中国北方粳稻基于SNPs的遗传结构分析和主要农艺性状的变化
2018-05-07王嘉宇徐正进隋国民
高 虹,徐 铨,王嘉宇,钟 鸣,徐正进,隋国民
(1.辽宁省农业科学院,辽宁 沈阳 110101;2.沈阳农业大学 水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,北方超级粳稻育种教育部重点实验室,辽宁 沈阳 110866;3.辽宁省水稻研究所,辽宁 沈阳 110000;4.沈阳农业大学 生物科学技术学院,辽宁省农业生物技术重点实验室,辽宁 沈阳 110866)
近年来,北方粳稻育种主要通过引入不同生态型水稻基因资源培育粳稻品种。在“理想株型”和“籼粳杂交”优势理论的指导下,中国北方现代粳稻育成品种产量不断提高,但随之遗传多样性和遗传结构也产生了变化[1]。Sun等[2]研究表明,20世纪90年代以后,北方粳稻基于SSR分子标记分析的群体遗传多样性变得更加丰富了,是由于籼粳杂交在引入籼稻优良血缘的同时固定粳稻优良等位的基因。由此产生的基因组构成的改变是一个重要的进化生物学问题。前人对中国粳稻遗传多样性和遗传结构的研究,多基于SSR位点和籼粳特异位点的变异[3-7],虽然变异丰富但不足以精确解析遗传背景相似粳稻栽培品种的群体结构。大多数研究的品种多集中在老品种和地方品种,因此,不太可能受到人工杂交的影响,从而保持了原有的遗传特点[8]。目前,分析品种的多样性和遗传结构,比较基因组序列的遗传分化,可以使用高分辨率的单核苷酸多态性(SNP)标记[9-14]。
本研究将选择的北方粳稻品种分为3个部分(按不同育种阶段划分):早期品种(中国北方早期耕种的农家品种)、现代品种(现在生产实践上推广的粳稻育成品种)、外来品种(育种过程中引入的日本、韩国、朝鲜等粳稻品种),利用189个平均分布基因组的功能SNPs分析北方粳稻的遗传多样性和群体结构,结合现代粳稻育成品种主要农艺性状的变化,研究育种行为对北方粳稻基因组遗传基础的影响和对产量及生理形态特性的影响,旨在为北方粳稻进一步育种选择提供参考方向。
1 材料和方法
1.1 试验材料
试验材料共109份:中国北方现代品种(现在生产实践上主要推广的粳稻育成品种)33份,早期品种(中国北方早期耕种的农家品种)26份,外来品种(育种过程中引入的日本、韩国、朝鲜等粳稻品种)25份,作为对照样本的籼稻品种25份。
1.2 试验方法
于2013年在沈阳农业大学水稻研究所试验田种植粳稻,将株高和生育期相近的材料邻近种植,2次重复。3行区,行长1.5 m,栽植密度为30.0 cm×13.3 cm。4月12日育苗,5月20日移栽,人工插秧,每穴单苗。试验田为沙壤土,(NH4)2HPO4、KCl作为底肥施用,施用量分别为300,225 kg/hm2,尿素作为底肥、分蘖肥和粒肥施用,施用量分别为187.5,112.5,75.0 kg/hm2。
1.3测定项目及方法
1.3.1 试验材料DNA提取 取试验材料秧苗叶片,利用CTAB法提取基因组DNA。
