不同温度下两株球孢白僵菌侵染西花蓟马的生长动力学及其毒力
2018-05-14刘晓晨吴圣勇雷仲仁王海鸿
刘晓晨,吴圣勇,雷仲仁,王海鸿
不同温度下两株球孢白僵菌侵染西花蓟马的生长动力学及其毒力
刘晓晨,吴圣勇,雷仲仁,王海鸿
(中国农业科学院植物保护研究所植物病虫害生物学国家重点试验室,北京 100193)
【目的】通过比较两株球孢白僵菌()在不同温度条件下侵染西花蓟马()的生长动力学及其毒力,探讨球孢白僵菌菌体增殖和杀虫毒力的关系,为提高球孢白僵菌对西花蓟马的杀虫效率提供理论支持。【方法】首先,在20、25和30℃ 3个温度条件下,连续记录第1—8天西花蓟马被两株球孢白僵菌菌株(SCWJ-2和GZGY-1-3)侵染后的死亡率,并以未被真菌感染的西花蓟马为对照,计算其累积校正死亡率,选择第3天的数据(对照死亡率为2%—5%)比较两个菌株的杀虫效率。其次,在20、25和30℃ 3个温度条件下,连续记录第1—8天内平板培养的上述两个菌株的菌落直径,选取第3天两个菌株的菌落生长直径数据进行比较分析。最后,提取20、25和30℃温度下,西花蓟马分别被两株球孢白僵菌侵染第1、2和3天的混合DNA,通过实时荧光定量PCR(qRT-PCR)技术定量球孢白僵菌在每个宿主样品体内的拷贝数,选取第3天两个菌株的基因拷贝数进行比较分析。【结果】生物测定结果显示,在检测温度(20—30℃)范围内,球孢白僵菌菌株GZGY-1-3和SCWJ-2对西花蓟马成虫均具有较高致病性,无论任何温度和何种菌株,西花蓟马从处理后第2天开始有死亡个体出现,第8天时,SCWJ-2和GZGY-1-3造成的校正死亡率分别为83%—91%和79%—90%。以第3天的校正死亡率为指标(对照死亡率为2%—5%),30℃下菌株SCWJ-2毒力显著高于GZGY-1-3(<0.05),25℃和20℃下两个菌株毒力无显著差异(>0.05)。平板培养试验表明,在检测温度(20—30℃)范围内,两个菌株菌落直径随着时间推移而增加,第8天时,菌株SCWJ-2和GZGY-1-3的菌落直径分别为31—36、28—32 mm。选取第3天菌落生长直径数据进行比较,菌株SCWJ-2在3个温度下的菌落直径均显著大于菌株GZGY-1-3(<0.05)。实时荧光定量结果显示,除30℃下的菌株SCWJ-2之外,两个菌株在西花蓟马体内的基因拷贝数都在第1天时下降,2 d后又逐渐上升。使用第3天西花蓟马体内的基因拷贝数进行分析,在30℃下菌株SCWJ-2真菌拷贝数显著大于GZGY-1-3(<0.05),25℃和20℃下两个菌株基因拷贝数无显著差异(>0.05)。【结论】真菌在被侵染虫体内的基因拷贝数受到菌株和温度的影响,这与生物测定结果相一致。与菌株GZGY-1-3相比,SCWJ-2更适合高温条件下对西花蓟马的防治。
球孢白僵菌;西花蓟马;真菌生长动力学;实时荧光定量PCR
0 引言
【研究意义】西花蓟马()是一种重要的世界性农业害虫[1-3],对农业生产构成较大威胁[4-6]。球孢白僵菌()是一种广谱性虫生真菌,已被大量应用于西花蓟马的防治[7-12]。虫生真菌孢子附着并穿透昆虫体壁[13]后,菌丝在昆虫血腔内生长,能够产生有毒代谢产物[14]、消耗宿主体内营养、穿透宿主的重要器官和阻止血淋巴流动等,最终导致宿主昆虫死亡[15]。然而,室内适温下筛选出的对西花蓟马具有相似杀虫效率的球孢白僵菌菌株,在田间实际应用时效率却显著不同[16-17]。菌体增殖能力的差异是引起菌株间毒力差异的重要原因[18],田间温度可能通过影响真菌菌体增殖,导致不同菌株间最终杀虫效率不同[19]。因此,研究不同温度对球孢白僵菌侵染的生长动力学影响,对提高田间防治效果具有重要意义。【前人研究进展】目前,许多评价真菌菌体增殖与杀虫效率关系的研究大多是通过体外试验完成的。