脊髓背角细胞外信号调节激酶磷酸化参与调控2,4-二硝基氟苯诱导的小鼠慢性痒
2018-05-04吴品雯
吴品雯, 张 浩, 方 浩,3*
1. 复旦大学附属中山医院闵行分院麻醉科,上海 201199 2. 复旦大学附属肿瘤医院麻醉科,上海 200032 3. 复旦大学附属中山医院麻醉科,上海 200032
近年来痒觉分子机制研究取得巨大进展。其中,特异性表达于脊髓背根神经节(dorsal root ganglion,DRG)的Mas相关G蛋白偶联受体(Mas-related G protein coupled receptors,Mrgprs)家族蛋白被认为是调节痒觉的特异性亚群[1-4]。白细胞介素-31(interleukin-31,IL-31)及其受体参与初级神经元痒觉的信号转导[3]。脊髓背根神经节Toll样受体TRL7、TRL3和TRL4亚型表达能够增强瘙痒感觉[6-8]。神经肽类的利钠肽前体B(natriuretic peptide precursor b,NPPB)在DRG能够表达瞬时感受器受体V1(transient receptor potential vanilloid 1,TRPV1)阳性神经元,特异性调节痒觉行为;NPPB突变小鼠对痒觉行为反应选择性消失,而痛觉没有发生变化[6-8]。此外,神经递质和神经调解物(如P物质和谷氨酸)在初级感觉神经元中参与痒觉形成。
本课题组预实验结果表明,小鼠脊髓背角的细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)信号通路参与了组胺及2,4-二硝基氟苯(2,4- dinitrofluorobenzene,DNFB)诱导的痒觉,提示ERK磷酸化可能参与脊髓背角慢性痒相关的信号调节。因此,本研究以ICR(Institute of Cancer Research)小鼠为研究对象,采用过敏性皮炎模型探讨脊髓背角ERK磷酸化与慢性痒信号调节的相关性。
1 材料与方法
1.1 主要仪器及试剂 DNFB(配制成溶液以丙酮为溶剂)、丝裂原活化的细胞外信号调节激酶(mitogen-activated extracellular signal-regulated kinase,MEK)抑制剂U0126购自美国Sigma-Aldich公司。水合氯醛、异丙醇和75%乙醇溶液购自中国医药集团总公司。兔抗鼠磷酸化ERK(pERK)抗体、兔PKC-R抗体、鼠抗GFAP抗体、鼠NeuN抗体和鼠Iba1抗体,以及生物素化羊抗兔二抗和亲和素Cy3购自美国Millipore公司。TaKaRa试剂盒购自日本TaKaRa公司。
1.2 动物分组及模型制备
1.2.1 动物来源及饲养 小鼠为ICR品系,来源于上海市实验动物资源中心(西普尔-必凯公司),200只,雄性,8~12周龄。小鼠每5只1笼装,自由饮水和摄食,啮齿类动物饲料喂养,环境温度为20~25℃,相对湿度为70%,12 h光暗交替照明(7:00~19:00)。