1.3.2 SNPs位点的设计与检测 根据Huang等[15]对籼粳稻核心材料高通量测序后的分析结果设计了189个SNPs位点。利用美国 SEQUENOM 公司的时间飞行质谱技术(MassARRAY)检测试验材料的基因型,委托华大基因完成。
1.3.3 农艺性状测定 于齐穗期选取每个小区长势中等的单茎5个,利用徒手切片法分别观察穗颈大维管束数目、穗颈小维管束数目。抽穗后30 d 每个株系选取5个单茎测定株高、穗长、穗颈弯曲度及各节间长度,上三叶叶长、叶宽。成熟期调查每小区中间1行植株的有效穗数,按平均有效穗数割取5株,风干后称量穗重,按一次枝梗众数法选取其中10穗,分别对穗粒数、结实率、千粒质量进行测定。
1.4 数据分析
利用POPGENE 2.0计算样本间基于189个功能SNPs位点变异的多样性指数。利用Structure version 2.3.1分析群体结构。利用MEGA 6.0软件绘制Neighbor-joining系统进化树。数据采用SPSS 19.0软件进行统计分析及显著性检测。
2 结果与分析
2.1 基于功能 SNPs位点变异的群体遗传多样性分析
分析中国北方粳稻基于189个功能SNPs位点变异的群体遗传多样性。结果表明(图1):韩国和日本等外来引入品种有效等位基因数(1.111 2)和遗传多样性指数(0.115 1)最高,北方早期老品种的等位基因数最高(1.366 0),现代粳稻育成品种的等位基因数(1.149 5)、有效等位基因数(1.055 1)、遗传多样性指数(0.053 2)均最低,并且遗传多样性指数极显著低于外来品种与早期品种,群体遗传多样性指数由高到低分别为:外来品种、早期品种、现代品种。

不同大小写字母分别表示差异达到1%和5%显著水平。Different uppercase and lowercase letters indicate significant difference at 1% and 5% levels,respectively.
2.2 基于功能SNPs位点变异的群体遗传结构分析
分析3个不同育种时期和来源的粳稻与对照籼稻的群体遗传结构,结果表明(图2),当k=2时,Δk值达到最大,籼稻组群与粳稻组群产生了明显的分化,当k=3时,北方现代粳稻品种独立成群,与另外2个粳稻组群发生了分化,当k=4时,早期粳稻品种内部群体结构出现了分化,外来粳稻和现代粳稻没有分化,仍然保持各自为单独的一个类群;当k=5时,仍然是早期粳稻品种产生了进一步的遗传结构分化。群体结构划分结果说明:北方现代粳稻品种的群体结构与早期品种和外来品种产生了一定分化而且更为独立,虽然早期粳稻品种内部遗传结构较为复杂,但相对于现代品种来说,与外来品种有着更加紧密的群体遗传构成。
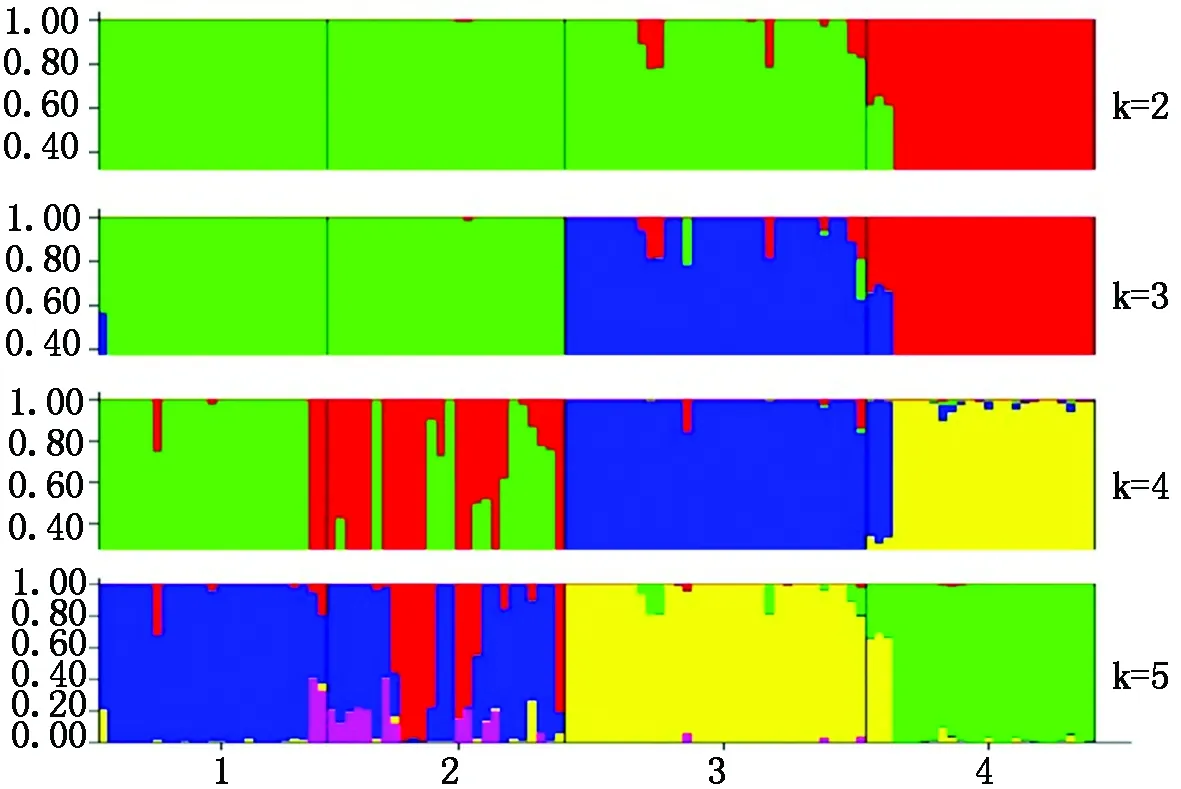
1.外来粳稻品种;2.早期粳稻品种;3.现代粳稻品种;4.籼稻品种。1.Exotic japonica;2.Early breeding phase japonica;3.Modern breeding phase japonica;4.Indica rice varieties.