例如,检测液体培养的菌丝和孢子数,固体培养的菌落直径等[20-25]。EKESI等[20]报道了温度对2株球孢白僵菌和4株绿僵菌()的孢子萌发和菌落直径生长的影响,进而影响了对蓟马()的致病性;Adamo等[24]研究表明,随着温度的升高灵杆菌()或蜡状芽孢杆菌()生长速率加快,进而影响对寄主蟋蟀()的致病能力;HUNT等[25]检测了不同温度下昆虫体内病原菌的生长状况,把感染绿僵菌()的果蝇()分别于不同温度环境下处理相同时间后采样,培养并计数菌落的数量,结果表明昆虫体表病原菌生长状况与温度密切相关。然而,上述方法都无法模拟真菌当时所处的寄主体内营养和免疫压力环境,不能很好地代表真菌的自然生长动力学。关于检测真菌在活体内增殖的方法,曾有报道使用显微镜法测定血球计数板上被感染的昆虫体内芽生孢子和/或菌丝片段的数量[26],但这一技术在感染初期和末期得到的结果很不精确,因为宿主在感染初期其血淋巴中的菌体很少,而在感染后期芽生孢子和菌丝片段又太多,均无法精确计数[26]。【本研究切入点】实时荧光定量PCR(qRT-PCR)技术为准确定量真菌在昆虫体内生长数量提供了便利[27-29]。但目前为止,还未有利用此项技术分析同种真菌不同菌株在不同温度下昆虫宿主体内生长动力学的研究。【拟解决的关键问题】针对适温下对西花蓟马杀虫毒力相似,而高温下显著不同的球孢白僵菌两个菌株GZGY-1-3和SCWJ-2,在设定的温度下,比较两个菌株经平板培养的菌落直径和侵染蓟马后在虫体内的基因拷贝数,结合生物测定数据,分析真菌菌体增殖和杀虫效率间的关系,以期为提高田间应用球孢白僵菌防治西花蓟马的效率提供理论依据。
1 材料与方法
试验于2016—2017年在中国农业科学院植物保护研究所完成。
1.1 供试昆虫
西花蓟马成虫于2013年采自北京市昌平区的辣椒上,在室内用豆角饲养,置于人工气候箱内(MLR-351H,SANYO Electric Co., Ltd)。饲养温度(25±1)℃,光周期14L﹕10D,相对湿度60%—80%。
1.2 供试菌株
球孢白僵菌菌株SCWJ-2和GZGY-1-3分别采集于四川和贵州,现保藏于中国典型培养物保藏中心,保藏号分别为CGMCC No.9253和No.9254。试验时,用无菌接种环于斜面上刮下分生孢子,涂于CMA产孢培养基(琼脂20 g·L-1、硝酸铵1 g·L-1、玉米粉20 g·L-1、蛋白胨5 g·L-1、麦麸10 g·L-1、水合硫酸镁1 g·L-1和硝酸二氢钾3 g·L-1)上,在恒温培养箱(GXZ-9240A)光周期12 L﹕12D,(25±1)℃中培养15 d。
孢子萌发率检测:用接种环刮下2 mg孢子粉置于50 mL的三角瓶中,加入10 mL高温灭菌的萌发液(4%葡萄糖,1%酵母提取物,0.05吐温-80),在(25±1)℃、180 r/min摇床上培养18 h,于400×显微镜(OLYMPUS,BX51)下镜检,萌发率>90%可用于试验。
孢子悬浮液制备:将孢子加入灭菌的0.05%吐温-80溶液,配成107个孢子/mL的悬浮液。
1.3 生物测定
取约2 000头西花蓟马成虫置于上述孢子悬浮液中浸泡5 s,对照组蓟马用0.05%的吐温溶液处理5 s。用细毛笔将西花蓟马挑到滤纸上,吸干多余液体,然后转移至放有豆角叶片的培养皿(直径约为7 cm)中,用保鲜膜密封培养皿,并用细针头在保鲜膜上扎孔保持透气。分别放入20、25和30℃的培养箱内,光周期12 L﹕12D,每天记录死亡的西花蓟马数量,并把死亡蓟马挑入一个垫有湿润滤纸的灭菌的培养皿内,5—6 d后显微镜观察是否有菌丝长出,以确定是否为白僵菌侵染致死。连续记录8 d,计算其累积校正死亡率并制作西花蓟马的累积校正死亡率曲线。每个处理重复6次,每次100头西花蓟马。在3个温度下,选择第3天的数据(对照死亡率为2%—5%)比较两个菌株杀虫效率。
