太平洋褶柔鱼秋生群产卵场环境变化及对资源丰度的影响
2018-05-09武胜男陈新军
武胜男,余 为,陈新军
(1.上海海洋大学海洋科学学院,上海 201306;2.海洋渔业科学与食物产出过程功能实验室,青岛国家海洋科学技术实验室,青岛 266237;3.大洋渔业资源可持续开发教育部重点实验室,上海 201306;4.国家远洋渔业工程技术研究中心,上海 201306;5.农业部大洋渔业开发重点实验室,上海 201306)
近几十年来,传统海洋经济鱼类资源逐渐衰退,头足类渔业在世界渔业发展中开始占据重要地位。太平洋褶柔鱼(Todarodes pacificus)为大洋性经济头足类,具有1年生命周期[1],主要分布在西北太平洋21°~50°N海域,主要作业渔场分布在日本海、日本太平洋沿岸以及我国东、黄海[2]。太平洋褶柔鱼具有春、夏、秋、冬繁殖群体,各群体在生长、繁殖等方面存在显著差异[3-4]。目前各国商业渔船主要捕捞秋生群和冬生群,其中,秋生群体在日本海的渔获量占总量的70%左右[5]。秋生群体主要在9-11月进行产卵活动,产卵场从东海北部延伸到九州西岸以及日本海西部沿岸,翌年5-6月在对马暖流及北方冷水边界形成的极锋附近密集集结,7月资源量达到顶峰[6]。已有研究表明,太平洋褶柔鱼的资源丰度、渔场分布与海流、海表层温度(SST)、海表层盐度(SSS)、叶绿素 a(Chl-a)浓度[4,7-8]等因素变化有关,同时还受到厄尔尼诺(El Niño)和拉尼娜(La Niña)现象等气候变化事件的影响[9]。唐峰华等[9]认为,太平洋褶柔鱼渔场一般分布在冷暖流交汇区暖水一侧,冷暖流交汇形成的流隔有利于形成渔场。日本学者志村健等[6]研究表明,太平洋褶柔鱼卵团适宜的孵化水温为 15~23℃。SAKURAI等[10]利用日本1975-1996年渔业捕捞数据和水温数据,对太平洋褶柔鱼产卵场分布进行分析,认为从20世纪80年代末开始,对马海峡和附近的秋生群和冬生群产卵场出现了重叠。桜井泰憲等[11]认为,大气温度与风速对太平洋褶柔鱼年渔获量的变化有显著影响。
研究气候变化对海洋渔业资源变动影响具有重要意义[12],当前太平洋褶柔鱼资源受气候变化和海洋环境影响的研究相当缺乏,特别是不同的气候变化对其资源的变动影响。因此本文以单位捕捞努力量渔获量(CPUE)表征太平洋褶柔鱼资源丰度,利用交相关分析法分析厄尔尼诺、太平洋年代际涛动、产卵场SST、Chl-a浓度及最适产卵温度范围占产卵场总温度范围的比例(Ps)与太平洋褶柔鱼秋生群资源丰度的相关性,以此探讨气候和海洋环境变化对太平洋褶柔鱼秋生群资源丰度的潜在影响,为太平洋褶柔鱼资源的合理利用提供科学依据。
1 材料与方法
1.1 材料来源
(1)渔业生产数据来自日本渔业机构提供的2015年度太平洋褶柔鱼秋生群资源评价报告,时间为2002-2011年,数据来源于 http://abchan.fra.go.jp/digests27/index.html。生产捕捞范围主要分布在日本北陆沿岸、对马海峡附近以及东海海域。数据包括年份、渔获量、捕捞努力量等(表1)。
