植被类型对土壤非水稳性团聚体交换性盐基离子分布的影响
2018-05-07伍炫蓓何淑勤王家沐赵弘益
伍炫蓓,何淑勤,2*,王家沐,石 畅,赵弘益
(1.四川农业大学林学院,成都 611130;2.四川农业大学水土保持与荒漠化防治重点实验室,成都 611130)
团聚体不仅是构成土壤的基本单元[1],同时也是土壤养分贮存和物质循环的主要场所[2]。由于土壤孔隙的分布和连续性受团聚体数量分布和空间排列方式的影响,导致土壤的水力性质存在差异[3],而不同粒径大小的团聚体在养分的保持和供应方面发挥不同的作用[4],从而决定着土壤质量的优劣和肥力的高低[5-6]。土壤盐基离子作为评断土壤质量的标准之一,其含量和比例关系一定程度上能够反映了盐基离子的生物有效性以及土壤保肥、供肥和缓冲能力[7-9]。土壤是植被生长的基础,可为植被提供必要的养分,植被的变化也影响着土壤的形成与发育[10-11]。不同植被条件下土壤团聚体中的营养元素含量及分布会受到植物循环的影响[12]。已有研究表明,土壤中的交换性盐基离子含量与植被的吸收利用呈相关性[13],土壤交换性盐基离子在剖面分布能够反映营养元素被植物吸收情况和淋溶作用[14]。目前,关于土壤团聚体中元素分布的研究多集中在有机碳、全氮、全磷等方面[15-17],且以土地利用方式、人为管理措施对其分布的影响为主[18-20],而对不同植被条件下土壤团聚体中交换性盐基离子分布的研究较缺乏。
杂谷脑河流域属于典型的山地森林/干旱河谷交错带,是我国典型的生态脆弱区,区域内地形、地貌复杂多样,地质活跃,气候与植被沿海拔高度的变化明显,其生态环境状况与成都平原以及整个长江上游的生态可持续发展与社会经济发展有密切联系。长期以来人类不合理的资源开发,导致该区域生态系统遭到严重破坏,引发水土流失严重,自然灾害频发等问题。而有关这一区域的土壤团聚体中交换性盐基离子分布特征研究较少。基于此,本文选取四川西部理县杂谷脑河支流流域内典型的天然次生林、灌草林、乔灌林、混交幼林、岷江柏幼林和退耕岷江柏林6种植被条件土壤作为研究对象,采用野外调查与室内分析相结合的方法,分析不同植被条件下土壤团聚体交换性K+、Ca2+、Mg2+含量及分布特征,以期为杂谷脑河流域区域植被恢复以及可持续利用提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于四川省西部杂谷脑河流域,隶属于理县甘堡乡,该区域属山地季风气候,具有明显干湿季、日夜温差较大的特点,是典型的干旱河谷区。该区域最低海拔1 400 m,最高海拔5 922 m,平均海拔2 700 m,年降水量584 mm,年蒸发量1 399 mm;年最低极温-20℃左右,最高极温37℃左右,年均温12℃左右;该地土壤类型主要为山地棕壤、山地褐土。植被主要以灌丛和低覆盖草地为主[21];植被类型垂直分布情况由下至上分别为退耕岷江柏林、岷江柏(人工幼林)、混交幼林、乔灌草、灌丛和天然次生林,其基本情况见表1。

表1 试验样地基本情况Table 1 Basic characteristic of sample plots
1.2 试验方法
1.2.1 样地选择与样品采集
试验于2015年5月—2016年6月进行,在野外调查的基础上,根据试验区内植被的变化和地理位置进行综合考虑,选出6种典型植被,每种植被类型设置3个样地,分别在样地等分设置6个20 m×20 m的乔木样方、沿对角线设置6个5 m×5 m的灌木样方和1 m×1 m的草本样方。每一样地内采用“S”型5点取样,分别采集0~10、10~20 cm原状土,用于测定土壤非水稳性团聚体。同时,采集混合土样,用于测定土壤基本理化性质(见表2)。