1.4 菌落直径测定
参考TEFERA等[23]的方法,将15 mL的萨氏培养基倒入直径9 cm的灭菌培养皿冷却,取50 μl 107个/mL的球孢白僵菌孢子悬浮液滴于培养基中央,然后分别置于20、25和30℃温度环境,光周期14L﹕10D的培养箱内培养。每天定时测量菌落直径并进行比较,每个温度下每个菌株重复6次。
1.5 西花蓟马体内球孢白僵菌生长动力学
取样:用上述球孢白僵菌孢子悬浮液处理西花蓟马后,分别放置于20、25和30℃下,于0、24、48和72 h取样,每个时间点取6次重复,每个重复100只。提取DNA进行后续绝对荧光定量试验。
DNA提取:研磨方法参考Bell等[27],在装有样品的研磨管中分别加入0.25 g直径为0.2 mm的锆珠和0.8 mm的硅珠,然后放在TissueLyzerTM(Qiagen)组织研磨仪中30 Hz下干磨1 min。研磨期间,为了保持每个管中真菌都能够被充分研磨,每隔15 s中断一次,进行研磨管位置的重新调整。然后加入200μl核裂解液,继续如上研磨。研磨结束,采用Wizard®基因组DNA纯化试剂盒(Promega,USA)提取组织体内的混合DNA,用Nuclease-Free Water溶解。
标准曲线和样品定量:通过球孢白僵菌的转录间隔区(ITS2)与核糖体RNA序列(GenBank登录号:AF345539)设计qRT-PCR中的引物和探针。探针:6-FAM-ACAGCTCGCACCGGA-MGB;上游引物:5′-GCCGGCCCTGAAATGG-3′;下游引物:5′-GATTCGAGGTCAACGTTCAGAAG-3′。qPCR试验在Applied Biosystems 7500 Real-Time PCR System 上运行,运行体系为20 μl:8 μlNuclease-Free Water,10 μl TaqMan Universal Master Mix Ⅱ,无UNG,上下游引物各0.5μl,1 μl探针。运行过程:95℃预变性2 min,95℃15 s,60℃1 min循环40次。标准曲线制作:用无菌的0.05%吐温-80配置108个/mL球孢白僵菌孢子悬浮液,取1 mL至研磨管离心甩去上清,然后按上述方法研磨提取DNA。将提取的108个GZGY-1-3孢子的DNA按10倍梯度从108逐级稀释至102(考虑到试验时DNA的剂量,100个孢子的DNA量被认为是仪器检测的极限),利用qPCR制得。通过标准曲线来对样品进行绝对定量。
通过对标准品进行10倍梯度稀释后测定了7个点(108—102)的扩增曲线,得出其标准曲线:=-3.353+35.864(2=0.998)。与生物测定和菌落直径试验相一致,选取被侵染第3天的蓟马体内基因拷贝数,进行同一温度下两个菌株间数据比较。
1.6 数据处理与分析
用Abbott公式计算累积校正死亡率。用SAS 9.2软件进行差异显著性分析。相同温度下,第3天两个菌株间杀虫毒力差异性、平板培养菌落直径差异性以及被侵染的西花蓟马体内菌体基因拷贝数的差异性均使用检验,显著性水平为<0.05。基因拷贝数在数据分析前进行对数转换。
2 结果
2.1 球孢白僵菌GZGY-1-3和SCWJ-2菌株杀虫毒力比较
生物测定结果显示,在检测温度(20—30℃)范围内,球孢白僵菌菌株GZGY-1-3和SCWJ-2对西花蓟马成虫均具有较高致死率。无论任何温度和何种菌株,西花蓟马从处理后第2天开始死亡,第8天时,SCWJ-2和GZGY-1-3造成的校正死亡率分别为83%—91%和79%—90%(图1)。以第3天的校正死亡率为指标(对照死亡率为2%—5%),在适温(20和25℃)下,两个菌株杀虫毒力间没有显著差异,较高温度(30℃)下菌株SCWJ-2毒力显著高于GZGY-1-3(<0.05)(图2)。