1.2.2 慢性痒模型的建立 面颊部慢性痒模型:每次涂抹DNFB溶液前1 d备皮,选取小鼠右侧颊部,棉棒蘸取DNFB溶液,涂抹于右侧面颊。颈背部慢性痒模型:每次涂抹DNFB前1 d备皮,涂抹后立即放入笼盒中,开始录像30 min,并记录30 min内小鼠的搔抓行为及搔抓行为的潜伏期,最后统计30 min内搔抓的总数。5 min为1个单位,小鼠后肢抬起搔抓颈部皮肤至后肢撤回至地面记为1次搔抓行为。录像及搔抓行为计数均采用双盲法。
1.3 痒觉行为学观察 小鼠随机分为2组:实验组涂抹DNFB/丙酮溶液4次制备瘙痒模型,对照组涂抹丙酮溶液。4次涂抹完成(第1、4、11、18天)后,先观察最后1次涂抹后1、6、12、24 h共4个时间点的小鼠搔抓行为,并在4次涂抹DNFB或丙酮后分别在第1、3、7、14天观察小鼠是否有自发性搔抓行为。DNFB实验组小鼠分为2组:鞘内注射U0126组和鞘内注射PBS对照组,同样记录小鼠搔抓行为次数及搔抓潜伏期,均采用双盲法。鞘内注射U0126或PBS时,动作要轻盈、准确、迅速。首先要准确定位小鼠腰部髂脊(小鼠背部最突出处),注射器的针头与脊髓约成45°夹角,进针及注药时小鼠尾巴抬动,说明注射效果良好。鞘内注射后,分别在第1、3、7、14天观察小鼠搔抓行为。
1.4 免疫荧光组织化学染色
1.4.1 脊髓切片的获取 10%水合氯醛按10 μL/g的剂量进行小鼠腹腔内注射麻醉,麻醉后迅速用眼科剪剖开小鼠胸部,暴露小鼠心脏,眼科剪去除右心耳,将注射器针头插入左心室(最佳注射点为心尖处,进针不宜过深,以防穿破左心室造成灌注液进入肺部),快速推注0.01 mol/L PBS 20 mL,至清凉液体流出。推注过程中,肺部及肝脏逐渐变为白色后,缓慢推注4%多聚甲醛溶液20 mL;推注过程中出现四肢抽动、僵硬、胡须直立,说明多聚甲醛已进入小鼠大脑,待抽动停止及全身组织变硬后开始取材,仔细完好地分离出颈部脊髓。取材成功后,4%多聚甲醛溶液固定24 h,30%蔗糖脱水24 h,当组织沉底说明脱水效果好,滤纸吸去组织残余蔗糖,OCT胶包埋组织,放置-20℃。待组织完全冻结后,用刀将组织从模具中取出,固定于底座上,冰冻切片机进行颈部脊髓切片,脊髓切片厚度为30 μm,采用漂浮切片法保存。
1.4.2 单标染色 采用流动清水冲洗80孔免疫组织化学板,然后将其晾干。用抗体稀释液稀释抗体,加入兔抗鼠pERK一抗(按1∶500 稀释),室温下孵育1 h,置于4℃冰箱过夜。将所有脊髓组织切片从组化板中取出,0.01 mol/L PBS漂洗5 min 3次。加入生物素化的马抗兔二抗(1∶500 稀释),避光及室温下孵育2 h。PBS漂洗5 min 3次,加入亲和素Cy3(1∶500 稀释),室温孵育1 h。PBS漂洗7 min 3次,挑出组织切片,拖拽至玻片,75%甘油封固,4℃保存。荧光显微镜观察组织形态学并拍片保存。
1.4.3 双标染色 基本操作同单标方法,差别在于:一抗为兔抗鼠pERK(1∶500 稀释)和鼠抗GFAP(1∶1 000 稀释),或兔抗鼠pERK(1∶500 稀释)和鼠抗NeuN(1∶1 000 稀释),或兔抗鼠pERK(1∶500 稀释)和鼠抗Iba1(1∶1 000 稀释);二抗为生物素化的羊抗兔bio-HAR和DAM-488(1∶500 稀释);三抗为亲和素Cy3(1∶500 稀释)。各步骤间用0.01 mol/L PBS漂洗10 min 3次。