(2)环境数据包括产卵场SST和Chl-a浓度,时间为2002-2011年9-11月。范围覆盖产卵场[13]30.5°~39°N、124.5°~138°E海域(图 1),空间分辨率为0.1°×0.1°。SST和Chl-a浓度数据 均 来 源 于 Ocean-Watch网 站 (http://oceanwatch.pifsc.noaa.gov/las/servlets/dataset)。厄尔尼诺指数(NI)采用Niño 3.4区海表温度距平值(SSTA)表示,数据来自美国NOAA气候预报中心网站(http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml)。太平洋年代际涛动指数(PDOI)来源于美国华盛顿大学数据库(http://research.jisao.washington.edu/pdo/PDO.latest)。
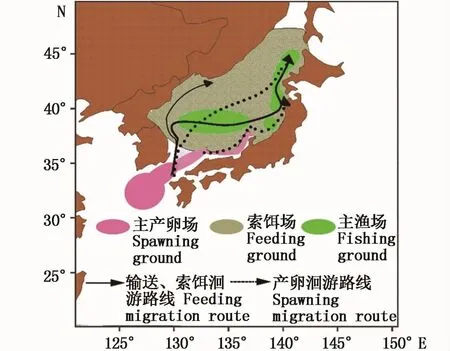
图1 日本太平洋褶柔鱼秋生群分布区域图Fig.1 Distribution of autumn cohort of Todarodes pacificus
1.2 研究方法
(1)前人研究表明,单位捕捞努力量的渔获量(CPUE)可作为太平洋褶柔鱼资源丰度的指标[14]。本文CPUE的计算公式为:

式(1)中,CPUE为各年单位捕捞努力量渔获量,单位为t·d-1;∑Catch为各年所有渔船总渔获量,单位为t;∑Effort为各年总捕捞努力量,单位为d。
(2)不同气候事件的定义。本文采用美国NOAA气候预报中心的标准定义 El Niño与La Niña事 件 (http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml),即 Niño 3.4区 SSTA(下面简称 NI)连续5个月滑动平均值超过+0.5℃,则认为发生1次El Niño事件;若连续5个月低于-0.5℃,则认为发生一次La Niña事件[14]。此外,取各年 PDOI的平均值,若PDOI年平均值为正,则为正位相;若PDOI年平均值为负,则为负位相。本文据此定义2002-2011年发生的不同气候变化。

表1 2002-2011年日本太平洋褶柔鱼渔业统计数据Tab.1 Fishery data of Todarodes pacificus from 2002 to 2011 in Japan
(3)太平洋褶柔鱼秋生群产卵场温度范围为15~23℃,最适产卵温度范围18.9~22.3℃[6]。据此计算2002-2011年9-11月各月最适产卵温度范围占产卵场总温度范围的比例(Ps)。
Ps的计算公式为:

(4)用MATLAB软件利用交相关分析法分别估算NI和PDOI与太平洋褶柔鱼CPUE以及产卵场SST、Chl-a浓度和Ps的相关性。
(5)根据(2)中定义的气候变化,分析不同气候条件下太平洋褶柔鱼秋生群产卵场各环境变量的变化,据此推断气候和海洋环境变化对太平洋褶柔鱼资源丰度可能产生的潜在影响。
2 结果与分析
2.1 CPUE和产量的年间变化
由图2可知,2002-2011年期间,太平洋褶柔鱼秋生群渔获量变化范围为34 746~87 366 t,2002年产量最高,2011年产量最低;CPUE变化范围为2.271~3.162 t·d-1,同样在 2002年最高,而2007年CPUE最低。由此可以看出,太平洋褶柔鱼秋生群产量和CPUE年间波动较大,产量呈逐年下降趋势,2002-2006年产量明显高于2007-2011年,CPUE的波动较产量更为剧烈。
