温带成分稀少,为5属,占11.60%,如忍冬属(Lonicera)、荚蒾属(Viburnum)、槭属(Acer)等。种类组成的地理成分统计表明,该群落受到热带成分的强烈影响,处于南亚热带向热带扩散的边缘带。
3.2 优势种群和群落结构特征分析
(1)重要值。根据群落垂直结构特性,分层计算各物种的重要值[9](表3),根据重要值原则可将群落命名为“山蒲桃+红鳞蒲桃-小果柿”群落。整体看来,山蒲桃种群占有较大的优势,其它种群如红鳞蒲桃、小果柿、短序润楠、刺葵、红枝蒲桃、白背槭等其优势度相近,表明该群落是一个多优势种群落,从种群性质以及存在3种蒲桃属植物和刺葵来看,其热带性特征明显,并且也是南亚热带代表性群落退居山顶的一种孑遗类型。群落中最主要的特征种为小果柿,种群长势优良,其在乔木层有101株,灌木层有189株,种群相对显著度小而相对多度大,显示灌木枝干细长而多分株,生长年限长。其它几个优势种重要值相近,表明种间竞争剧烈。

表2 山蒲桃+红鳞蒲桃-小果柿群落种子植物科属的分布区类型组成Tab.2 The Family and generic areal-types of seed plants in Sygyzium levinei+S.hancei-Diospyros vaccinioides community

接上表
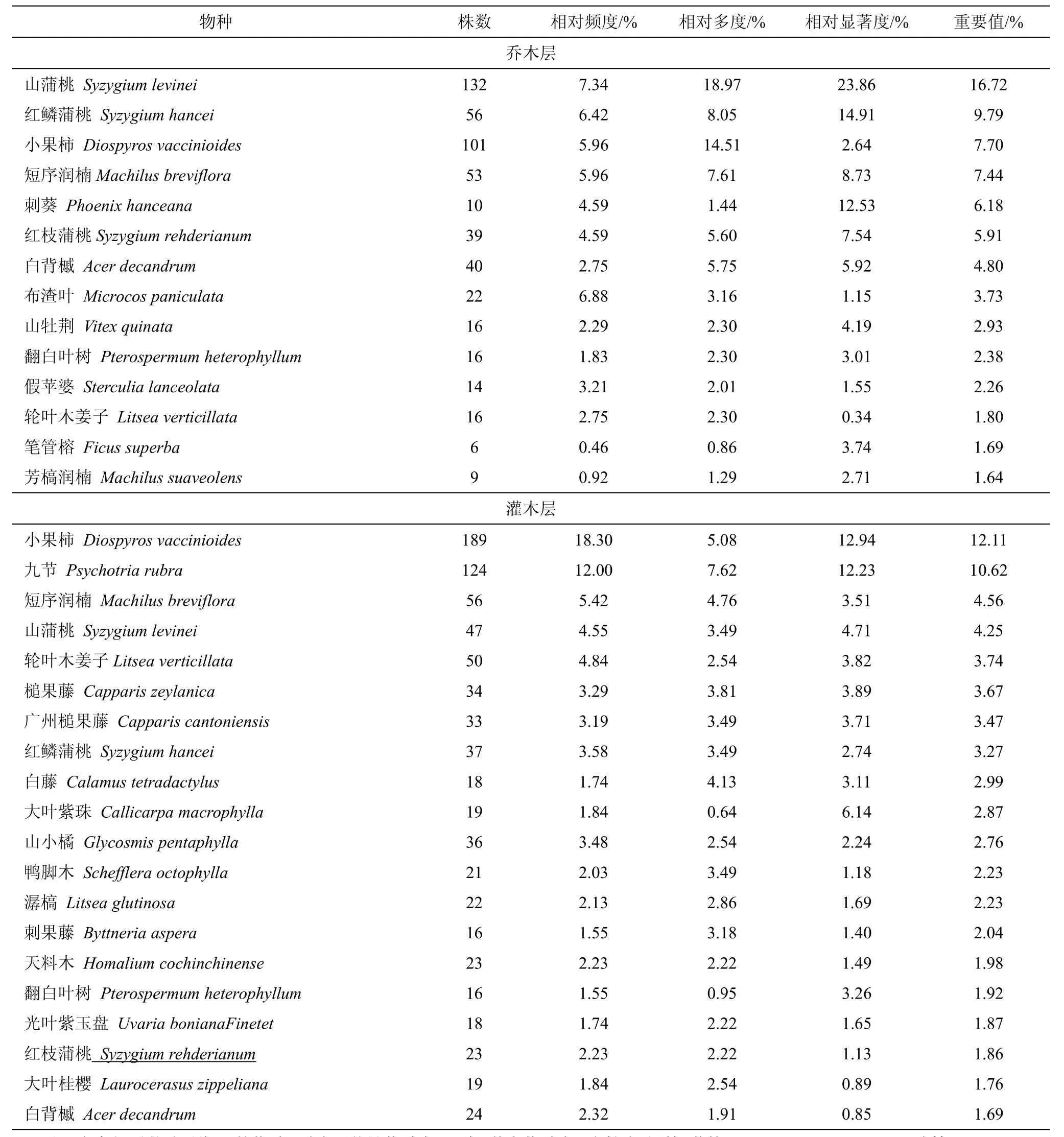
表3 山蒲桃+红鳞蒲桃-小果柿群落乔灌层主要物种的重要值Tab.3 Important value(IV)of dominant populations in Sygyzium levinei+S.hancei--Diospyros vaccinioides community
(2)生活型分析。群落中各物种的生活型谱,可 反映出当地环境因子对植物生态习性的影响。长期趋同演变和适应的结果下,使得具有不同基因型的植物,长期在同一环境条件生长下形成相似的外貌、结构和生理特征[14–15]。参照RAUNKIAER的分类等级划分[14],统计该群落的植物生活型谱如表4所示,群落以高位芽植物占主导地位,其中又以小高位芽植物最丰富,38种占37.26%,以乔木亚层植物与林下常见灌木为主,如小果柿、九节、大叶紫珠等;其次为中高位芽25种占24.51%,以乔木层主要优势种为主;地上芽、地面芽与一年生草本种类极少,如山麦冬(Liriope spicata)、碎米莎草(Cyperus iria)等;藤本植物共11种,皆为高位芽植物,如崖豆藤属(Millettia)与刺果藤属。