鸡血藤醇提物及其活性成分儿茶素对辐射损伤小鼠脾脏JAK/STAT通路的影响
2018-05-03董宪喆王琪珊郭代红解放军总医院临床药理研究室北京00853重庆医科大学
董宪喆,王琪珊,牛 可,谭 潇,郭代红,刘 屏(.解放军总医院临床药理研究室,北京 00853;.重庆医科大学,
重庆 400016)
随着人们生活水平的提高,手机、电脑、核能和核技术等低剂量辐射的危害日益严重。同时,临床大部分恶性肿瘤患者放疗过程中长期低剂量辐射会造成骨髓抑制、免疫器官功能减退,造血功能障碍,也能直接杀伤外周血细胞,这些副作用成为限制恶性肿瘤长期放射治疗的主要因素[1]。
目前抗辐射药物主要有激素类、天然药物、硫醇类及其衍生物、细胞因子类等[2]。鸡血藤为豆科植物密花豆(Spatholobus suberectus Dunn)的干燥藤茎。《纲目拾遗》等记载鸡血藤为“血分之圣药”,“去淤血,生新血”,具有活血补血,调经止痛,舒筋活络。用于月经不调,血虚萎黄,风湿痹痛,痛经,经闭,麻木瘫痪等[3]。现代药理研究表明鸡血藤具有抗放射线、降血压、抑菌、抗癌的功效,近年来临床上常用于治疗贫血,各种原因引起红细胞、白细胞、血小板等全血象减少等疾病,具有很好的疗效。基于此,本实验采用亚急性辐射损伤小鼠模型,以小鼠脾脏为主要研究对象,更深层次的研究鸡血藤醇提物及儿茶素抗辐射作用及其分子机制。
1 实验材料
1.1 动物
SPF级ICR小鼠(6 ~ 8周,18 ~ 22 g),购于北京维通利华实验动物技术有限公司,雌雄各半,动物许可证号:SCXK(京)2012-0001。在动物适应环境1周后正式进行实验。
1.2 主要仪器
60Coγ钴源(军事医学科学院); R-200型旋转蒸发仪(瑞士BUCHI公司);ABJ80-4NM电子天平(德国KERN公司);电泳仪、半干槽(美国Bio-Rad公司);EC3 Imaging Systems凝胶成像系统(美国UVP公司);3K15冷冻高速离心机(德国Sigma公司),MX-S旋涡混合器(美国SCILOGEX公司);KS4000i控制型空气浴恒温摇床(德国IKA公司);BIO-Photometer型核酸蛋白测定仪(德国Eppendorf公司);MyCycler、iCycler系列荧光定量PCR仪(美国Bio-Rad公司);1420-012 Wallac Victor3 多标记酶标仪(Perkin Elmer公司);金属浴(金银杏生物科技北京有限公司)。
1.3 主要试剂
鸡血藤中药饮片(北京绿野药业有限公司,产地:福建);儿茶素(德国Sigma公司);注射用氨磷汀(大连美罗药厂,0.4 g/支);JAK2、STAT5a、P-STAT5a、STAT5b、P-STAT5b、P-JAK2一抗,β-actin,鼠IgG2b同型对照和羊抗鼠IgG二抗(美国Abcam公司);BCA蛋白浓度测定试剂盒(碧云天生物技术研究所);RIPA裂解液(普利莱北京基因技术有限公司);蛋白磷酸酶抑制剂混合物(普利莱北京基因技术有限公司);超敏发光液(普利莱北京基因技术有限公司);DEPC处理水(北京索莱宝科技有限公司);SsoFast™EvaGreen®Supermix试剂盒(200×20 µL reactions,货号1725200,美国Bio-RAD公司);iScript cDNA Synthesis kit反转录试剂盒(美国Bio-RAD公司)。