1.2.2 测定方法
将混合土样在室内自然风干后,去除小石块、植被杂质等,用沙维诺夫干筛法[22]分离出>5、5~2、2~1、1~0.5、0.5~0.25 mm 和<0.25 mm 共 6 级非水稳性团聚体。交换性K+采用火焰光度法测定,交换性Ca2+和Mg2+采用原子吸收法测定[23]。
1.2.3 数据与分析
采用SPSS 19.0和EXCEL 2010软件进行数据处理、作图与分析。

表2 试验样地土壤基本性质Table 2 Soil properties of sample plots
2 结果与分析
2.1 土壤非水稳性团聚体的组成和分布情况
由表3可知,不同植被条件对土壤团聚体的数量和分布均有一定的影响。不同植被条件下,团聚体总体随粒径的减小呈先减少后增加的趋势。>2 mm粒径团聚体数量远大于其他粒径的团聚体,约占团聚体的62.5%,其中乔灌林含量最高(84.41%,82.39%),而退耕岷江柏林含量最低(37.44%,50.89%),在0~10 cm土层中,除灌草林5~2 mm粒径团聚体含量最高外,其余植被条件下均为>5 mm粒径团聚体含量最高,而在10~20 cm土层中,除灌草林和混交幼林5~2 mm粒径团聚体含量最高外,其余植被条件下均为>5 mm粒径团聚体含量最高。不同植被条件下大团聚体含量占主导地位,以乔灌林最高(96.30%,96.03%)天然次生林次之(94.21%,96.11%),退耕岷江柏林最低(73.24%,79.29%),除乔灌林大团聚体含量随土层深度的加深而降低外,其余植被条件均随土层深度的加深而增加。天然林较退耕林地及人工幼林受人为活动少,林分丰富,腐殖质层较厚,对土壤养分的供应能力更强。

表3 不同植被条件下非水稳性团聚体的分布特征Table 3 Distribution of soil non-water-stable aggregates under different vegetation types
2.2 土壤非水稳性团聚体中交换性K+的分布
由图1可知,在0~10 cm土层,随团聚体粒径的变化,各植被条件下土壤团聚体中交换性K+含量分布无明显规律,其中天然次生林、灌草林、乔灌林和退耕岷江柏林在2~1 mm粒径团聚体中交换性K+含量最高,而混交幼林和岷江柏幼林在>5 mm粒径团聚体中交换性K+含量最高,6种植被条件均在0.5~0.25 mm团聚体交换性K+含量最低。
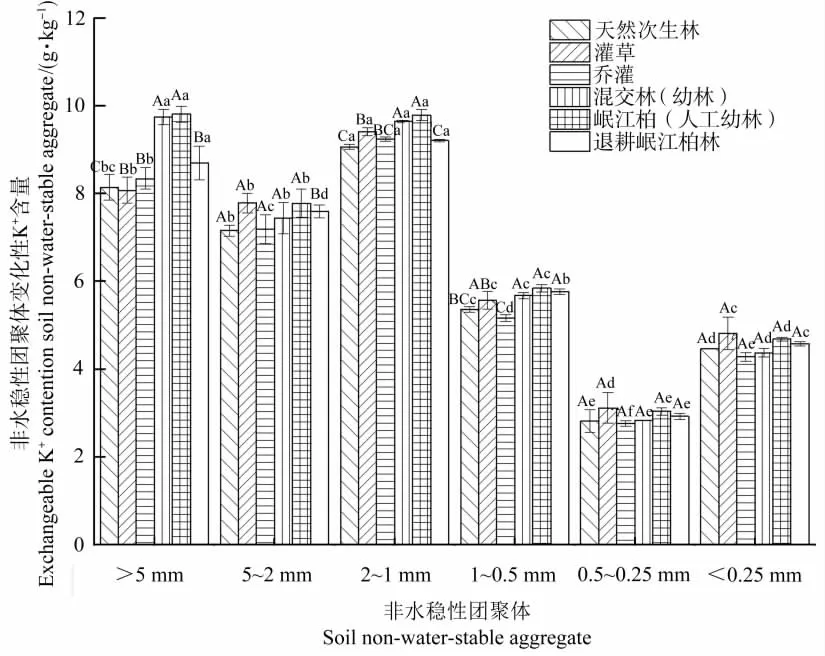
图1 不同植被条件下0~10 cm土层土壤团聚体中交换性K+的分布特征Figure 1 Distribution of exchangeable K+content in soil non-water-stable aggregates under different vegetation types at soil depth of 0~10 cm

图2 不同植被条件下10~20 cm土层土壤团聚体中交换性K+的分布特征Figure 2 Distribution of exchangeable K+content in soil non-water-stable aggregates under different vegetation types at soil depth of 10~20 cm