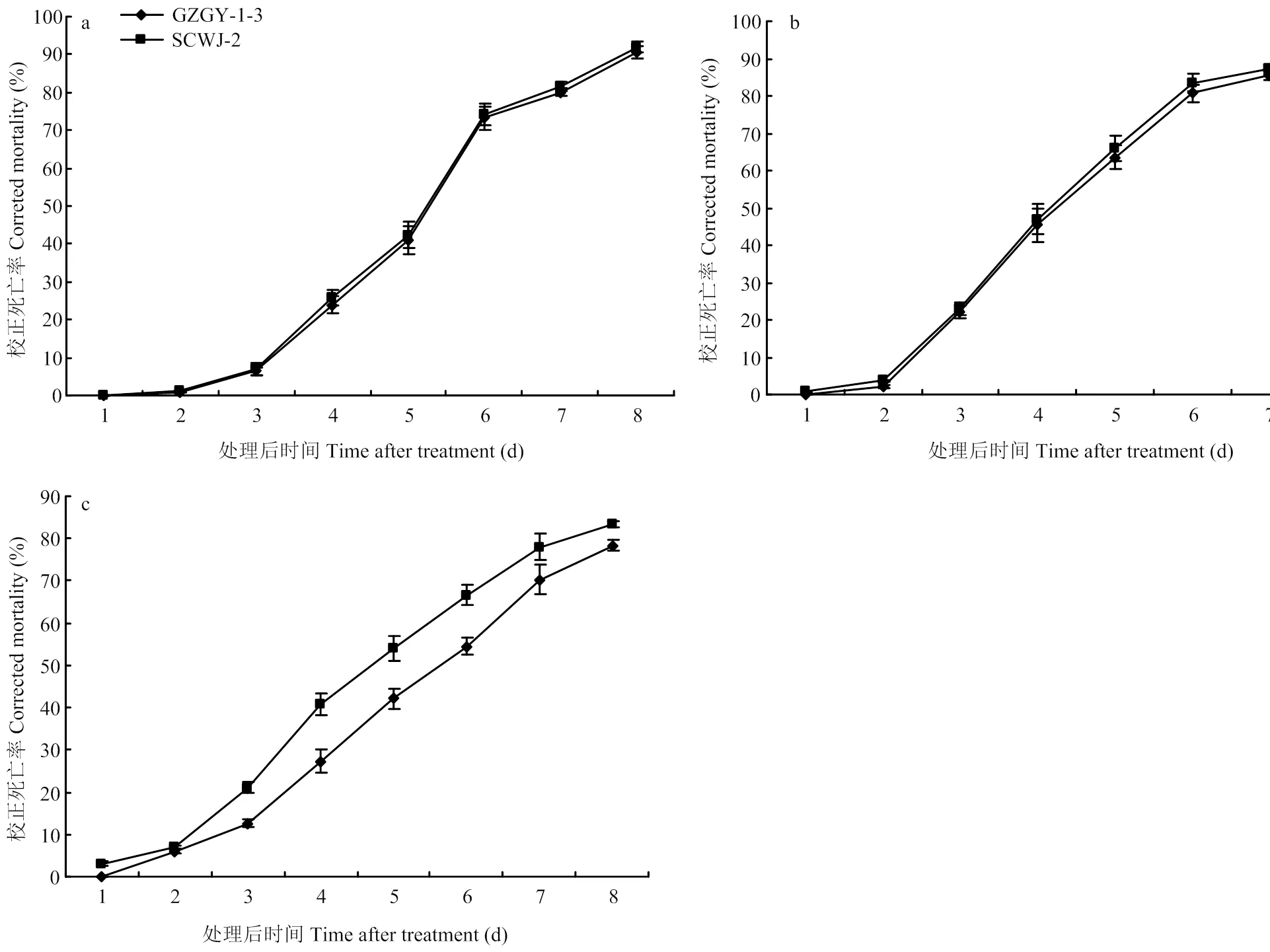
a:20℃;b:25℃;c:30℃
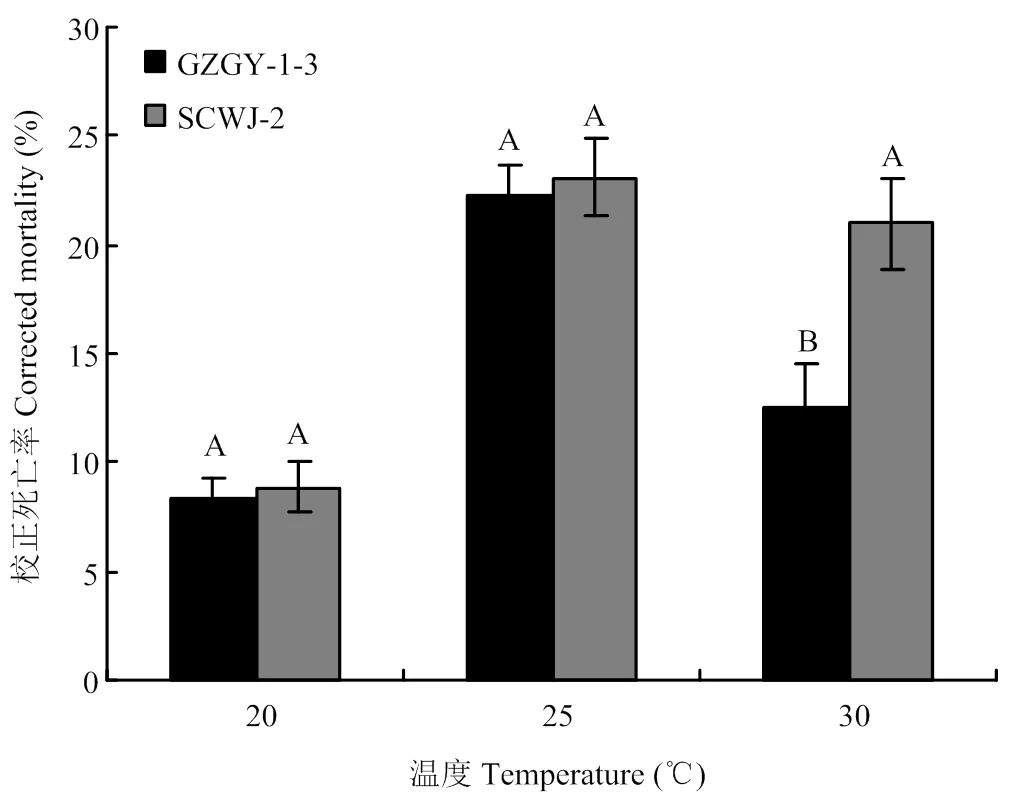
同一温度下柱上不同字母表示菌株间差异显著(P<0.05)。下同
2.2 球孢白僵菌GZGY-1-3和SCWJ-2菌株菌落直径比较
平板培养结果显示,在检测温度(20—30℃)范围内,两个菌株菌落直径随着时间推移而增加,第8天时,菌株SCWJ-2和GZGY-1-3的菌落直径分别为31—36、28—32 mm(图3)。对应生物测定结果,选取第3天菌落生长直径数据,在20、25和30℃下,菌株SCWJ-2菌落直径均显著大于菌株GZGY-1-3(<0.05)(图4)。
2.3 球孢白僵菌GZGY-1-3和SCWJ-2菌株在被侵染的蓟马体内基因拷贝数
如图5所示,除30℃下的菌株SCWJ-2之外,两个菌株在西花蓟马体内的基因拷贝数都在第1天时下降,2 d后又逐渐上升。使用第3天西花蓟马体内的基因拷贝数进行分析,结果如图6所示,在同一温度(20或25℃)下,菌株SCWJ-2和菌株GZGY-1-3在西花蓟马体内的基因拷贝数无显著差异,而在30℃下,菌株SCWJ-2的基因拷贝数显著高于菌株GZGY-1-3(<0.05)。
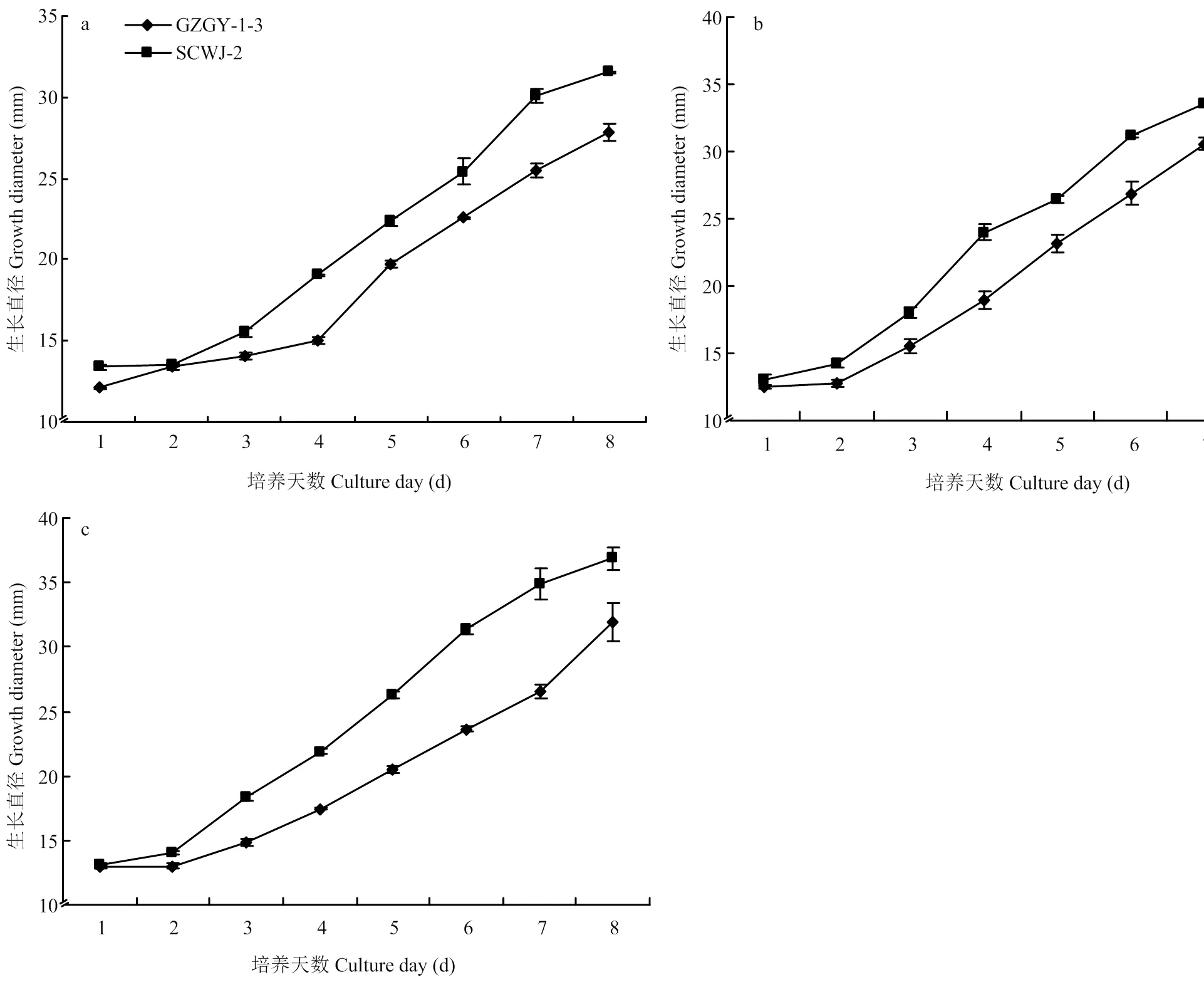
a:20℃;b:25℃;c:30℃
图3 不同温度下两株球孢白僵菌菌株的菌落直径日生长
Fig. 3 The daily diameter growth of the two strains ofunder different temperatures

图4 不同温度下球孢白僵菌菌株GZGY-1-3和SCWJ-2菌落直径
3 讨论
虫生真菌作为防治害虫的重要生防因子,其杀虫效率受实际应用时的环境温度的影响。环境温度通过影响菌体增殖进而影响虫生真菌的杀虫毒力。以前的研究方法中,以显微镜法[26]为代表的活体检测无法进行准确计数,而离体培养的方式[20-23]不能很好的代表被侵染的寄主体内的真菌所处的营养或免疫条件[29]。因此,本研究通过PCR方法检测真菌在寄主昆虫体内的基因拷贝数,以检测离体平板菌落直径的方法为对照,分析其与生物测定试验结果的关系。结果表明,在3个温度下,球孢白僵菌菌株GZGY-1-3和SCWJ-2在虫体内的基因拷贝数差异,而不是平板菌落直径差异,与生物测定毒力的差异相一致。在20℃或25℃下两个菌株的杀虫效率相似,在30℃下菌株SCWJ-2杀虫效率显著高于GZGY-1-3,更适于高温环境对西花蓟马的防治。

a:20℃;b:25℃;c:30℃
菌株SCWJ-2在30℃相对高温下,杀虫效果显著高于GZGY-1-3,可能由于其更好的高温耐受力。考虑到SCWJ-2和GZGY-1-3分别来自四川和贵州,这两个地区的温度环境有一定的差异,菌株来源地不同可能造成两者在相对高温下耐热力不同,进而造成杀虫毒力的不同。这与俞佳等的结论基本相似[30-31]。
袁盛勇等[11,32]研究表明,球孢白僵菌菌株MZ041016和MZ060812在孢子浓度分别为3.6×108和3×108个/mL时,对西花蓟马成虫的致死率为分别为82.31%和88.42%;王雅卉等[33]研究表明,菌株CYT4侵染西花蓟马第6天后累积校正死亡率为49.78%。本研究生物测定结果显示,25℃下两个菌株在孢子浓度107个/mL时,8 d后对西花蓟马的致死率分别为89%(菌株SCWJ-2)和86%(菌株GZGY-1-3),达到了对西花蓟马较好的毒力效果。综合考虑对高温的耐受能力,菌株SCWJ-2和GZGY-1-3是有潜力用于防治西花蓟马的高效菌株[12]。