1.4 鸡血藤醇提物制备
鸡血藤中药饮片剪成小块,8倍量体积75%乙醇浸泡过夜后回流提取3次,每次1 h,合并滤液,减压旋蒸至浸膏,冻干,得鸡血藤乙醇提取物粉末。
2 实验方法
2.1 分组及给药
把SPF级ICR小鼠随机分为正常组(CON组),模型组(MODEL组),阳性对照组(P-CON组,氨磷汀,43.6 mg·kg-1,照射前30 min尾静脉注射给药),鸡血藤醇提物低、中、高剂量组(JXT-L组、JXT-M组、JXT-H组,相当于生药10、20、40 g·kg-1体重),儿茶素低、高剂量组(EGCG-L组、EGCG-H组,50、100 mg·kg-1体重),每组18只。除正常组外,各组均接受6.0 Gy剂量60Coγ射线一次性全身照射(剂量率117.29 cGy·min-1,时间290 s)。自照射当日起,每只动物灌胃给予相应药物0.2 mL,正常组、模型组给予等量生理盐水,每日1次,连续给药28 d。
2.2 外周血象的测定
各组于照射前及给药21 d和28 d后剪尾尖采血20 μL,BC-2800Vet兽用全自动血液细胞分析仪测定外周血白细胞(WBC),红细胞(RBC),血小板(PLT)计数水平,观察给药后小鼠外周血象的变化。
2.3 Western blotting法测定小鼠脾脏相关蛋白的表达
采用Western blotting法测定小鼠脾脏中β-actin、JAK2、P-JAK2、STAT5a、P-STAT5a、STAT5b、P-STAT5b等蛋白的表达。用分析天平称量小鼠脾脏的重量(确定加入裂解液的量),在冰上用剪刀将组织剪成小于1 mm3的小块,按照每100 mg组织加入1 mL裂解液的比例加入裂解液。在玻璃匀浆器中研磨至95%破碎,转移至2 mL EP管中,冰上放置10 min,每隔2 min放在旋涡混合仪上振荡10 s。充分裂解后,4 ℃ 12 000 g离心10 min,上清转移至新EP管中,测定蛋白浓度后放在100 ℃金属浴中煮沸5 min,冷却后放入– 20 ℃保存(一个月内短期使用)。制好胶后,将样品加入浓缩胶中进行电泳(电压为55 V),当蛋白跑至浓缩胶与分离胶分界处时,调整电压至135 V,跑至分离胶底端电泳结束。电泳完毕后,取下胶,切除浓缩胶和多余分离胶,剪取大小合适的PVDF膜放入甲醇中活化1 min,然后300 mA低温转膜90 min。转膜完毕后,在5% BSA溶液中室温封闭3 h。然后按说明书推荐比例加入抗体,放在摇床上室温孵育1 h,4℃孵育过夜。TBST洗膜3次,每次10 min,加入相应二抗室温孵育3 h。TBST洗膜3次,每次10 min。加上适量显色液,室温反应2 min后将膜移至UVP凝胶成像仪中显影,并分析各条带光密度值。
2.4 小鼠脾脏中相关蛋白基因定量PCR实验
引物设计:设计并合成β-actin、JAK2、STAT5a、STAT5b、Bcl-2、Caspase-3、Bax的上下游引物,引物信息详见表1。