由图2可知,在10~20 cm土层,随团聚体粒径减小,天然次生林中交换性K+含量呈先增加后降低的趋势,其余植被条件交换性K+分布特征各异,其中混交幼林和岷江柏幼林在>5 mm粒径团聚体中交换性K+含量最高,而其余植被条件下均为在2~1 mm粒径团聚体中交换性K+含量最高。
不同植被条件下各粒径团聚体中交换性K+含量之间无明显差异,说明土壤团聚体中交换性K+含量受植被类型的影响不大。通过对比不同土层深度团聚体中交换性K+含量可发现,在0~10 cm土层中,除0.5~0.25 mm粒径外,其余粒径团聚体中交换性K+含量均高于10~20 cm土层的相同粒径团聚体中交换性K+含量,而且各植被条件下0.5~0.25 mm粒径团聚体中交换性K+含量随土层深度的加深而增加。同时,通过比较不同粒径团聚体中交换性K+的含量分布可知,5~1 mm粒径团聚体中交换性K+含量明显大于>1 mm粒径团聚体中交换性K+含量,说明较大粒径团聚体具有较强供应交换性K+的能力。
2.3 土壤非水稳性团聚体中交换性Ca2+的分布
由图3可知,在0~10 cm土层,除灌草林和乔灌林交换性Ca2+含量随团聚体粒径的减小先降低后增加外,其余植被条件下土壤团聚体中交换性Ca2+分布特征各异且粒径之间差异显著。其中灌草林、混交幼林、岷江柏幼林和退耕岷江柏林在<0.25 mm粒径团聚体中交换性Ca2+含量最高,天然次生林在0.5~0.25 mm粒径团聚体中交换性Ca2+含量最高,而乔灌林在>5 mm粒径团聚体中交换性Ca2+含量最高。同时,除天然次生林在>5 mm粒径团聚体中交换性Ca2+含量最低外,其余植被条件均在2~1 mm粒径团聚体中交换性Ca2+含量最低。
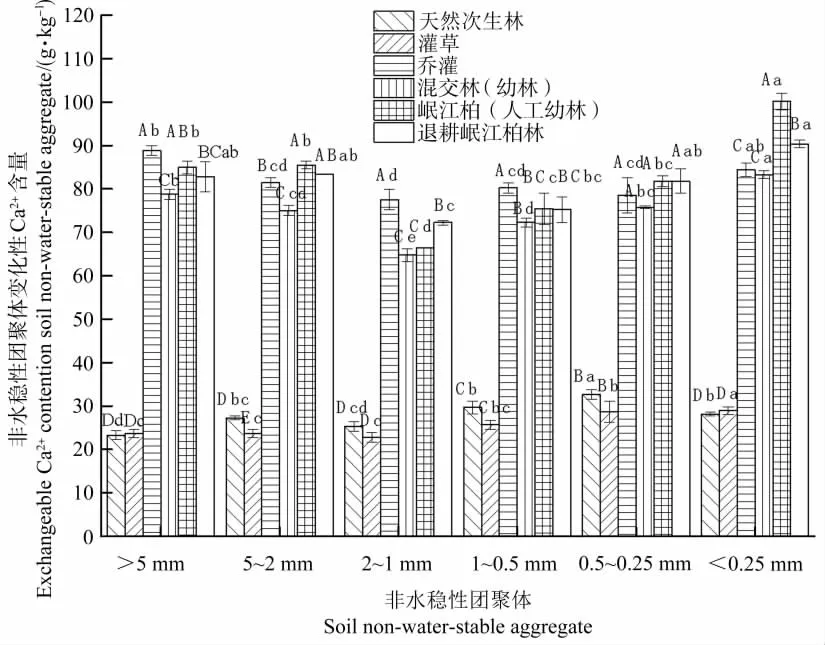
图3 不同植被条件下0~10 cm土层土壤团聚体中交换性Ca2+的分布特征Figure 3 Distribution of exchangeable Ca2+content in soil non-water-stable aggregates under different vegetation types at soil depth of 0~10 cm
由图4可知,在10~20 cm土层,随团聚体粒径的减小,天然次生林交换性Ca2+含量先增加后降低,在0.5~0.25 mm粒径团聚体中交换性Ca2+含量达到最高,而在>5 mm粒径团聚体中交换性Ca2+含量最低,而灌草林、乔灌林、混交幼林及岷江柏幼林交换性Ca2+含量随团聚体粒径的减小呈先降低后增加的趋势,在2~1 mm粒径团聚体中交换性Ca2+含量最低,其中灌草林、乔灌林和混交幼林在<0.25 mm粒径团聚体中交换性Ca2+含量最低,而岷江柏幼林含量在>5 mm粒径团聚体中交换性Ca2+含量最低。退耕岷江柏林交换性Ca2+含量随团聚体粒径的变化无明显规律。
通过对比不同土层深度团聚体中交换性Ca2+含量可发现,在0~10 cm土层中,各粒径团聚体中交换性Ca2+增幅最低185.3%,最高282.4%,说明土壤团聚体中交换性Ca2+含量受植被条件影响较显著;在10~20 cm土层中,各粒径团聚体中交换性Ca2+增幅较0~10 cm土层更大,最高可达到318.3%,说明随着土层深度的加深,土壤团聚体中交换性Ca2+含量变化受植被条件的影响更加明显。
2.4 土壤非水稳性团聚体中交换性Mg2+的分布