2.2 不同气候变化的定义
依据 El Niño和 La Niña事件的定义[14],2002年1月-2012年12月共发生El Niño事件4次,分别是2002年6月-2003年2月,2004年7月-2005年4月,2006年9月 -2007年1月和2009年7月-2010年4月;发生La Niña事件3次,分别是2007年8月-2008年6月,2010年7月-2011年4月以及2011年8月-2012年3月(图3)。

图2 2002-2011年日本太平洋褶柔鱼各年渔获量及CPUE分布图Fig.2 Annual catch and catch per unit effort(CPUE)of Todarodes pacificus during 2002-2011 in Japan
依据 PDO不同位相的定义[14],2002-2006年位于PDO正位相(PDO暖期),2007-2011年位于PDO负位相(PDO冷期)。
2.3 NI和 PDOI与 CPUE、产卵场 SST、Chl-a浓度及Ps的交相关分析
从图4中可以看出,NI与CPUE呈显著正相关关系,滞后时间为0年时产生最大正影响,对应交相关系数为0.65(P<0.05);NI与产卵场SST呈显著负相关关系,滞后时间为0年时产生最大负影响,对应交相关系数为-0.80(P<0.05);NI和与产卵场Chl-a浓度无显著相关性;NI与产卵场Ps值呈显著正相关关系,在0年时产生最大正影响,对应交相关系数为0.63(P<0.05)。

图3 2002-2011年各月PDOI和Niño 3.4区SSTA时间序列图Fig.3 Time series of monthly Pacific Decadal Oscillation index(PDOI)and Niño 3.4 sea surface temperature(SSTA)during 2002-2011

图4 NI和PDOI分别与太平洋褶柔鱼秋生群CPUE、Chl-a浓度及Ps值的交相关系数Fig.4 Cross correlation coefficients between NI,PDOI and CPUE,Chl-a and Ps
PDOI与CPUE具有显著正相关,滞后时间为-1~0年,在-1年时产生最大正影响,但此时无生物学意义,而对应滞后时间0年时的交相关系数为0.63(P<0.05);PDOI与产卵场SST呈显著负相关性,滞后时间为0年时产生最大负影响,对应交相关系数为 -0.73(P<0.05);PDOI与产卵场Chl-a浓度和Ps值均无显著相关性(图4)。
2.4 不同气候条件下秋生群产卵场环境变量的变化
2.4.1 El Niño和 La Niña年份对应产卵场环境变量变化
根据 El Niño和 La Niña事件的定义,2002年、2004年、2006年以及2009年9-11月发生El Niño事件,定义为 El Niño年;2007年、2010年和2011年9-11月发生 La Niña事件,定义为 La Niña年。据此分析了2002-2011年9-11月发生不同异常环境事件下太平洋褶柔鱼产卵场SST、Chl-a浓度和 Ps值的变化(图 5,表2)。