为了进一步明确外来和籼型基因的引入以及育种行为对北方现代粳稻育成品种群体遗传结构的影响,构建了基于功能SNPs变异的系统进化树如图3所示。籼稻样本与外来粳稻品种和早期的粳稻遗传距离最远,说明在功能位点变异水平上的遗传分化属于亚种间的分化。虽然北方现代粳稻与籼稻还保持着一定的遗传距离和明显的界限,相对于关系较为紧密的早期品种和外来品种,现代品种随着籼型基因引入北方早期农家老品种并不断进行选择和淘汰后,已经逐渐脱离传统的粳稻基因组背景,形成了新的独立的遗传结构。
2.3 中国北方现代粳稻育成品种主要农艺性状的变化
对3个不同育种时期和来源的中国北方粳稻与对照籼稻的产量及产量构成因素分析比较,结果表明(表1),现代粳稻育成品种产量最高,比外来粳稻增产11.12%,比早期粳稻增产38.13%,且差异达到极显著水平。与外来粳稻品种相比,现代粳稻育成品种的有效穗数显著减少,但穗粒数显著高于早期粳稻和外来粳稻(增幅20%),同时现代粳稻育成品种的结实率最高,提高了6%~10%,且与早期粳稻达到了显著水平,虽然千粒质量极显著低于籼稻品种,但与早期粳稻和外来粳稻间差异不显著。从产量构成因素来看,北方现代粳稻育成品种穗粒数比早期粳稻和外来粳稻显著增多,且结实率较高是其增产的主要原因。

图3 中国北方粳稻与对照籼稻进化树聚类结果Fig.3 Neighbor-joining tree for japonica in Northern China and indica
3个不同育种阶段和来源的粳稻与对照籼稻株高及各节间长差异分析比较(表2),结果表明,各品种从基部至上各节间长度增加。株高、倒二节间长和倒三节间长表现出相同的变化趋势:早期品种>外来品种>现代品种>籼稻品种,现代粳稻育成品种平均株高比早期农家品种降低了20 cm左右,且二者间差异显著;现代粳稻倒一节间长度最短,低于早期粳稻、外来粳稻和籼稻品种,且与外来粳稻和早期粳稻间差异达到了显著和极显著水平;倒四节间长虽然低于早期粳稻,高于外来粳稻和籼稻品种,但差异并不显著。这说明倒一和倒三节间长度的降低对现代粳稻育成品种株高的影响较大。

表1 不同育种阶段水稻品种产量及产量构成因素的差异 Tab.1 Differences of yield and its components among rice varieties in different breeding phases and indica
注:同一列中数字后不同大小写字母分别表示差异达到1%和5%显著水平。表2-4同。
Note:Within a column,data followed by different uppercase and lowercase letters indicate significant difference at 1% and 5% levels,respectively. The same as Tab.2-4.