由图5可知,在0~10 cm土层,随团聚体粒径的变化,各植被条件下土壤团聚体中交换性Mg2+含量分布无明显规律。天然次生林在2~1 mm粒径团聚体中交换性Mg2+含量最高,在5~2 mm粒径团聚体中交换性Mg2+含量最低。灌草林、乔灌林、混交幼林和岷江柏幼林在>5 mm粒径团聚体中交换性Mg2+含量最高,其中乔灌林、混交幼林和岷江柏幼林在1~0.5 mm粒径团聚体中交换性Mg2+含量最低,而灌草林在5~2 mm粒径团聚体中交换性Mg2+含量最低。退耕岷江柏林在5~2 mm粒径团聚体中交换性Mg2+含量最高,在1~0.5 mm粒径团聚体中交换性Mg2+含量最低。

图4 不同植被条件下10~20 cm土层土壤团聚体中交换性Ca2+的分布特征Figure 4 Distribution of exchangeable Ca2+content in soil non-water-stable aggregates under different vegetation types at soil depth of 10~20 cm

图5 不同植被条件下0~10 cm土层土壤团聚体中交换性Mg2+的分布特征Figure 5 Distribution of exchangeable Mg2+content in soil non-water-stable aggregates under different vegetation types at soil depth of 0~10 cm
由图6可知,在10~20cm土层,随团聚体粒径的变化,各植被条件下土壤团聚体中交换性Mg2+含量分布同样无明显规律。各植被条件均在<0.25 mm粒径团聚体中交换性Mg2+含量最高,其中天然次生林、灌草林在2~1 mm粒径团聚体中次之,其余植被条件下在>5 mm粒径团聚体中次之,同时,各植被条件均在5~2 mm粒径团聚体中交换性Mg2+含量最低。
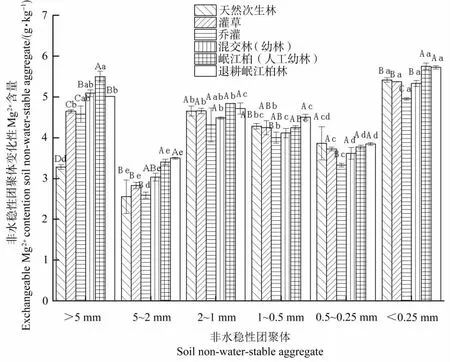
图6 不同植被条件下10~20 cm土层土壤团聚体中交换性Mg2+的分布特征Figure 6 Distribution of exchangeable Mg2+content in soil non-water-stable aggregates under different vegetation types at soil depth of 10~20 cm
通过对比不同土层深度团聚体中交换性Mg2+含量可发现,在0~10 cm土层,5~2 mm粒径团聚体中交换性Mg2+含量增幅最大,>5 mm粒径团聚体中增幅次之,说明植被条件对较大团聚体中交换性Mg2+含量的影响较大;在10~20 cm土层中,各粒径团聚体中交换性Mg2+含量随植被条件变化的分布规律与其在0~10 cm土层相似。随土层深度的加深,除1~0.5 mm和0.5~0.25 mm粒径团聚体中交换性Mg2+含量增加外,其余粒径团聚体中交换性Mg2+含量均降低。
3 讨论