总体来说,当发生 El Niño事件时,产卵场SST偏低,Chl-a浓度偏高,Ps偏大;当发生 La Niña事件时,产卵场SST偏高,Chl-a浓度偏低,Ps偏小。
2.4.2 PDO正位相和负位相对应产卵场环境变量的变化
根据PDO不同位相的定义,2002-2006年位于PDO正位相(PDO暖期),2007-2011年位于PDO负位相(PDO冷期)。据此分析了位于PDO正位相年份和PDO负位相年份太平洋褶柔鱼产卵场SST、Chl-a浓度和Ps值的变化情况(图6,表3)。
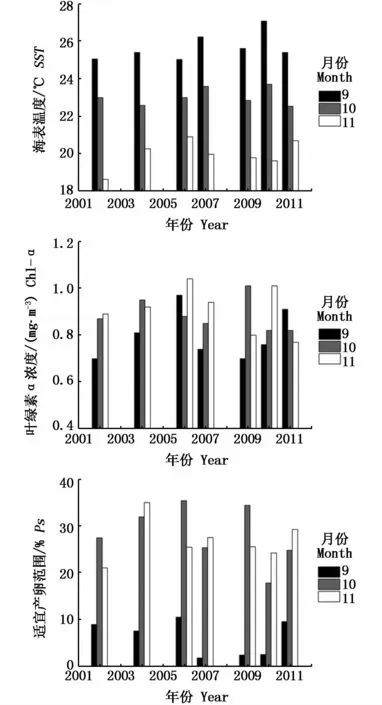
图5 El Niño和 La Niña年份产卵场9-11月SST、Chl-a浓度和Ps值Fig.5 SST,Chl-a and Ps on the spawning ground corresponding to El Niño and La Niña years from September to November

表2 El Niño和La Niña年份产卵场9-11月SST、Chl-a浓度和Ps值的变化Tab.2 Variation of SST,Chl-a and Ps of spawning grounds corresponding to El Niño and La Niña years from September to November
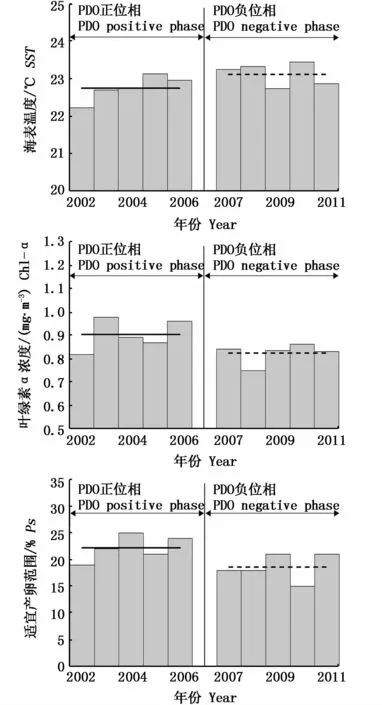
图6 PDO正负位相对应产卵场SST、Chl-a浓度和Ps值Fig.6 SST,Chl-a and Ps corresponding to PDO positive phase and negative phase
以上结果显示,当PDO位于正位相时,产卵场SST偏低,Chl-a浓度偏高,Ps偏大;当PDO位于负位相时,产卵场SST偏高,Chl-a浓度偏低,Ps偏小。
3 讨论
柔鱼科种类为短生命周期大洋性经济种,具有重要的经济和生态价值,但其资源丰度与分布受到气候与海洋环境变化的显著影响[13,15-21]。前人主要通过不同时空尺度气候和海洋环境变化研究头足类资源丰度及分布,描述大尺度异常气候事件对柔鱼科资源渔场和产卵场环境的影响[15-16]。例如徐冰等[15]研究认为,El Niño和 La Niña对茎柔鱼(Dosidicus gigas)产卵场 Ps有显著影响。当发生 La Niña事件时,产卵场 Ps会增大,并且形成广泛的上升流,饵料丰富,这些环境变化有利于茎柔鱼的索铒与繁殖生长,增加茎柔鱼的资源补充量;反之El Niño事件发生时则不利于茎柔鱼资源补充量的增长。本文结果与其恰好相反,当发生El Niño事件时,茎柔鱼产卵场Ps减小,太平洋褶柔鱼秋生群Ps增大;当发生La Niña事件时,反之,即 El Niño事件和 La Niña事件对两种头足类资源丰度的影响是不同的,推测与两种头足类产卵场的最适温度范围有关,茎柔鱼的最适温度范围为24~28℃,明显高于太平洋褶柔鱼秋生群的最适产卵温度范围。唐峰华等[16]认为北太平洋天气变化对柔鱼(Ommastrephes bartramii)捕捞效率和捕捞量有重要影响。他们研究了2009年北太平洋海域形成的台风北移过程,直接导致该海域环境不利于柔鱼生长和生存,北太平洋柔鱼产量剧减。