图6 不同温度下球孢白僵菌菌株GZGY-1-3和SCWJ-2侵染西花蓟马3 d后体内基因拷贝数对数
白僵菌侵染西花蓟马的实时荧光定量结果表明,与处理 0 h 相比,菌株SCWJ-2和GZGY-1-3的基因拷贝数大部分在处理24 h时下降,而处理48和72 h时开始回升。侵染初期孢子拷贝数下降可能与以下因素有关:(1)蓟马的活动将体表部分孢子抖落;(2)孢子穿透昆虫表皮所造成的损耗。Anderson等[28]在研究球孢白僵菌和绿僵菌感染家蝇()时也发现了相似的生长模式。而30℃下菌株SCWJ-2拷贝数在24 h时并未下降反而上升,这可能是由于30℃为菌株SCWJ-2的适宜温度,宿主体内的真菌增殖弥补了体表真菌的损失和消耗。
30℃下,菌株SCWJ-2在蓟马体内的拷贝数显著高于GZGY-1-3,这与前者比后者具有更高的杀虫毒力这一生物测定结果相一致。在20或25℃下,两个菌株在虫体内拷贝数和生物测定毒力方面结果一致,均无显著差异,这说明在一定程度上,菌株在宿主昆虫体内增殖的差异性决定了菌株的毒力不同。此外,真菌在昆虫体内增殖时,还会产生毒素等[34],这是否也会导致不同温度下不同菌株毒力的差异,还需进一步研究。
菌落直径(真菌在寄主体外生长)检测结果显示,在3个检测温度下,菌株SCWJ-2菌落直径均大于GZGY-1-3,20℃和25℃下两个菌株的菌落直径差异性与生物测定结果不一致,只有在30℃下与生物测定结果相一致。可见,检测离体情况下平板培养的真菌菌落直径这一方法有一定的局限性。真菌在入侵到昆虫体内时,会接触到昆虫血腔中特有的营养环境[35]和免疫响应系统[25,36],这是与离体培养基所提供的环境截然不同的。此外,温度对平板培养的真菌菌落直径的影响通常选取的是第7—10天的数据[19,37-38],远远超过了真菌在昆虫体内的增殖时期。因此,真菌在活体昆虫内的增殖很难用体外平板上菌落直径这一数据来准确描述。与之相比,不同温度下宿主昆虫体内真菌的基因拷贝数更好地解释了耐热性不同菌株杀虫效率的差异。
4 结论
在适温下,球孢白僵菌菌株GZGY-1-3和SCWJ-2均对西花蓟马有较好的杀虫效率。在较高的温度下,菌株SCWJ-2比GZGY-1-3在被侵染的蓟马体内基因拷贝数更大,引起的校正死亡率更高,更适合高温季节对西花蓟马进行生物防治。真菌在虫体内的基因拷贝数很好地解释了不同菌株和不同温度条件引起的杀虫毒力的差异。结果为球孢白僵菌在高温季节防治西花蓟马的可行性提供了理论支持。
[1] REITZ S R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest., 2009, 92(1): 7-13.
[2] REITZ S R, Gao Y L, LEI Z R. Thrips: pests of concern to China and the United States., 2011, 10(6): 867-892.
[3] 雷仲仁, 问锦曾, 王音. 危险性外来入侵害虫——西花蓟马的鉴别、危害及防治. 植物保护, 2004, 30(3): 63-66.
LEI Z R, WEN J Z, WANG Y. The identification, damage and control of an invasive western flower thrips ()., 2004, 30(3): 63-66. (in Chinese)