表1 各基因引物序列信息Tab 1 Information of primer sequences of each gene
总RNA提取:把小鼠脾脏组织置于液氮中研磨至白色粉末状,加入1 mL Trizol溶液继续研磨至液态,用加样枪取出至EP管中。室温放置5 min,使其充分裂解。加入200 μL氯仿,振荡混匀后,室温放置15 min后,4 ℃ 12 000 r·min-1离心15 min,取上层水相至新离心管中,按每毫升Trizol加入0.5 mL异丙醇,加入0.5 mL异丙醇混匀,室温放置5 ~ 10 min。4 ℃12 000 r·min-1离心10 min,弃上清。加入1 mL 75%乙醇,温和振荡离心管,悬浮RNA沉淀,4 ℃ 8000 r·min-1离心5 min,尽量去上清,室温晾干5 ~ 10 min。将上述RNA样品用30 μL的无RNA酶水溶解后用核酸蛋白浓度测定仪测定RNA浓度。
逆转录反应:逆转录采用iScriptTMcDNA Synthesis kit的方法。取200 μL的PCR管,冰上依次加入5×iScript reaction mix(4 μL)、逆转录酶(1 μL)、无核酸酶水(14 μL)和RNA模板(1 μg total RNA,1 μL),总反应体系为20 μL;反应条件为:25 ℃,5 min;42 ℃,30 min;85 ℃,5 min。
qPCR反应:qPCR扩增使用SsoFast™ EvaGreen®Supermix的方法,在Bio-Rad IQ-5实时定量PCR仪中进行。取200 μL的PCR管,依次加入SYBGreen super mix(10 μL)、上下游引物(各1 μL)、无核酸酶水(6 μL)和cDNA模板(2 μL),总反应体系为20 μL;反应条件为:预变性:95 ℃,30 s;变性:95 ℃,5 s;退火/延伸:55 ~ 60 ℃,10 s;反应40个循环。
2.5 统计学分析
实验结果用均数±标准差来表示,实验数据用SPSS 17.0统计软件进行单因素方差分析,组间比较采用Dunnett-t检验。
3 实验结果
3.1 对模型小鼠外周血象的影响
与正常组比,模型组小鼠WBC,RBC,PLT水平均有所下降。与模型组相比,鸡血藤醇提物中、高剂量组给药28 d可显著升高小鼠WBC和RBC水平,高剂量组给药21 d可显著升高PLT水平;儿茶素高剂量组给药28 d可显著升高小鼠WBC水平,给药21 d可显著升高PLT水平,详见图1。
3.2 对模型小鼠脾脏中凋亡相关基因表达的影响
RT-PCR结果显示,给药28 d后,与正常组相比,模型组脾脏组织中Bax和Caspase的表达显著上调(P <0.01),Bcl-2的表达显著下调(P < 0.01);鸡血藤醇提物各剂量组与模型组相比,均可显著下调Bax和Caspase的mRNA的表达(P < 0.05),上调Bcl-2的表达(P < 0.01),且都显著低于模型组(P < 0.01)。儿茶素各剂量组与模型组相比,均可显著下调Bax和Caspase的mRNA的表达(P < 0.01),上调Bcl-2的表达(P < 0.01)。鸡血藤醇提物组下调基因表达作用更明显,详见图2。