相关文献表明[11],台风对太平洋褶柔鱼的渔获量变化有显著影响,但本文并未涉及,待今后进一步研究。有些学者直接研究柔鱼科鱼类主要作业渔场和产卵场的环境变量如SST、Chl-a浓度、SSS和海流等对该资源丰度的影响[17-18]。例如,宋来军等[17]研究表明,太平洋褶柔鱼渔场各月与各旬的SST均存在差异,7-9月SST呈低、高、低的分布规律,8月中旬达到最高值,7月和9月SST较低。其高产渔场在朝鲜东海岸海域冷暖水团的交汇处,主要分布海域的温度范围为23.5~26.0℃。该结果高于太平洋褶柔鱼秋生群的温度范围,表明同一种头足类在不同海域的温度分布范围有所差异。此外,沈新强等[18]通过北太平洋柔鱼渔场调查资料,认为渔场 Chl-a浓度变化范围为0.03~0.32 mg·m-3,平均值为 0.13 mg·m-3。Chl-a浓度高值海域与高温海域相对应,冷涡区含量较低,暖涡区含量较高。Chl-a浓度与浮游动植物存在正相关关系,对柔鱼中心渔场的资源补充量可能产生间接影响。
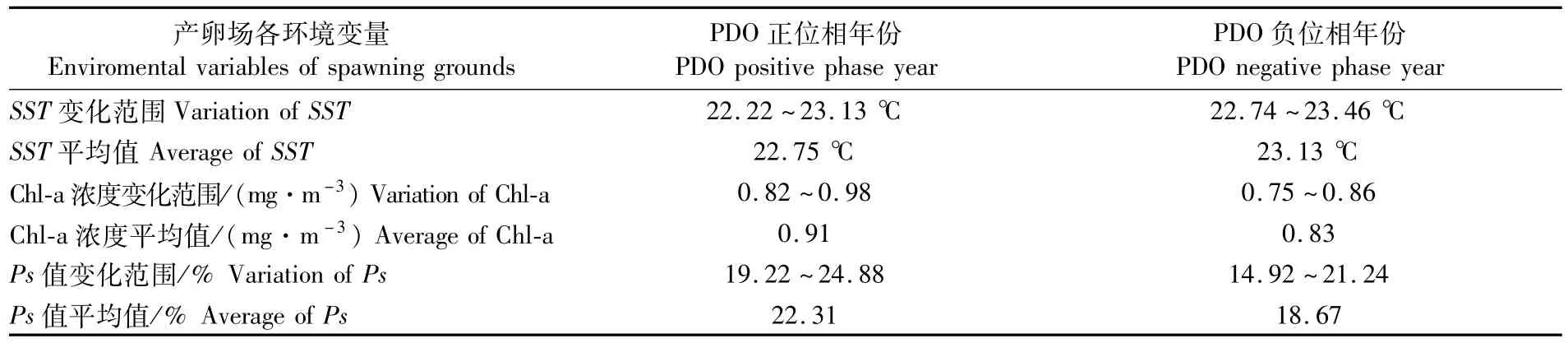
表3 PDO正位相对应产卵场SST、Chl-a浓度和Ps值的变化Tab.3 Variation of SST,Chl-a and Ps corresponding to PDO positive phase and negative phase
已有学者研究了海洋环境因子对太平洋褶柔鱼资源丰度及其渔场分布的影响[22-23]。根据日本学者SHIMORA等[6]等研究,太平洋褶柔鱼是广分布的暖温性鱼类,不同海区以及不同生长阶段,其分布的水温发生相应变化。唐峰华等[9]认为,2010-2013年对马暖流和朝鲜暖流较强,2012-2013年里曼寒流较强,有利于太平洋褶柔鱼渔场的形成。李建生等[4]研究认为,东海太平洋褶柔鱼渔场SSS范围为27.72~34.72 psu,但更多鱼群集中在32.5~34.7 psu之间。
3.1 El Niño和La Niña事件对太平洋褶柔鱼秋生群的影响
研究气候与海洋环境变化对头足类资源的影响对其资源的合理利用有重要意义,从1999年起,关于全球性气候变化的海洋生物资源变动的研究急剧增加,其中,最受关注的是大时间尺度条件下海水的温度变化和转变机制以及长期的海洋气候变动[13,19]。本研究发现,El Niño和 La Niña事件对太平洋褶柔鱼秋生群资源量和产卵场环境产生一定的影响。一般来说,El Niño事件发生时,SST持续变暖,高于La Niña事件发生时的 SST[11],而本研究中 El Niño年份产卵场的 SST却明显低于拉尼娜年份,推测极端气候事件对不同海域不同时期的不同鱼种产生的影响存在差异,其影响机制需要进一步研究。此外,可能由于在La Niña事件发生期间,该鱼种在过暖的水域中延迟成熟,相应地CPUE减少。本研究NI与太平洋褶柔鱼产卵场Chl-a浓度无显著相关性(图4),但Chl-a浓度在不同环境条件下存在明显变化,说明Chl-a浓度在一定程度上影响了太平洋褶柔鱼秋生群的资源丰度,但不是关键因子,推测Chl-a浓度与其他因子共同作用于产卵场,进而对资源丰度产生相应的影响。此外,本研究El Niño年份对应的太平洋褶柔鱼秋生群产卵场Ps总体偏高,高于 La Niña年份(图 5),与CPUE相对应。