[4] Ullman D E, Meideros R, Campbell L R, Whitfield A E, Sherwood J L, German T L. Thrips as vectors of tospoviruses., 2002, 36: 113-140.
[5] 张友军, 吴青君, 徐宝云, 朱国仁. 危险性外来入侵生物——西花蓟马在北京发生危害. 植物保护, 2003, 29(4): 58-59.
ZHANG Y J, WU Q J, XU B Y, ZHU G R. Dangerous invasive alien species - Western flower thrips in Beijing., 2003, 29(4): 58-59. (in Chinese)
[6] 王海鸿, 雷仲仁, 李雪, 代安国, 陈翰秋. 西藏发现重要外来入侵害虫——西花蓟马. 植物保护, 2013, 39(1): 181-183.
WANG H H, LEI Z R, LI X, DAI A G, CHEN H Q. An important invasive pest,, inspected in Tibet., 2013, 39(1): 181-183. (in Chinese)
[7] 张慧, 吴圣勇, 李娟, 张璐璐, 张林雅, 雷仲仁. 不同培养基继代培养球孢白僵菌对西花蓟马毒力和产孢量的影响. 中国农业科学, 2016, 49(15): 2977-2987.
ZHANG H, WU S Y, LI J, ZHANG L L, ZHANG L Y, LEI Z R. Influence of subculture on virulence toand conidial production of the entomopathogenic fungus., 2016, 49(15): 2977-2987. (in Chinese)
[8] LEE S J, KIM S, KIM J C, LEE M R, HOSSAIN M S, SHIN T S, KIM T H, KIM J S. Entomopathogenicgranules to control soil-dwelling stage of western flower thrips,(Thysanoptera: Thripidae)., 2017, 62(5): 639-648.
[9] WU S Y, GAO Y L, ZHANG Y P, WANG E D, XU X N, LEI Z R. An entomopathogenic strain ofagainstwith no detrimental effect on the predatory mite: Evidence from laboratory bioassay and scanning electron microscopic observation., 2014, 9(1): e84732.
[10] 李银平, 雷仲仁, 王海鸿. 对西花蓟马高效的球孢白僵菌菌株筛选及产孢特性研究. 中国生物防治学报, 2013, 29(2): 219-226.
LI Y P, LEI Z R, WANG H H. Selection ofstrains againstand their conidial production characteristics., 2013, 29(2): 219-226. (in Chinese)
[11] 袁盛勇, 张宏瑞, 孔琼, 李正跃, 骆弟乾, 王学丽, 董丽玲. 球孢白僵菌MZ060812菌株对西花蓟马的致病性研究. 西北农林科技大学(自然科学版), 2010, 38(3): 145-149.