1.5 酪酰胺信号放大(TSA)技术染色 第1天:漂白液(3% H2O22 mL、甲醇16 mL、PBS 2 mL)漂白脊髓切片10 min;PBS漂洗3次,每次5 min;TNT漂洗3次,每次5 min;TNB封闭液封闭1 h;加入兔抗鼠pERK抗体(稀释比例1∶10 000),将脊髓切片挑回组化板内,并置于4℃冰箱过夜。第2天:TNT漂洗3次,每次5 min;加入用TNB稀释的兔抗性辣根过氧化物酶(稀释比例为1∶100),室温孵育2 h;TNT漂洗3次,每次5 min;加入TSA染色剂(1∶50稀释),孵育约20 min,具体以组织染色程度为准,随时观察,染色效果好即可终止染色;PBS漂洗3次,每次5 min;pERK染色完成后,加入兔抗鼠Iba1抗体(稀释比例1∶1 000),4℃冰箱过夜。第3天:PBS漂洗3次,每次5 min;加入二抗DAR-488(稀释比例1∶500),室温孵育2 h;PBS漂洗3次,每次5 min。

2 结 果
2.1 DNFB对小鼠行为学的影响 结果(图1)表明:在面颊部模型中,DNFB组和丙酮对照组小鼠涂抹药物30 min内的擦涂行为次数差异无统计学意义(P=0.879),而丙酮对照组小鼠搔抓行为次数少于DNFB组,差异有统计学意义(P=0.02),提示DNFB诱发的小鼠行为是痒觉。DNFB组(n=12)小鼠经4次药物涂抹,第1、4、11、18天每次涂抹后30 min内搔抓行为次数均明显多于丙酮对照组(n=5),差异有统计学意义(P<0.05,图2)。
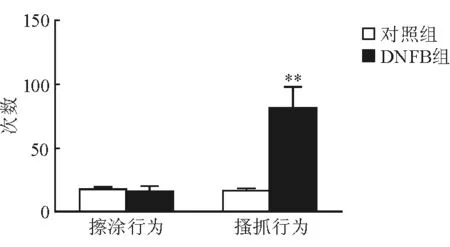
图1 面颊部模型中两组小鼠特征行为的对比
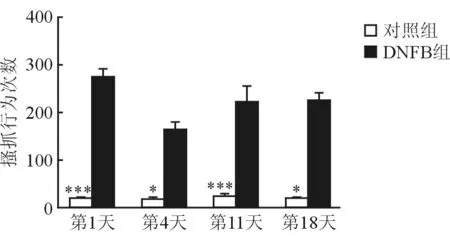
图2 两组小鼠不同时间点搔抓行为的对比
2.2 MEK抑制剂U0126对慢性痒行为的抑制作用 DNFB溶液反复刺激皮肤后,会对小鼠造成明显的皮肤病理性增生和损伤,同时会诱发小鼠自发性搔抓行为,并且这种自发性搔抓行为可持续14 d。进一步的研究结果(图3)提示:与注射PBS相比,鞘内注射U0126可显著减少小鼠的自发性搔抓行为,差异有统计学意义(P<0.001)。
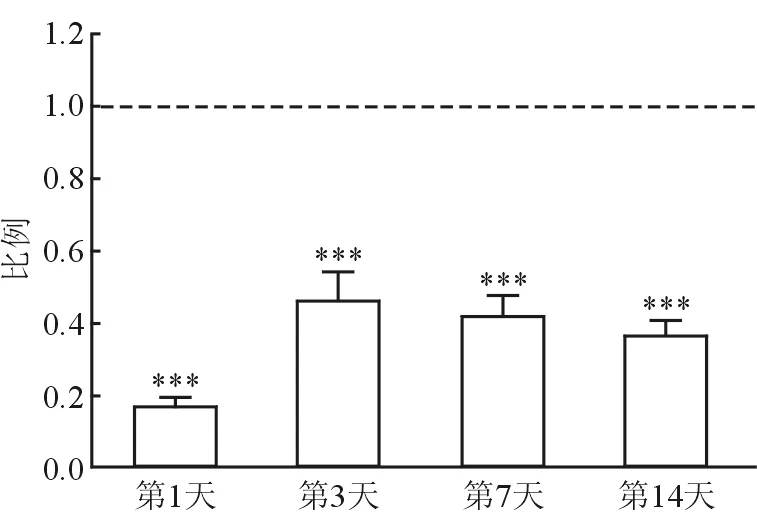
图3 MEK抑制剂U0126对自发性慢性痒行为的抑制作用
2.3 脊髓背角pERK参与DNFB诱导的慢性痒 免疫荧光组织化学染色结果(图4)显示:与PBS对照组相比,鞘内注射U0126可明显减少pERK阳性表达神经元的数量,最后1次涂抹DNFB后的第1、3 、7、14天分别减少了47%、59%、40%、50%,差异有统计学意义(P<0.001)。
2.4 慢性痒背部模型pERK的神经元定位 免疫荧光染色结果(图4)显示:pERK阳性表达细胞主要与NeuN共标,而不与GFAP、Iba1共标,提示pERK阳性表达细胞是神经元细胞,与急性痒一致。pERK阳性表达细胞与表达在Ⅱ内层的PKC-r不共标,提示pERK阳性表达神经元依然位于脊髓背角Ⅰ层,与急性痒状态基本一致。
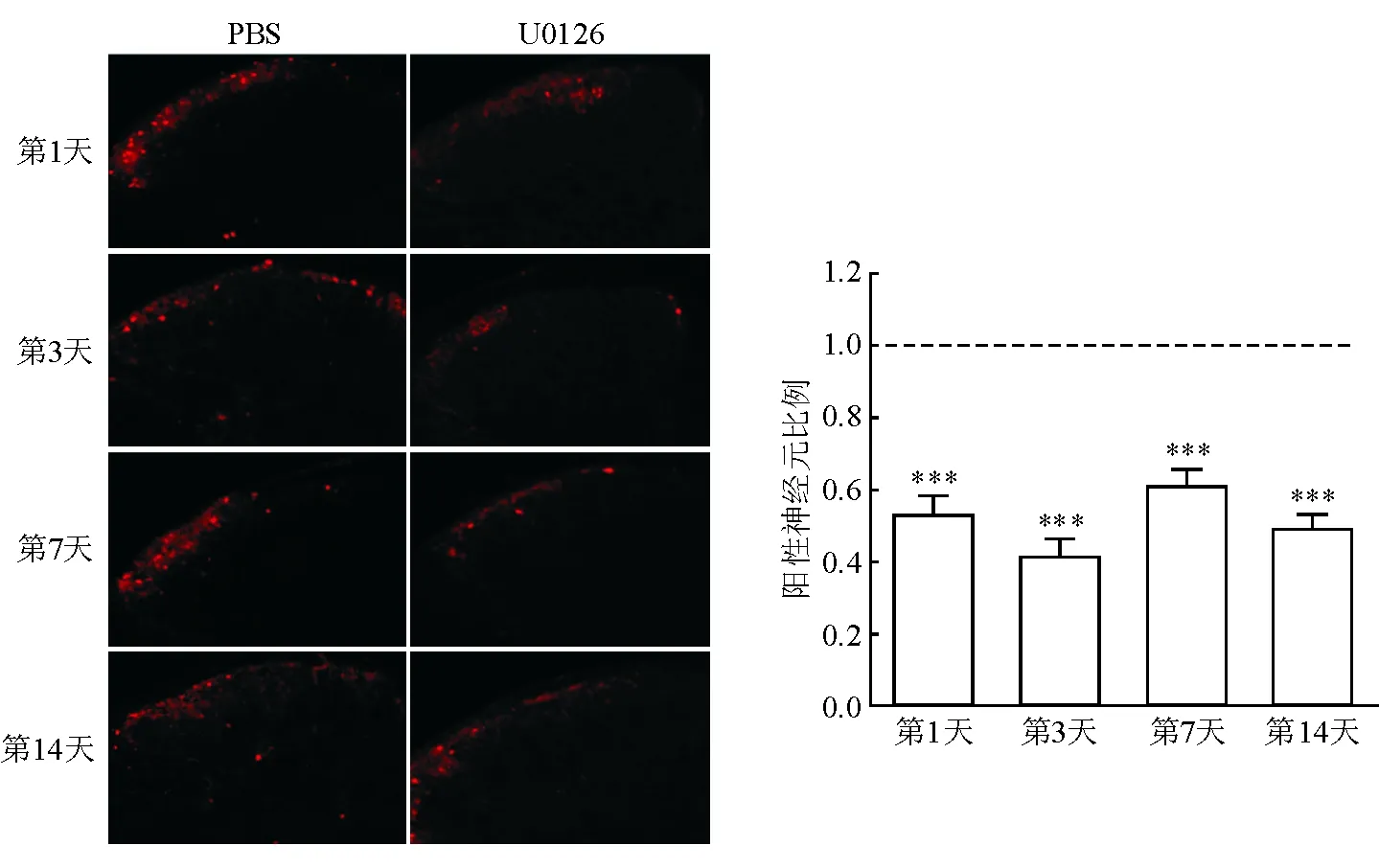