本研究表明,杂谷脑河流域不同植被条件下,各土层团聚体中交换性K+含量岷江柏幼林最高,天然次生林最低;交换性Ca2+含量在0~10 cm土层中岷江柏幼林含量最高,天然次生林最低,在10~20 cm土层退耕岷江柏林含量最高,灌草林最低;交换性Mg2+含量在0~10 cm土层中岷江柏幼林含量最高,天然次生林最低,在10~20 cm土层中退耕岷江柏林含量最高,乔灌林最低。本研究对土壤团聚体中交换性盐基离子分布的研究中表明,交换性K+、Ca2+和Mg2+的含量在>0.25mm大团聚体中分布较多,均高于在<0.25 mm微团聚体中的含量,与曾路生等[24]的研究结果一致。不同植被条件下土壤团聚体总体随粒径的减小呈先降低后增加的趋势,与郑子成等[25]的研究结果一致。各粒径团聚体中交换性盐基离子平均含量呈现出 Ca2+>K+>Mg2+,与曾路生等[24]和王晟强等[26]Ca2+>Mg2+>K+的研究结果不一致,这可能与种植条件、人为管理模式有关。曾路生等[24]研究对象为设施栽培条件土壤,实施“高肥”的管理模式,施肥量是露地土壤的3~4倍,施肥不仅可增加团聚体有机胶结物质[27],同时也改善了土壤结构[28],进而促进Ca2+和Mg2+的积累,同时由于设施栽培条件下土壤pH有所降低,呈潜在酸化趋势,有利于土壤中过剩的K+与土壤胶体上Ca2+进行置换,故导致K+平均含量较低。王晟强等[26]研究对象为茶园土壤,土壤植茶后,同样土壤呈酸化趋势,再加上人为定期施肥的影响,故与本研究结果不一致。而本文所研究的区域,以自然植被恢复为主,Ca2+、Mg2+、K+含量主要与成土母质与植被类型有关,且土壤pH均较高(见表1),故出现上述结果。不同植被条件下交换性K+和Ca2+在各粒径中分布呈显著差异,且无明显变化规律,交换性Mg2+在各粒径中分布较平均,但也无明显变化规律,与王晟强等[26]交换性K+含量随粒径的减小先增加后降低、交换性Ca2+含量随粒径的减小而增加、交换性Mg2+含量在各粒径中分布较均匀的研究结果相差较大,植被类型是造成这一差异的主要因素。因此,在研究区,不仅要选择合理的植被,同时也应通过施肥等土壤改良措施,改善土壤团聚体数量和质量,以增强土壤的供肥保肥能力。
4 结论
①不同植被条件下,以>2 mm大团聚体占主体地位,各植被条件团聚体含量总体表现为:乔灌>天然次生林>灌草>混交林(幼林)>岷江柏(幼林)>退耕岷江柏林;随团聚体粒径的减小,团聚体含量总体呈现先降低后增加的趋势。
②不同植被条件下Ca2+、K+在各粒径中分布呈显著差异,且无明显变化规律,Mg2+在各粒径中分布较均匀。各粒径团聚体中交换性盐基离子平均含量呈现出Ca2+>K+>Mg2+,表明不同交换性盐基离子分布受团聚体粒径大小的影响各异,交换性Ca2+较交换性K+、Mg2+更易在研究区土壤团聚体中累积。
③不同植被不仅影响着各粒径团聚体的分布,同时也影响着不同粒径团聚体中Ca2+、K+、Mg2+的分布。在研究区植被恢复过程中,不仅要选择合理的植被,同时也应改善土壤团聚体数量和质量,以增强土壤的供肥保肥能力。
参考文献:
[1]李玮,郑子成,李廷轩,等.不同植茶年限土壤团聚体及其有机碳分布特征[J].生态学报,2014,34(21):6326-6336.
[2]SIX J,CONANT R T,PAUL E A,et al.Stabilization mechanisms of soil organic matter:Implications for C-saturation of soils[J].Plant and Soil,2002(41):155-176.
[3]袁俊吉,蒋先军,胡宇,等.不同植被覆盖对养分在土壤水稳性团聚体中分布特征的影响[J].水土保持学报,2009,23(6):112-118.
[4]陈恩凤,周礼恺,武冠云.微团聚体的保肥供肥性能及其组成比例在评判土壤肥力水平中的意义[J].土壤学报,1994,31(1):18-25.