YUAN S Y, ZHANG H R, KONG Q, LI Z Y, LUO D Q, WANG X L, DONG L L. Detection of pathogenicity ofMZ060812 against, 2010, 38(3): 145-149. (in Chinese)
[12] ZHANG T, REITZ S R, WANG H H, LEI Z R. Sublethal effects of(Ascomycota: Hypocreales) on life table parameters of(Thysanoptera: Thripidae)., 2015, 108(3): 975-985.
[13] 张烨, 雷仲仁, 王海鸿, 吉青战. 球孢白僵菌HsbA蛋白的原核表达及免疫定位. 中国农业科学, 2013, 46(21): 4534-4541.
ZHANG Y, LEI Z R, WANG H H, JI Q Z. Prokaryocyte expression and immune localization of HsbA in., 2013, 46(21): 4534-4541. (in Chinese)
[14] HAJEK A E, LEGET R J S. Interactions between fungal pathogens and insect hosts., 1994, 39: 293-322.
[15] Clarkson J M, CHAMLEY A K. New insights into the mechanisms of fungal pathogenesis in insects., 1996, 4(5): 197-203.
[16] 王海鸿, 王登杰, 张桃, 李雪, 雷仲仁. 一种球孢白僵菌菌株GZGY-1-3及其应用: ZL 2014 1 0449467.4[P]. (2017-04-19)[2017- 11-21].
WANG H H, WANG D J, ZHANG T, LI X, LEI Z R. A strain ofGZGY-1-3 and its application: ZL 2014 1 0449467.4[P]. (2017-04-19)[2017-11-21]. (in Chinese)
[17] 王海鸿, 王登杰, 张桃, 雷仲仁. 一种球孢白僵菌菌株SCWJ-2及其应用: ZL 2014 1 0449469.3[P]. (2017-05-24)[2017-11-21].
WANG H H, WANG D J, ZHANG T, LEI Z R. A strain ofSCWJ-2 and its application: ZL 2014 1 0449469.3[P]. (2017-05-24) [2017-11-21]. (in Chinese)
[18] KERSHAW M J, MOORHOUSE E R, BATEMAN R, REYNOLDS S E, CHAMLEY A K. The role of destruxins in the pathogenicity offor three species of insect., 1999, 74(3): 213-223.
[19] MWAMBURI L A, LAING M D, MILLER R M. Effect of surfactants and temperature on germination and vegetative growth of., 2015, 46(1): 67-74.
[20] EKESI S, MANIANIA N K, AMPONG-NYARKO K. Effect of temperature on germination, radial growth and virulence ofandon., 1999, 9(2): 177-185.
[21] Yeo H, Pell J K, Alderson P G, Clark S J, Pye B J. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and on their pathogenicity to two aphid species., 2003, 59(2): 156-165.
[22] SHIMAZU M. Effects of temperature on growth ofF-263, a strain highly virulent to the Japanese pine sawyer,, especially tolerance to high temperatures., 2004, 39(3): 469-475.
[23] TEFERA T, PRINGLE K. Germination, radial growth, and sporulation ofandisolates and their virulence to(Lepidoptera: Pyralidae) at different temperatures., 2003, 13(7): 699-704.
[24] Adamo S A, Lovett M M. Some like it hot: the effects of climate change on reproduction, immune function and disease resistance in the cricket., 2011, 214(12): 1997-2004.
[25] HuntV L,ZhongW,McClureC D,MlynskiD T, DuxburyE M,CharnleyA K, PriestN K. Cold-seeking behaviour mitigates reproductive losses from fungal infection in., 2016, 85(1): 178-186.
[26] OUEDRAOGO R M, CUSSON M, GOETTEL M S, BRODEUR J. Inhibition of fungal growth in thermoregulating locusts,, infected by the fungusvar.., 2003, 82(2): 103-109.
[27] BELL A S, BLANFORD S, JENKINS N, THOMAS M B, READ A F. Real-time quantitative PCR for analysis of candidate fungal biopesticides against malaria: technique validation and first applications., 2009, 100(3): 160-168.
[28] ANDERSON R D, BELL A S, BLANFORD S, PAAIJMANS K P, THOMAS M B. Comparative growth kinetics and virulence of four different isolates of entomopathogenic fungi in the house fly (L)., 2011, 107(3): 179-184.
[29] 王登杰, 王海鸿, 张桃, 雷仲仁. 三株球孢白僵菌侵染烟粉虱的比较生长动力学及其毒力. 菌物学报, 2016, 35(5): 577-585.
WANG D J, WANG H H, ZHANG T, LEI Z R. Comparative growth kinetics and virulence of three isolates ofin the whiteflies., 2016, 35(5): 577-585. (in Chinese)
[30] 俞佳. 球孢白僵菌不同来源菌株分生孢子耐热性及其对桃蚜的毒力评价[D]. 杭州: 浙江大学, 2006.
YU J. Variation in conidial thermotolerance and virulence of originally differentstrains against[D].Hangzhou: zhejiang university, 2006. (in Chinese)
[31] VIDAL C, FARGUES J, Lacey L A. Intraspecific variability of: Effect of temperature on vegetative growth., 1997, 70(1): 8-26.
[32] 袁盛勇, 张宏瑞, 孔琼, 王平, 孙士卿, 李正跃, 肖春. 球孢白僵菌MZ041016菌株对西花蓟马的毒力测定. 华中农业大学学报, 2011, 30(2): 197-199.
YUAN S Y, ZHANG H R, KONG Q, WANG P, SUN S Q, LI Z Y, XIAO C. Detection on the virulence ofMZ041016 against(Pergande)., 2011, 30(2): 197-199. (in Chinese)
[33] 王雅卉, 郑长英, 王俊平. 球孢白僵菌对西花蓟马成虫的毒力及体表侵染的扫描电镜观察. 中国生物防治学报, 2011, 27(3): 324-330.