常绿的小高位芽植物占优,地上芽与地面芽植物缺少,木质藤本高位芽植物生长茂盛,一定程度上反映出该群落具有温暖潮湿的气候特征,据本调查结果可见群落植被与内伶仃岛的湿热气候特性吻合。一般在气候高温潮湿的地区,反映高位芽植物占比相对较大,而地上芽与地面芽植物较少[16],随着纬度北移移或海拔的增加气候趋向寒冷干旱,则表现为高位芽植物减小,但地上芽植物与隐芽植物增多[17]。
(3)乔木层优势种群的年龄结构
群落主要优势种群的年龄结构,可在一定程度上反映种群动态及其演替趋势。根据五级立木径级结构划分年龄段,山蒲桃+红鳞蒲桃-小果柿群落主要优势种群的年龄结构如图1所示。
图1表明,山蒲桃、红鳞蒲桃、短花序楠、红枝蒲桃的径级结构大致呈壮年期时期,II、III、IV级立木所占比例较大。山蒲桃种群结构为倒金字塔型,IV级树种所占比例最大,III、II、I级立木依次减少,显示该种群为衰退型种群。红鳞蒲桃的种群结构在本群落中是最显著的,V、IV、III、II级立木大致相当于金字塔形,种群增长减缓,趋向稳定期,尽管I级立木较少,但仍是能够适应该群落,整体显示长期内红鳞蒲桃将仍然为优势种。短花序楠IV、III、II级立均较多,但V、I级很少,应该是属于稳定型种群。小果柿IV、V级苗木缺失,而II、III级占绝大部分,少量I级幼苗,种群发展潜力大。刺葵属棕榈科植物,是一个特殊的种群,按径级分级看,种群IV级成树占有极高比例,V级缺失,III、II贫乏,I级幼树也缺失,重要值虽在群落中占据较前位置,但明显为衰退型种群,在演替中将逐渐衰退。而白背槭种群呈典型J型,V、IV、III、II、I级依次增加,为增长型种群,并且I级幼树占48%,种群处于快速上升阶段,未来在群落中的优势度会持续增加,或发展为群落中的优势种群之一。

表4 山蒲桃+红鳞蒲桃-小果柿群落物种生活型谱组成Tab.4 Life form spectrum of Sygyzium levinei+S.hancei-Diospyros vaccinioides community
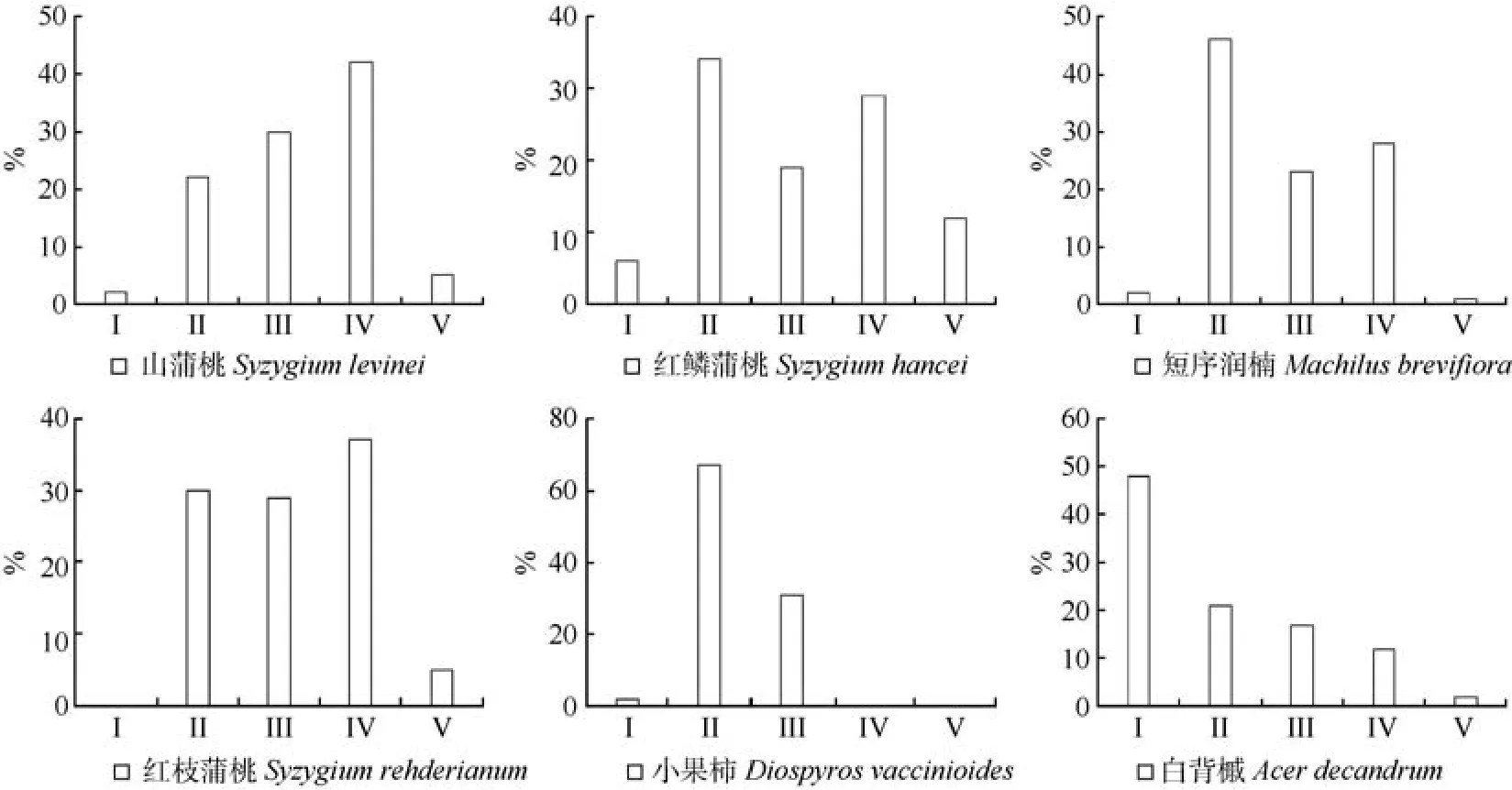