结合交相关分析结果,本文研究表明,当发生El Niño事件时,产卵场 SST降低,Ps增加,导致CPUE上升,即太平洋褶柔鱼秋生群资源丰度较高;相反,当发生La Niña事件时,产卵场SST较高,Ps减少,导致CPUE减小,即不利于太平洋褶柔鱼秋生群资源量的补充,进而影响其资源丰度。虽然NI与太平洋褶柔鱼秋生群产卵场Chl-a浓度无显著相关性,但总体来说El Niño年份产卵场Chl-a浓度要高于La Niña年份,同样有利于产生有利的资源补充条件。
3.2 PDO变化对太平洋褶柔鱼秋生群的影响
研究发现,PDO变化在一定程度上影响太平洋褶柔鱼秋生群资源量和产卵场环境。PDO位于正位相年份时,太平洋褶柔鱼秋生群产卵场SST明显低于PDO负位相年份。当PDO位于正位相(暖期)时,西太平洋海域海水变冷,部分东太平洋海域海水变暖[24],这与太平洋褶柔鱼秋生群的分布相一致。PDOI与产卵场Chl-a浓度无显著相关性(图4),但PDO正位相年份中产卵场Chl-a浓度年间变化较负位相时更为明显,且PDO正位相Chl-a浓度平均值为0.91 mg·m-3,要高于PDO负位相产卵场Chl-a浓度0.83 mg·m-3。这说明产卵场Chl-a浓度受PDO正位相影响更大。PDO位于正位相时,随着混合层深度的加深,浮游植物响应环境条件的变化存在经度方向上的差异[25],即浮游植物在经度方向上发生转移,进而产卵场Chl-a浓度发生变化。此外,PDOI与产卵场Ps也无显著相关性(图4),但PDO正位相年份中各年产卵场Ps显著低于负位相年份。以上结论表明,PDO模态位于正位相时,太平洋褶柔鱼秋生群产卵场SST较低,Chl-a浓度和Ps较高,导致CPUE变大,即太平洋褶柔鱼秋生群资源丰度高;相反,当PDO模态位于负位相时,产卵场SST较高,Chl-a浓度和Ps降低,导致CPUE变小,太平洋褶柔鱼秋生群资源丰度处于较低水平。
本文仅对太平洋褶柔鱼秋生群体9-11月(即产卵期)产卵场环境进行研究,未来研究还需要考虑全年环境数据,结合海表盐度、海流、海区深度等环境因子综合探索太平洋褶柔鱼资源丰度变化及关键影响因子。
参考文献:
[1] 杨林林,姜亚洲,程家骅.黄海南部太平洋褶柔鱼种群结构与繁殖生物学[J].生态学杂志,2010,29(6):1167-1174.YANG L L,JIANG Y Z,CHENG J H.Population structure and reproductive biology of Todarodes pacificus in Southern Yellow Sea[J].Chinese Journal of Ecology,2010,29(6):1167-1174.
[2] 唐峰华,刘尊雷,黄洪亮,等.日本海太平洋褶柔鱼生物学特征的年际变化[J].动物学杂志,2015,50(3):381-389.TANG F H,LIU Z L,HUANG H L,et al.Interannual variability of biology characteristics of Japanese Common Squid(Todarodes pacificus)in the Sea of Japan[J].Chinese Journal of Zoology,2015,50(3):381-389.