WANG Y H, ZHENG C Y, WANG J P. Virulence oftoadults and scanning electron microscopic observation on its infection process., 2011, 27(3): 324-330. (in Chinese)
[34] 任洁, 韩雪梅, 刘昭, 雷仲仁. 烟粉虱血细胞对白僵菌的防御反应. 中国蔬菜, 2013(12): 61-65.
REN J, HAN X M, LIU Z, LEI Z R. Defense response of hemolymph into, 2013(12): 61-65. (in Chinese)
[35] 张慧, 吴圣勇, 王晓青, 雷仲仁. 球孢白僵菌对葱蝇成虫血淋巴蛋白质及游离氨基酸的影响. 中国农业科学, 2017, 50(3): 591-598.
ZHANG H, WU S Y, WANG X Q, LEI Z R. Changes in the contents of proteins and free amino acid in haemolymph ofadult infected by., 2017, 50(3): 591-598. (in Chinese)
[36] Ferguson L V, Heinrichs D E, Sinclair B J. Paradoxical acclimation responses in the thermal performance of insect immunity., 2016, 181(1): 77-85.
[37] 张永军, 王中康, 殷幼平, 裴炎. 球孢白僵菌的生物学特性及对小麦蚜虫的毒力. 西南农业大学学报, 2001, 23(2): 144-146.
ZHANG Y J, WANG Z K, YIN Y P, PEI Y. Biological characteristics of three isolates ofand their virulence against wheat aphids., 2001, 23(2): 144-146. (in Chinese)
[38] 魏灵燕, 宋健, 曹伟平, 冯书亮, 王金耀, 朱宝成, 杜立新. 球孢白僵菌不同分离株的生物学特性的研究. 中国生物防治学报, 2012, 28(4): 502-507.
WEI L Y, SONG J, CAO W P, FENG S L, WANG J Y, ZHU B C, DU L X. Study on biological characteristics of differentstrains., 2012, 28(4): 502-507. (in Chinese)
growth kinetics and virulence of TwoStrains inunder different temperatures
LIU Xiaochen, WU Shengyong, LEI Zhongren, WANG Haihong
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193)
【Objective】The objective of this study is to compare the growth kinetics and virulence of two strains ofinfected Western flower thrips () under different temperatures, analyze the relationship between the proliferation and insecticidal virulence of, and to provide a theoretical support for improving the efficiency ofagainst.【Method】Firstly, from 1st to 8th day, the mortality of the infectedwith two strains of(SCWJ-2 and GZGY-1-3) under 20, 25, and 30℃ was recorded, and cumulative corrected mortality was calculated while uninfectedwas used as control. The mortality ofinfected with the two strains at the 3rd day (the mortality of control was 2%-5%) was compared. Secondly, under 20, 25 and 30℃, the colony diameter of the two strains was recorded continuously from 1st to 8th day. The colony diameter of the two strains at the 3rd day was compared. Finally, the DNA mixture ofinfected with fungi from 1st to 3rd day under 20, 25 and 30℃ was extracted, respectively. The real-time fluorescence quantitative PCR (qRT-PCR) was used to quantify the copy number of fungi within each insect host sample. The gene copy of two strains under the same temperature at the 3rd day was compared. 【Result】The bioassay results ofagainstshowed that within the tested temperature range (20-30℃), both strains GZGY-1-3 and SCWJ-2 were highly lethal toadults. No matter what the temperature or the strain, the dead individualsappeared from the 2nd day after treatment. On the 8th day, the corrected mortality ofinfected by strain SCWJ-2 and GZGY- 1-3 was 83%-91% and 79%-90%, respectively. The corrected mortality ofinfected by the two strains on the 3rd day was compared (the morality of control were 2%-5%). the virulence of strain SCWJ-2 was significantly higher than that of GZGY-1-3 under 30℃(<0.05), but there was no significant difference between them under 25 and 20℃(>0.05). Within the tested temperature range from 20 to 30℃, the colony diameter of two strains increased with time. On the 8th day, the colony diameter of the strains SCWJ-2 and GZGY-1-3 was 31-36 and 28-32 mm, respectively. On the 3rd day, the colony diameter of strain SCWJ-2 was significantly larger than that of strain GZGY-1-3 under all test temperatures(<0.05). The results of qRT-PCR showed that, with the exception of strain SCWJ-2 at 30℃, the gene copy number of both strains indecreased on the 1st day and gradually increased after 2 days at all the three temperatures. The gene copy number of strain SCWJ-2 withinwas significantly higher than that of strain GZGY-1-3 under 30℃(<0.05), but there was no significant difference between them under 25 and 20℃(>0.05). 【Conclusion】 The number of fungal gene copies within insect host was affected by strains and temperatures, which is in accordance with the result of bioassay. Compared withstrain GZGY-1-3, strain SCWJ-2 is more suitable for controllingunder high temperatures.
;; fungal growth kinetics; real-time fluorescence quantitative PCR (qRT-PCR)
(责任编辑 岳梅)
10.3864/j.issn.0578-1752.2018.08.006
2017-10-30;
2017-11-21
国家重点研发计划(2017YFD0201205)、国家现代农业产业技术体系(CARS-25-B-07)
刘晓晨,E-mail:1804486941@qq.com。
雷仲仁,Tel:010-62815930;E-mail:leizhr@sina.com。通信作者王海鸿,Tel:010-62815930;E-mail:wanghaihong2020@sina.com