图1 山蒲桃+红鳞蒲桃-小果柿群落优势种群的年龄结构图Fig.1 Age-structure of dominant populations in Sygyzium levinei+S.hancei-Diospyros vaccinioides community
(4)小果柿种群的高度级结构
小果柿在本群落中较为特殊,不仅占据乔木层部分空间,亦是灌木层优势度最大的种群,在深圳区也是绝无仅有,特殊少见。小果柿在习性上仍为灌木,现按高度级分析其垂直结构以补充分析种群年龄结构。小果柿种群的高度级按0.5 m的间隔划分。如图2所示,群落中小果柿种群的优势度主要集中在1.0—4.5 m间,最高可至8 m,整体看来具有增长的趋势,是一个适应性较强的种群。小果柿种群分株多而枝径细长,或为获得更多阳光与空间而促使其向高生长。不同生境中种群呈现各异的结构与特征,被认为是物种在特定环境长期演替而表现的适应性,是一种特定的适应性进化现象[18]。
3.3 频度级与物种多样性特征
(1)频度级。基于种群多度的RAUNKIAER频度定律可用于分析群落中种群分布的均匀度,频度与种群个体在群落中分布的均匀程度呈正相关。按照RAUNKIAER的频度定律,一般各频度级的规律为:A>B>C≧D
山蒲桃+红鳞蒲桃-小果柿群落频度级结构见图3。A级频度比例为68%,B级为17%,C级与D级相等,皆为7%,E级最小,仅占1%。即本群落的频度规律为:A>B>C=D>E,与标准频度规律有差异。A级与E级频度呈两极分化,A级相当高,显示群落中有较多的偶见种,群落的物种多样性倾向于集群分布,E级所占极小,显示频繁出现的物种并不多,群落均匀度较小,优势种不明显,或处于多优势种的活跃发展阶段。还有一个原因,这与该样地接近山顶、坡度较陡有一定关系,地貌上的差异使部分种群生长受到限制,从而导致物种的分布不均匀。
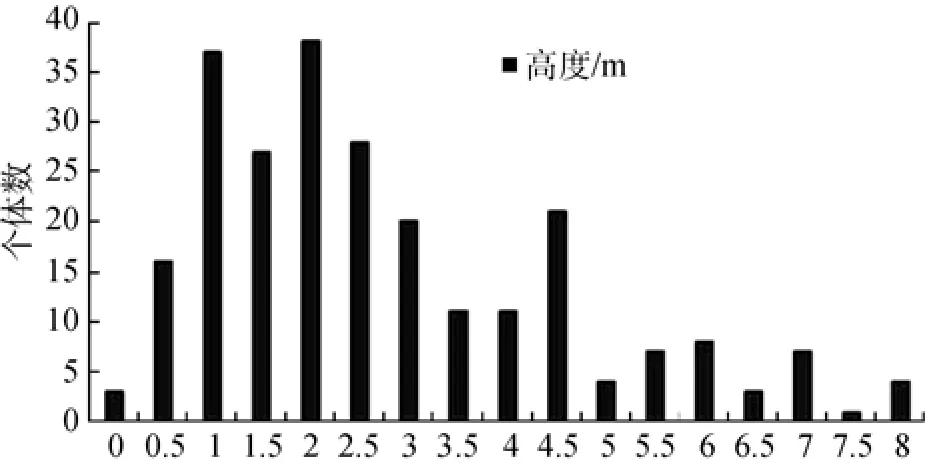
图2 山蒲桃+红鳞蒲桃-小果柿群落小果柿种群高度级结构图Fig.2 Individual number at height classes of Diospyros vaccinioides in Sygyzium levinei+S. hancei-Diospyros vaccinioides community

图3 山蒲桃+红鳞蒲桃-小果柿群落种群的频度级图解Fig.3 Individual number of frequency classes in Sygyzium levinei+S.hancei-Diospyros vaccinioides community