[3] 杨林林,姜亚洲,程家骅.东海太平洋褶柔鱼生殖群体的空间分布及其与环境因子的关系[J].生态学报,2010,30(7):1825-1833.YANG L L,JIANG Y Z,CHENG J H.Spatial distribution of mature stock of Pacific squid,Todarodes pacificus and its relation to environmental factors in the East China Sea[J].Acta Ecologica Sinica,2010,30(7):1825-1833.
[4] 李建生,严利平.东海太平洋褶柔鱼的数量分布及其与环境的关系[J].海洋渔业,2004,26(3):193-198.LI J S,YAN L P.Quantity distribution of Todarodes pacificus and its relation with environment in the East China Sea[J].Marine Fisheries,2004,26(3):193-198.
[5] 伊東祐方.総括 スルメイカの漁業,生活史および資源の現状の概要[C]//スルメイカ漁況予測精度向上のための資源変動機構に関する研究.東京:農林水產技術会議事務局,1972,4(9).ITOU T.Fisheries of Todarodes pacificus,Life history and current situation of resources[C]//Study on the accuracy of fishing forecasting of Todarodes pacificus.Tokyo:Norin Suisan Gijutsu Kaigi Jimukyoku,1972,4(9).
[6] SHIMURA T,SAKURA Y,UJIR,et al.Distribution of Egg-mass and paralarva of the common squid Todarodes pacificus near the Oki Islands[R].Tottori Prefectural Fisheries Experimental Station and Hokkaido University Faculty of Fisheries,2010(38):17-44.
[7] 董正之.黄海太平洋褶柔鱼资源现状与开发[J].海洋科学,1996,20(6):34-38.DONG Z Z.On the present status and exploratory problems of resource of Todarodes pacificus in the Yellow Sea[J].Marine Sciences,1996,20(6):34-38.
[8] 胡飞飞,陈新军.太平洋褶柔鱼秋生群资源补充量预报模型研究[J].广东海洋大学学报,2015,35(6):64-69.HU F F,CHEN X J.Study of forecasting model of recruitment for the autumn-spawning Group of Todarodes pacificus[J].Journal of Guangdong Ocean University,2015,35(6):64-69.
[9] 唐峰华,史贇荣,朱金鑫,等.海洋环境因子对日本海太平洋褶柔鱼渔场时空分布的影响[J].中国水产科学,2015(5):1036-1043.TANG F H,SHI Y R,ZHU JX,et al.Influence of marine environment factors on temporal and spatial distribution of Japanese common squid fishing grounds in the Sea of Japan[J].Journal of Fishery Sciences of China,2015(5):1036-1043.
[10] SAKURAI Y,KIYOFUJI H,SATOH S,et al.Changes in inferred spawning areas of Todarodes pacificus(Cephalopoda:Ommastrephidae)due to changing environmental conditions [J].ICES Journal of Marine Science,2000,57(1):24-30.
[11] SAKURAI Y,KIYOFUJI H,SATOH S I.Stock fluctuations of the Japanese common squid,Todarodes pacificus,related to recent climate changes(Proceedings of international commemorative symposium 70th anniversary of the Japanese society of fisheries science)[J].Fisheries Science,2002,68(23):226-229.
[12] 王亚民,李 薇,陈巧媛.全球气候变化对渔业和水生生物的影响与应对[J].中国水产,2009,397(1):21-24.WANG Y M,LI W,CHEN Q Y.Impacts and response of global climate change on fisheries and aquatic organism[J].China Fisheries,2009,397(1):21-24.
[13] 桜井泰憲.スルメイカの繁殖生態と気候変化に応答する資源変動[J].水產振興,2014(48):1-54.SAKURAI T.Variation in reproductive ecology and climate and their impacts on the squid abundance[J].Fisheries Revitalization,2014(48):1-54.
[14] 余 为,陈新军,易 倩.西北太平洋海洋净初级生产力与柔鱼资源量变动关系的研究[J].海洋学报,2016,38(2):64-72.YU W,CHEN X J,YI Q.Relationship between spatio-temporal dynamics of neon flying squid Ommastrephes bartraamii and net primary production in the northwest Pacific Ocean [J].Acta Oceanologica Sinica,2016,38(2):64-72.