图4 两组小鼠脊髓背角pERK阳性神经元的对比
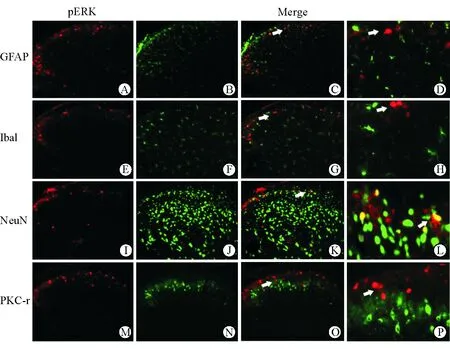
图5 慢性痒背部模型pERK的神经元定位
3 讨 论
慢性痒常与皮肤炎性疾病、系统性疾病、免疫功能紊乱等相关,炎性介质通过直接激活或敏化痒觉神经元参与其中。Inagaki等[8]尝试采用0.15% DNFB/丙酮溶液涂抹于腹部备皮区域建立小鼠过敏性皮炎并伴随频繁搔抓行为模型,以此来探讨痒觉的发生机制,取得初步成效。面颊模型可以从行为学上将痛觉和痒觉区分出来[4]。本研究面颊部模型中,DNFB组小鼠搔抓行为(30 min内86次)明显超过对照组小鼠(30 min内17次),而擦涂行为没有明显区别。结果提示DNFB/丙酮诱导的行为是痒觉,证实慢性痒模型建立成功。为制备背部瘙痒模型,本研究在小鼠颈背部区域涂抹0.15%DNFB/丙酮溶液150 μL,在每个时间点DNFB组小鼠搔抓行为次数明显高于对照组。但本研究的搔抓次数低于其他文献结果[5-6],可能与计数时间、实验小鼠品系不同有关。为进一步验证MEK抑制剂U0126能否对慢性痒发挥作用,本研究对小鼠进行鞘内注射0.1 μg/μL U0126。结果发现,DNFB组小鼠搔抓次数明显减少,DNFB组pERK阳性表达神经元数量也显著少于对照组,与行为学结果相一致。行为学指标的检测结果表明,本研究成功建立了DNFB诱导的过敏性皮炎慢性痒模型,小鼠的慢性痒可持续14 d,可用于观察和研究脊髓背角表达pERK的神经元对慢性痒的影响及其与MAPK通路的关系。
pERK表达增多是细胞外调节激酶活化磷酸化的标志,同时也是脊髓背角参与痛觉信号处理的标志。本课题预试验结果证实,ERK磷酸化也是脊髓背角参与组胺释放和DNFB诱发痛觉信号调节的标志。本研究着重探讨DNFB诱发的过敏性皮炎是否会对小鼠造成慢性痒,以及脊髓在慢性痒模型中的作用。然而DNFB诱发的慢性痒需要ERK的持续磷酸化,并且使用MEK抑制剂U0126可有效地缓解搔抓行为。因此,还需要探索慢性痒模型在DRG水平的ERK表达情况,以进一步证实ERK参与痒觉的调节。
pERK阳性表达神经元的形态学特征对研究脊髓痒觉机制有重要作用[7-10]。为了探讨DNFB诱导的慢性痒模型中pERK阳性表达的细胞类型及位置是否发生改变,本研究分别与NeuN(神经元标志物)、GFAP(星形胶质细胞标志物)和Ibal(小胶质细胞标志物)共标。结果表明,pERK阳性表达细胞是神经元,且pERK阳性表达神经元依然位于脊髓背角Ⅰ层,与急性痒一致。
综上所述,慢性痒模型中pERK阳性表达细胞仍然是神经元,脊髓背角浅层神经元ERK磷酸化参与了DNFB诱导的慢性痒的调控。
[ 1 ] IKOMA A, STEINHOFF M, STNDER S, et al. The neurobiology of itch[J]. Nat Rev Neurosci, 2006,7(7):535-547.
[ 2 ] ELMARIAH S B, LERNER E A. Topical therapies for pruritus[J]. Semin Cutan Med Surg, 2011,30(2):118-126.