(2)物种多样性指数。计算群落Shannon多样性指数H′=3.64,Simpson指数D=0.95,Pielou均匀度指数Jsw=0.79,反映群落多样性较高,而种群分布均匀程度一般;群落中绝对优势种不明显,各物种多度差异较小,为多个优势种形成的共建群落。广东南亚热带常绿阔叶林的物种多样性指数(Shannon-Wiener指数)在4—5间,均匀度指数在0.7—0.8间[19–20]。与邻近地区群落的物种多样性对比,从表5来看,内伶仃岛的山蒲桃+红鳞蒲桃-小果柿群落物种多样性的各指数,与深圳东部山地常绿阔叶林相当,但相较天然保存良好的原生林多样性指数仍偏低。据文献描述,香港黄桐林为香港的地带性的原生植被,保存良好,可见多样性指数高于本群落。而群落Margalef丰富度指数为13.32,物种丰富度指数偏低,这与内伶仃岛属于孤岛,和大陆有地理隔绝有关;同时岛屿也都曾受到一定程度的干扰,使得岛上群落物种多样性低于原生性的亚热带常绿阔叶林。人类活动及原生生境对植物种群的影响是非常明显的,过度的人类活动将会严重干扰大多数极小种群、各类珍稀濒危种群、或特有种群的自我生长和演替。
4 结论与讨论
(1)关于山蒲桃+红鳞蒲桃-小果柿群落的特征。该群落共有维管植物共46科80属102种,主要优势科多为华夏区系表征科,如樟科、茜草科、番荔枝科、桃金娘科等;从种类组成的地理区系成分看,热带性科占93.55%,热带性属占88.40%,群落主要建群种为山蒲桃、红鳞蒲桃和短序润楠,灌木层优势种为小果柿、九节等,层间藤本植物生长茂密,整体上以热带成分占绝对优势,显示着较强的从南亚热带向热带常绿阔叶林过渡的特征。
关于群落内的种群竞争,小果柿也展示了其特殊性。作为低矮的阳生性树种,小果柿间断分布于台湾、海南与香港、深圳。在以往的群落调查中,小果柿多呈亚灌木状,并且是零散分布[26]。本群落中,小果柿在乔木第二亚层,与其它乔木竞争空间和阳光,因而向高生长旺盛,乔木层统计个体数共101株,其中高度4—5 m有38株,高度5—8 m共有50株,种群与乔木争夺群落空间位置;小果柿属喜阳树种,群落的高郁闭度或促使种群向高层生长,占据上层乔木层优良生态位;灌木层共统计189株,其中高度1—3 m有179株,种群处于良好发展态势。