[5]何淑勤,郑子成.不同土地利用方式下土壤团聚体的分布及其有机碳含量的变化[J].水土保持通报,2010,30(1):7-10.
[6]何淑勤,郑子成,宫渊波.不同退耕模式下土壤水稳性团聚体及其有机碳分布特征[J].水土保持学报,2011,25(5):229-233.
[7]刘晓利,何园球.不同利用方式和开垦年限下红壤水稳性团聚体及养分变化研究[J].土壤,2009,41(1):84-89.
[8]OVALLES F A,COLLINS M E.Soil-landscape relationships and soil variability in north central Florida[J].Soil Science Society of America Journal,1986,50(2):401-408.
[9]范庆峰,虞娜,张玉玲,等.设施蔬菜栽培对土壤阳离子交换性能的影响[J].土壤学报,2014,51(5):1132-1137.
[10]BINKLEY D,GIARDINA C.Why do tree species affect soils?The warp and woof of tree soil interactions.Biogeochemistry[J].Biogeochemistry,1998,42(1-2):89-106.
[11]杨宁,邹冬生,杨满元,等.衡阳紫色土丘陵坡地不同恢复阶段植被特征与土壤性质的关系[J].应用生态学报,2013,24(1):90-96.
[12]姜勇,郝伟,张玉革,等.潮棕壤不同利用方式营养元素随剖面深度的变化特征[J].水土保持学报,2006,20(3):93-97.
[13]鲁艳红,廖育林,聂军,等.长期施用氮磷钾肥和石灰对红壤性水稻土酸性特征的影响[J].土壤学报,2016,53(1):202-212.
[14]何淑勤,宫渊波,郑子成,等.不同植被条件下土壤团聚体交换性 K+、Na+的分布特征[J].水土保持学报,2014,28(6):228-233.
[15]魏亚伟,苏以荣,陈香碧,等.人为干扰对喀斯特土壤团聚体及其有机碳稳定性的影响[J].应用生态学报,2011,22(4):971-978.
[16]陈晓芬,李忠佩,刘明,等.不同施肥处理对红壤水稻土团聚体有机碳、氮分布和微生物生物量的影响[J].中国农业科学,2013,46(5):950-960.
[17]郭万伟,肖和艾,吴金水,等.红壤旱土和水稻土团聚体中磷素的分布特点[J].土壤学报,2009,46(1):85-92.
[18]胡宁,娄翼来,张晓珂,等.保护性耕作对土壤交换性盐基组成的影响[J].应用生态学报,2010,21(6):1492-1496.
[19]刘希玉,王忠强,张心昱,等.施肥对红壤水稻土团聚体分布及其碳氮含量的影响[J].生态学报,2013,33(16):4949-4955.
[20]李鉴霖,江长胜,郝庆菊.土地利用方式对缙云山土壤团聚体稳定性及其有机碳的影响[J].环境科学,2014,35(12):4695-4704.
[21]黄海,石胜伟,谢忠胜.杂谷脑河下游坡面泥石流发育特征及防治对策[J].水土保持研究,2013,20(6):111-116.
[22]文倩,赵小蓉,陈焕伟,等.半干旱地区不同土壤团聚体中微生物量碳的分布特征[J].中国农业科学,2004,37(10):1504-1509.
[23]鲁如坤.土壤农业化学分析方法[M].中国农业科技出版社,2000:30-34.
[24]曾路生,高岩,李俊良,等.寿光大棚土壤团聚体中交换性盐基离子组成与土壤团聚体性关系[J].水土保持学报,2011,25(5):224-228.
[25]郑子成,王永东,李廷轩,等.退耕对土壤团聚体稳定性及有机碳分布的影响[J].自然资源学报,2011,26(1):119-126.
[26]王晟强,郑子成,李廷轩,等.植茶年限对土壤团聚体中交换性盐基离子分布的影响[J].土壤学报,2013,50(5):1013-1020.
[27]柳开楼,黄晶,张会民,等.长期施肥对红壤旱地团聚体特性及不同组分钾素分配的影响[J].土壤学报,2018,55(2):443-454.
[28]王经纬,姚怡,徐江兵,等.长期施肥对旱地红壤团聚体磷素固持与释放能力的影响[J].土壤学报,2017,54(5):1240-1250.