[ 3 ] PATEL T, YOSIPOVITCH G. Therapy of pruritus[J].Expert Opin Pharmacother, 2010,11(10):1673-1682.
[ 4 ] GRIMSTAD O,SAWANOBORI Y,VESTERGAARD C,et al.Anti-interleukin-31-antibodies ameliorate scratching behaviour in NC/Nga mice: a model of atopic dermatitis[J]. Exp Dermatol, 2009,18(1):35-43.
[ 5 ] ZHANG L,JIANG G Y,SONG N J,et al.Extracellular signal-regulated kinase (ERK) activation is required for itch sensation in the spinal cord[J]. Mol Brain, 2014,7:25.
[ 6 ] ZHAO Z Q, HUO F Q, JEFFRY J, et al. Chronic itch development in sensory neurons requires BRAF signaling pathways[J]. J Clin Invest, 2013,123(11):4769-4780.
[ 7 ] BOURANE S, DUAN B, KOCH S C, et al. Gate control of mechanical itch by a subpopulation of spinal cord interneurons[J]. Science, 2015,350(6260):550-554.
[ 8 ] INAGAKI N, SHIRAISHI N, IGETA K, et al.Inhibition of scratching behavior associated with allergic dermatitis in mice by tacrolimus, but not by dexamethasone[J].Eur J Pharmacol,2006, 546(1-3):189-196.
[ 9 ] KREMER A E, OUDE ELFERINK R P, BEUERS U. Pathophysiology and current management of pruritus in liver disease[J]. Clin Res Hepatol Gastroenterol, 2011,35(2):89-97.
[10] FOSTER E,WILDNER H,TUDEAU L,et al.Targeted ablation, silencing, and activation establish glycinergic dorsal horn neurons as key components of a spinal gate for pain and itch[J]. Neuron, 2015,85(6):1289-1304.