(2)关于优势种群的相互竞争和生态位重叠。本群落中,蒲桃属就有3个优势种群,该属为旧世界热带分布属,是在内伶仃岛天然林中保存较好的、仅存的、孑遗性群落。出现同属3种而不是仅1种,体现了群落中物种多样性的演替规律总是以趋于多样化、复杂化为先导,这是一种本质属性。从竞争的程度看,山蒲桃、红鳞蒲桃、红枝蒲桃均处于旺盛生长阶段,在亲缘上他们同一属是相近的,因此对生态环境的要求、习性偏好也相近,其生态位多少出现重叠,因而生长旺盛而竞争激烈。竞争的结果,已呈现一定的趋势,即山蒲桃是一个衰退型种群,红枝蒲桃是一个短期内处于稳定状态的种群,衰退尚不明朗,而红鳞蒲桃种群则从增长型向稳定型过渡,可见三者间确实竞争激烈。群落内3个种群的个体大致趋向集群分布,均匀度低,群落处于剧烈的动态发展阶段。根据南亚热带森林群落研究的结果获知[20],随着演替的深入,群落的组成和结构将趋向于更加复杂和稳定。因而,该群落蒲桃属3种的演替关系尚不能一概而论。影响演替趋势的要素要归结于两方面:一是物种数量的多少,二是物种间相互作用的大小[14]。从蒲桃属3种的种群格局看,很难判断谁将最终被淘汰,目前的共优势种局面仍会延续,因此,该群落可定义为活跃的、相对稳定的、处于动态平衡的发展阶段。
(3)关于岛屿植被保护和生态恢复。内伶仃岛是一个大陆性岛屿,四面环海处于一个相对封闭的区域,加之耸立珠江口伶仃洋,常风较大,热量、雨量充沛,自世纪80年代建立自然保护区后,制定了封山育林、加强保护的管理方案。目前,全岛已禁止一切开发和经营活动,除保护区常驻人员、边警小组外,已撤出常住居民,果园也已退果还林,各类农用地、菜园已弃耕。因而,全岛植被恢复良好,天然次生林得到正常演替,其它人工林也开展逐渐得到改善。本群落近山顶,是该区域保护最好的常绿阔叶林群落。
(4)生态控制线下缘的生态修复。整个内伶仃岛均位于基本生态控制线范围内,近30年来群落与植被保护良好,次生常绿阔叶林得到进一步发展。而在生态控制线下缘,以及环岛公路上下10—30 m范围,受人类活动影响,天然植被几乎已被破坏,代之是在人工次生林的基础上发育、恢复形成的片段化植被。未来应根据具体情况,进行人工干预和生态修复,其一可以广泛地种植当地适生的乡土树种,增加群落的物种多样性,其二有计划地清理外来种,清理不合适的栽培种,扩大生态保护区域范围,全面构建和恢复全岛的天然林、次生林植被。
致谢:深圳市城管局公园管理中心孙延军高工,中山大学植物学专业本科生朱晓枭,研究生阴倩怡、许可旺、关开朗等参加了野外样地调查,特此鸣谢!