表2 不同育种阶段和来源水稻品种株高及各节间长度的差异Tab.2 Differences of plant height and each internode length among rice varieties in different breeding phases and indica cm
表3结果表明,不同育种阶段和来源的粳稻与对照籼稻剑叶、倒二叶、倒三叶长宽不同,现代粳稻育成品种剑叶、倒二叶、倒三叶叶长均最短,且与早期粳稻、外来粳稻和籼稻品种间差异达到了极显著水平;现代粳稻除剑叶宽小于籼稻外(差异不显著),倒二叶、倒三叶叶宽均为最宽,剑叶叶宽显著高于外来粳稻,倒二叶和倒三叶叶宽和早期粳稻与外来粳稻的差异分别达到了显著和极显著水平。现代粳稻育成品种功能叶叶片短而宽,有利于提高光能利用率,改善群体通风透光,对产量的增加有极大的贡献。
穗长是穗型的重要影响因素,本研究发现,现代粳稻育成品种穗长最短,穗弯曲角度最小,且与早期粳稻、外来粳稻和籼稻品种差异达到了显著或极显著水平,属于直立穗型 (表4)。现代粳稻穗颈大维管束数目大于早期粳稻、极显著大于外来粳稻、极显著小于籼稻品种;穗颈小维管束数目最多,且显著大于外来粳稻。穗颈维管束是物质向穗部运输的必经通道,维管束数目直接影响产物向籽粒的转移,现代粳稻育成品种穗颈大、小维管束数显著增加,说明光合产物运输系统比较发达,改善穗部性状,进而提高产量。

表3 不同育种阶段水稻品种功能叶片长宽的差异 Tab.3 Differences of leaf related traits among rice varieties in different breeding phases and indica cm

表4 不同育种阶段水稻品种穗型的差异 Tab.4 Differences of panicle related traits among rice accessions cultivates in different breeding phases
3 结论与讨论
3.1 中国北方粳稻基于功能 SNPs位点变异的群体遗传多样性
Sun等[2]研究结果表明,20世纪90年代后期,基于SSR分子标记分析的中国北方粳稻育成品种群体遗传多样性逐渐丰富,这可能与籼粳稻杂交育种90年代后期在我国北方开展有关,为中国北方粳稻群体遗传多样性的扩展做出了贡献。但本研究中,现代粳稻育成品种的遗传多样性指数却是最低,是由于SSR作为一种中性进化的位点其变异不受选择的制约,而本研究中的功能SNPs位点具有较大的遗传效应,由于育种过程中往往针对一个或几个育种目标进行选择,尤其是集中在几个核心亲本的利用,因此,在这一过程中功能基因位点频率逐渐下降,而中性进化位点的频率对此受到的影响较小。Yamamoto等[11]与Yonemaru等[13]研究日本农家老品种与育成品种的基因组遗传多样性结果表明,育成品种产生了一些新的单体型,但同时也失去了一些基因,而且多样性的丧失大于新单体型的产生。该结论与本研究结果相互印证,育种过程中通过人工和自然选择,选择优良基因加以利用,但在淘汰不利性状的同时损失了与其相连锁的性状,导致功能基因水平上遗传多样性的狭窄[16]。因此,在今后借助分子育种技术过程中应当注意,在利用不同生态型水稻优良基因进行品种改良时,应注意减轻基因多样性的损失。
3.2 中国北方粳稻基于功能 SNPs位点变异的群体遗传结构