3.3 对模型小鼠脾脏中JAK/STAT通路相关基因表达的影响
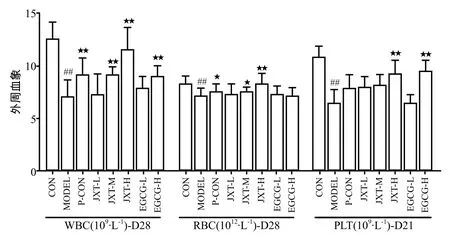
图1 各组药物对给药后小鼠外周血象的影响Fig 1 Effects of each medication on peripheral blood cells level in mice after administration

图2 各组药物对给药后28 d小鼠脾脏中Bax,Bcl-2,Casepase-3基因表达的影响Fig 2 Effects of each medication on the expressions of Bax,Bcl-2 and Casepase-3 in spleen cells of mice on 28th day after administration
与正常组相比,模型组脾脏组织中JAK/PJAK、STAT5a/PSTAT5a和STAT5b/PSTAT5b的基因表达情况显著下调(P < 0.01);鸡血藤醇提物各剂量组与模型组相比,均可显著上调JAK/STAT信号通路中mRNA的表达(P < 0.01)。儿茶素各剂量组与模型组相比,均可显著上调JAK/STAT信号通路中mRNA的表达(P < 0.01),与鸡血藤醇提物组相比,儿茶素组上调作用更显著。详见图3。
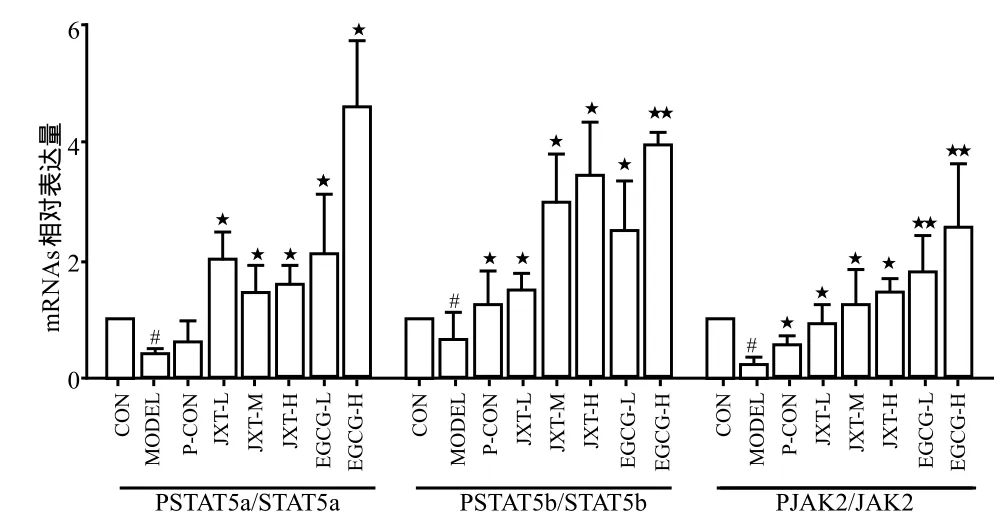
图3 各组药物对给药后28天小鼠脾脏中PSTAT5a/STAT5a,PSTAT5b/STAT5b,PJAK2/JAK2基因表达的影响Fig 3 Effects of each medication on the expressions of PSTAT5a/STAT5a,PSTAT5b/STAT5b and PJAK2/JAK2 pathways in spleen cells of mice on 28th day after administration
3.4 对模型小鼠脾脏JAK-STAT通路关键蛋白的影响
与正常组相比,在蛋白磷酸化水平上分析,模型组小鼠脾脏中STAT5a、STAT5b和JAK2的磷酸化比例在给药后均有所下降(P < 0.01),其中STAT5b下降最为显著;在总蛋白水平上分析,模型组小鼠脾脏中STAT5a和STAT5b总蛋白比例在给药后均有所下降(P < 0.01)。
鸡血藤醇提物各剂量组与模型组相比,在蛋白磷酸化水平上分析,给药后鸡血藤醇提物低剂量组和中剂量组的STAT5a磷酸化比例都显著升高(P < 0.01),鸡血藤醇提物低剂量组的JAK2磷酸化比例也有显著升高(P < 0.01),且都显著高于模型组(P < 0.05);在总蛋白水平上分析,鸡血藤醇提物中剂量组STAT5b总蛋白比例显著升高(P < 0.01),此外鸡血藤醇提物低、中剂量组JAK2总蛋白比例也显著升高(P < 0.01)。