参考文献
[1]崔大方,廖文波,昝启杰,等.广东内伶仃岛国家级自然保护区的植物资源[J].华南农业大学学报,2000,21(3):48–52.
[2]昝启杰,廖文波,陈继敏,等.广东内伶仃岛植物区系的研究[J].西北植物学报,2001,21(3):507–519.
[3]昝启杰,廖文波,蓝崇钰,等.广东内伶仃岛植被分类系统和典型群落的样地标志[J].生态科学,2000,19(1):10–18.
[4]中国科学院中国植物志编辑委员会.中国植物志[M](第60(1)卷).北京:科学出版社,1998:99.
[5]The IUCN Red List of Threatened Species[DB/OL].http://dx.doi.org/10.2305/IUCN.UK.1998.RLTS.T34784A9884 205,2016.3.
[6]陈里娥,杨琼,大可,等.内伶仃岛[M].广州:中山大学出版社,2014:51–52.
[7]覃朝锋,李贞,董汉飞.珠江口内伶仃岛植被[J].生态科学,1990(2):23–34.
[8]ANNE E.MAGURRAN.张峰主译.生物多样性测度[M].北京:科学出版社,2011:48-55,66–79.
[9]王伯荪,余世孝,彭少麟,等.植物群落学实验手册[M].广州:广东高等教育出版社,1996.
[10]APPANAH S,PUTZ FE.Climber abundance in virgin dipterocarp forest and the effect of prefelling climber cutting on logging damage[J].Malaysian Forester,1984,47:335–342.
[11]廖文波,张宏达.广东蕨类植物区系的特点[J].热带亚热带植物学报,1994,2(3):1–11.
[12]吴征镒,周浙昆,李德铢,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245–257.
[13]吴征镒.中国种子植物属的分布区类型专辑[J].云南植物研究,1991(增刊 IV):1–139.
[14]DIETER MUELLER-DOMBOIS,HEINZ ELLENBERG.鲍显诚,等译.植被生态学的目的和方法[M].北京:科学出版社,1986:2–10.
[15]郭泉水,江洪,王兵,等.中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究[J].生态学报,1999,19(4):573–574.
[16]陈红锋,严岳鸿,秦新生,等.海南铜铁岭热带低地雨林群落特征研究[J].西北植物学报,2005,25(1):103–112.
[17]李家湘,熊高明,徐文婷,等.中国亚热带灌丛植物生活型组成及其与水热因子的相关性[J].植物生态学报,2017,41(1):147–156.
[18]ESAU K.Anatomy of seedplants[M].2nd.New York:John Wiley and Sons Press,1977:315–372.
[19]彭少麟,周厚诚,陈天杏,等.广东森林群落的组成结构数量特征[J].植物生态学与地植物学学报,1989,13(1):10–17.
[20]彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1996:30–101.
[21]田广红,丁明艳,杨雄邦,等.珠海市淇澳岛肉实树群落及其物种多样性特征[J].植物科学学报,2013,31(5):461–466.
[22]彭少麟,王伯荪.鼎湖山森林群落分析I.物种多样性[J].生态科学,1983(1):11–17.
[23]胡晓敏,董安强,王发国,等.广东南岭大东山浙江润楠群落物种多样性与区系地理成分分析[J].植物科学学报,2011,29(3):265–271.
[24]张宏达,王伯荪,胡玉佳,等.香港植被[J].中山大学学报:自然科学论丛,1989,8(2):11–40.
[25]梁冠欣,郑希龙,王发国,等.澳门青洲山假柿木姜子群落特征及多样性研究[J].林业资源管理,2009(4): 5 4–60.
[26]汪殿蓓,暨淑仪,陈飞鹏,等.深圳南山区天然森林群落多样性及演替现状[J]生态学报,2003,23(7):1415–1422.