[15] 徐 冰,陈新军,李建华.海洋水温对茎柔鱼资源补充量影响的初探[J].上海海洋大学学报,2012,21(5):878-883.XU B,CHEN X J,LI JH.Preliminary study on the influence of water temperature on the recruitment of Dosidicus gigas[J].Journal of Shanghai Ocean University,2012,21(5):878-883.
[16] 唐峰华,崔雪森,樊 伟,等.北太平洋柔鱼渔获量与海洋环境关系的遥感学研究[J].海洋技术学报,2011,30(2):7-12.TANG F H,CUI X S,FAN W,et al.Study on relationship of neon flying squid yield in north Pacific Ocean with marine environment by remote sensing[J].Ocean Technology,2011,30(2):7-12.
[17] 宋来军,屈 笑,袁成玉.朝鲜东海岸海域太平洋褶柔鱼资源及渔场分析[J].大连海洋大学学报,2008,23(2):123-127.SONG L J,QU X,YUAN C Y.Analysis of fishing ground and resources of Todarodes pacificus squid in eastern coast of Korea[J].Journal of Dalian Ocean University,2008,23(2):123-127.
[18] 沈新强,王云龙,袁 骐,等.北太平洋鱿鱼渔场叶绿素a分布特点及其与渔场的关系[J].海洋学报,2004,26(6):118-123.SHEN X Q, WANG Y L, YUAN Q, et al.Distributional characteristics of chlorophyll a and relation to the fishing ground in the squid fishing ground of the northern Pacific Ocean[J].Acta Oceanologica Sinca,2004,26(6):118-123.
[19] 木所英昭.気候変化に対するスルメイカの日本海での分布回遊と資源量変動に関する研究[R].水產総合研究センター研究報告,2009(27):95-189.KIMURA H.Impact of climatic changes on the distribution,migration pattern and stock abundance of the Japanese common squid,Todarodes pacificus in the Sea of Japan[R].Bulletin of Fisheries Research Agency,2009(27):95-189.
[20] NAKATA J.Long-term changes in catch and biological features of Japanese common squid(Todarodes pacificus)in waters off the east coast of Hokkaido[J].Recent Advances in Cephalopod Fisheries Biology,1993:343-350.
[21] 浜部基次.日本海西南海域を主にしたスルメイカの生態学的研究[R].日本海区水產研究所研究報告,1966(16):13-55.HAMABE M.Study on ecology of Todarodes pacificus in southwestern Japan Sea[R].Bulletin of the Japan Sea National Fisheries Research Institute,1966(16):13-55.
[22] 中村好和.東北北海道太平洋海域に来遊したスルメイカの資源量推定[C]//イカ類資源漁海況検討会議研究報告 (平成6年度).北海道区水產研究所,1996:7-17.NAKAMURA S.Stock assessment of the migratory squid in the Pacific Ocean,Hokkaido[C]//Report of fisheries status review conference about cephalopods.Hokkaido Fisheries Research Institute,1996:7-17.
[23] 木所英昭,檜山義明.日本海におけるスルメイカの分布海域による成長の差異[R].日本海区水產研究所研究報告,1996(46):77-86.KIDOKO E,HINOKIYAMA G.Growth difference of regional distribution of Todarodes pacificus in the sea of Japan[R].Bulletin of the Japan Sea National Fisheries Research Institute,1996(46):77-86.
[24] MANTUA N,HARE S,ZHANG Y,et al.A Pacific interdecadal climate oscillation with impacts on salmon production[J].Bulletin of the American Meteorological Society,1997,78(6):1069-1079.
[25] SISWANTOE,HONDA M,SASAI Y,et al.Meridional and seasonal footprints of the Pacific Decadal Oscillation on phytoplankton biomass in the northwestern Pacific Ocean [J].Journal of Oceanography,2016,72(3):465-477.