以往研究大多根据地理生态差异划分水稻类型。随着分子标记技术的发展,亚洲栽培稻群体结构类群划分体系已经建立起来,对解析北方粳稻基因组构成具有很好的借鉴作用。Garris等[17]与Gaicedo等[18]利用SSR标记与STS变异位点将亚洲栽培稻划分为籼稻、奥斯稻、热带粳稻、温带粳稻以及香稻5类。Zhao等[19]利用SNPs基因芯片技术,进一步明确了亚洲栽培稻不同组群间基因相互渐渗区域。相对而言,日本粳稻的遗传改良在基因组成分上看并未发生本质的改变[10-11]。虽然我国北方与日本的主要粳稻产区维度相近,气候类型相似,但育种策略和选择的方向不同使基因组发展方向产生了差异[16]。我国北方现代粳稻育种不超过50年,但选择与生态环境压力已经将现代粳稻从传统粳稻中分离出来,已经将基因组塑造成了一个新的遗传结构与进化地位。
3.3 中国北方粳稻育成品种主要农艺性状的变化
20 世纪 80 年代以来,随着地理纬度相近的日本和韩国粳稻品种的引入和籼粳稻杂交育种的深入以及现代直立穗型育成品种的推广,中国北方粳稻遗传基础发生了变化,同时形态特征和产量结构也有明显不同。本研究中,现代粳稻育成品种比育种过程中引入的日本、韩国、朝鲜等外来粳稻品种的有效穗数显著减少,但穗粒数显著增加,产量最高,并且比早期耕种农家品种增产38.13%。与许多学者研究结果一致[20-23]。同时通过增加穗颈维管束数目,促进了物质运输能力,将结实率和千粒质量维持在较高水平,是北方粳稻超高产育种理论成功应用的体现。徐正进等[24]研究表明,株高平均值最佳范围基本在100~110 cm,北方现代粳稻育成品种成功将株高降低在最佳范围之内,有利于协调株高与生物产量之间的关系。早期农家品种穗型几乎都是弯穗型,随着外来粳稻引入,尤其是以辽粳5号为代表的直立穗型品种育成后,直立穗型逐渐成为北方现代粳稻育种方向之一[25],直立穗型品种叶片上冲直立,开张角度小,叶片短而宽和株型很好地结合,使植株的光合形态得到改良,气体扩散等生态条件优越。综上所述,在超高产遗传改良和超级稻育种实践的指导下,北方现代粳稻育成品种产量提高,株型生理特性得到改良,并且形成了独立的遗传结构,为进一步探索提高北方粳稻生产潜力提供科学依据与技术储备。
参考文献:
[1] 王海泽,刘 迪,唐 亮. 东北地区主栽水稻品种的籼型位点频率及其与产量的关系[J]. 沈阳农业大学学报,2014,45(6):661-667.
[2] Sun J,Liu D,Wang J Y,et al. The contribution of intersubspecific hybridization to the breeding of super-high-yieldingjaponicarice in Northeast China[J]. Theoretical and Applied Genetics,2012,125(6):1149-1157.
[3] 李红宇,侯昱铭,陈英华,等. 用SSR标记评估东北三省水稻推广品种的遗传多样性[J]. 中国水稻科学,2009,23(4):383-390.
[4] 张 科,魏海锋,卓大龙,等. 黑龙江省近年审定水稻品种基于SSR标记的遗传多样性分析[J]. 植物遗传资源学报,2016,17(3):447-454.
[5] 马作斌,王昌华,王 辉,等. 不同国家水稻品种的遗传多样性分析[J]. 植物遗传资源学报,2014,15(3):540-545.
[6] 刘 丹,王嘉宇,马殿荣,等. 东北地区杂草稻与栽培稻的遗传多样性及籼粳分化[J]. 植物遗传资源学报,2017,18(2):217-224.
[7] 杨玉蓉,孙建昌,王兴盛,等. 宁夏不同年代水稻品种的遗传多样性比较[J]. 植物遗传资源学报,2014,15(3):457-464.
[8] 邓宏中,王彩红,徐 群,等. 中国水稻地方品种与选育品种的遗传多样性比较分析[J]. 植物遗传资源学报,2015,16(3):433-442.
[9] Ebana K,Yonemaru J I,Fukuoka S,et al. Genetic structure revealed by a whole-genome single-nucleotide polymorphism survey of diverse accessions of cultivated Asian rice (OryzasativaL.)[J]. Breeding Science,2010,60(4):390-397.
[10] Nagasaki H,Ebana K,Shibaya T,et al. Core single-nucleotide polymorphisms-a tool for genetic analysis of the Japanese rice population[J]. Breeding Science,2010,60:648-655.
[11] Yamamoto T,Nagasaki H,Yonemaru J I,et al. Fine definition of the pedigree haplotypes of closely related rice cultivars by means of genome-wide discovery of single-nucleotide polymorphisms[J]. BMC Genomics,2010,11(1):267.
[12] Arai-Kichise Y,Shiwa Y,Nagasaki H,et al. Discovery of Genome-Wide DNA polymorphisms in a landrace cultivar of japonica rice by Whole-Genome sequencing[J]. Plant and Cell Physiology,2011,52(2):274-282.
[13] Yonemaru J I,Yamamoto T,Ebana K,et al. Genome-wide haplotype changes produced by artificial selection during modern rice breeding in Japan[J]. PLoS One,2012,7(2):e32982.
[14] 史亚兴,卢柏山,宋 伟,等. 基于 SNP 标记技术的糯玉米种质遗传多样性分析[J]. 华北农学报,2015,30(3):77-82.
[15] Huang X,Wei X,Sang T,et al. Genome-wide association studies of 14 agronomic traits in rice landraces[J]. Nature Genetics,2010,42(11):961-967.
[16] 高 虹. 亚种间杂交对东北粳稻的育种贡献[M]. 沈阳:沈阳农业大学,2013:27-28.
[17] Garris A J,Tai T H,Coburn J,et al. Genetic structure and diversity inOryzasativaL.[J]. Genetics,2005,169(3):1631-1638.
[18] Caicedo A L,Williamson S H,Hernandez R D,et al. Genome-wide patterns of nucleotide polymorphism in domesticated rice[J]. PLoS Genetics,2007,3 (9):e163.
[19] Zhao X,Yang L,Zheng Y,et al. Subspecies-specific intron length polymorphism markers reveal clear genetic differentiation in common wild rice (OryzarufipogonL.) in relation to the domestication of cultivated rice (O.sativaL.) [J]. Journal of Genetics and Genomics,2009,36(7):435-442.
[20] 龚金龙,邢志鹏,胡雅杰,等. 籼、粳超级稻产量构成特征的差异研究[J]. 核农学报,2014,28(3):500-511.
[21] 桂君梅,王林友,范小娟,等. 基于InDel分子标记的籼粳杂交稻与粳粳杂交稻的杂种优势比较研究[J]. 中国农业科学,2016,49(2):219-231.
[22] 王远征,王晓菁,李 源,等. 北方粳稻产量与品质性状及其相互关系分析[J]. 作物学报,2015,41(6):910-918.
[23] Xu Q,Chen W F,Xu Z J. Relationship between grain yield and quality in cultivars grown across different rice-growing areas[J]. Breeding Science,2015,65(3):226-232.
[24] 徐正进,陈温福. 中国北方粳型超级稻研究进展[J]. 中国农业科学,2016,49(2):239-250.
[25] 王嘉宇,徐正进,张世春. 水稻直立穗型研究进展与应用前景[J]. 种子,2005,24(8):58-61.