儿茶素各剂量组与模型组相比,在蛋白磷酸化水平上分析,给药后儿茶素高剂量组JAK2的磷酸化比例显著增高(P < 0.01),且显著高于模型组(P < 0.05);在总蛋白水平上分析,儿茶素低剂量组STAT5b总蛋白比例显著升高(P < 0.01)。详见图4 ~ 6。
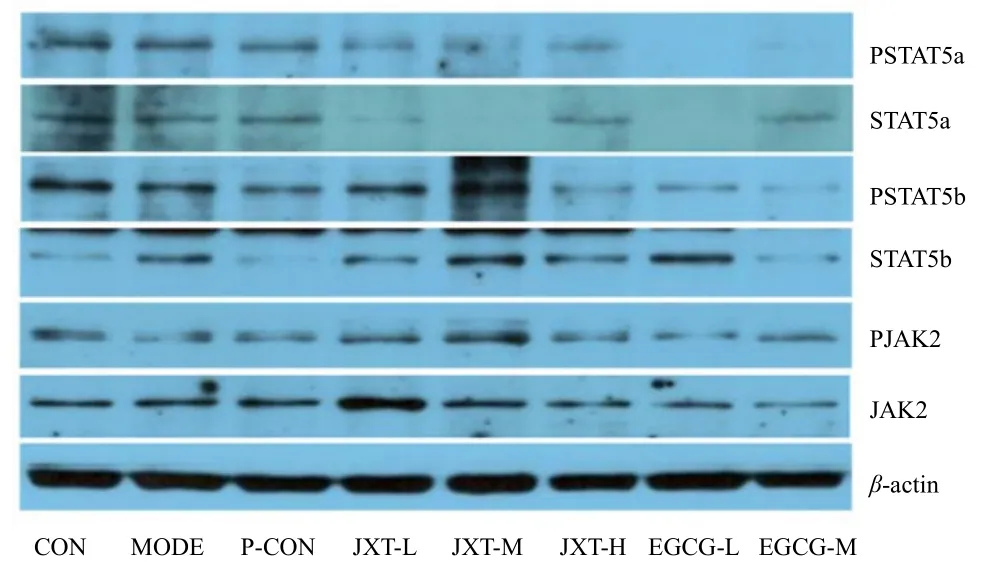
图4 Western Blotting法检测小鼠脾脏相关蛋白的表达Fig 4 The expression of related protein in spleen cells of mice using Western-Blotting
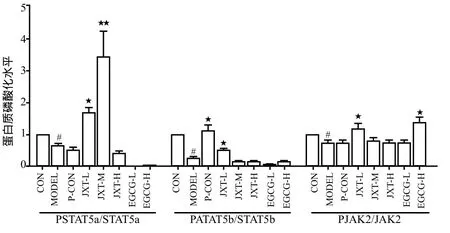
图5 各组药物对给药28 d后小鼠脾脏中STAT5a, STAT5b, JAK2磷酸化的影响Fig 5 Effects of each medication on the phosphorylation of STAT5a, STAT5b and JAK2 pathways in spleen cells of mice on 28th day after administration
4 讨论
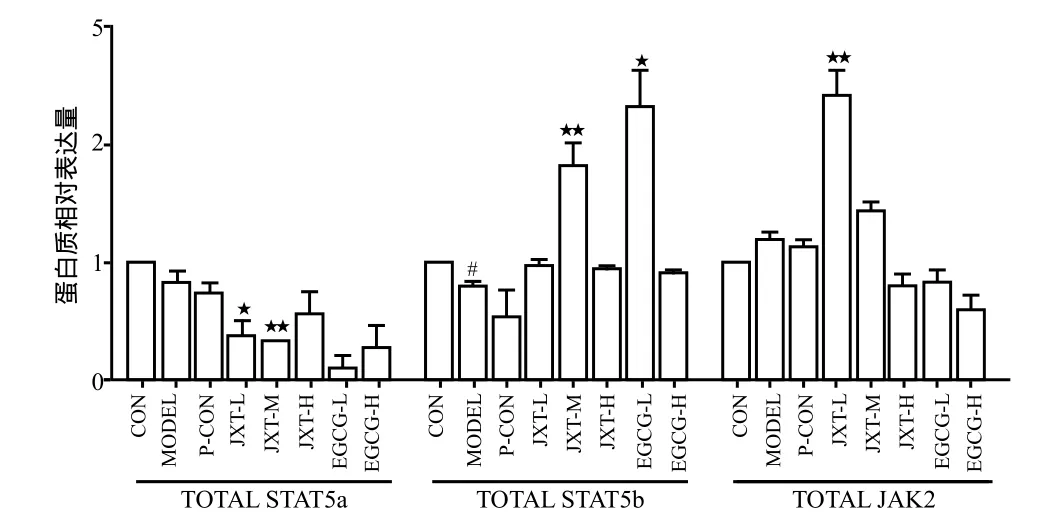
图6 各组药物对给药28天后小鼠脾脏中STAT5a, STAT5b, JAK2总蛋白量的影响Fig 6 Effects of each medication on the amount of total protein of STAT5a,STAT5b and JAK2 pathways in spleen cells of mice on 28th day after administration
辐射可造成机体多系统出现损伤,主要损害DNA、膜蛋白等生物大分子,从而使机体中细胞、免疫、血液、消化、生殖等系统发生损伤,甚至会诱发引发细胞的癌变、突变等[4]。由于造血系统对辐射高度敏感、更新活跃、增殖旺盛,因此常作为辐射最先损伤的系统[5]。临床常表现为造血功能障碍,外周血象降低。免疫系统对辐射也极为敏感,受辐射后小鼠的脾脏、胸腺等免疫器官及淋巴细胞、外周血白细胞等均受到严重损伤,基因突变、DNA链断裂和染色体变异,以及大量高活性和化学性质不稳定的自由基(ROS),诱发脂质过氧化,DNA和自由基相互作用造成组织器官损伤[6-7],从而导致机体免疫功能降低。目前抗辐射药物主要包括激素类、硫化物等[8-9],但明显的毒副作用限制了这些药物的应用。
鸡血藤是补血通脉之要药,本课题组前期研究已经发现并鉴定了鸡血藤抗辐射促造血的活性部位和4种以茶多酚类化合物为代表的主要活性成分:表儿茶素(epicatechin),儿茶素(catechin),表没食子儿茶素没食子酸酯(epigallocatechin,EGCG),没食子儿茶素(gallocatechin)等[10]。以儿茶素含量最多且刺激增殖活性相对较强,对各系造血祖细胞有较明显刺激作用,也是鸡血藤活血补血的最主要物质基础,其抗辐射作用也多有研究报道[11]。
本文通过研究发现,鸡血藤醇提物(儿茶素含量为17.2%)及其活性成分儿茶素可显著改善小鼠脾脏指数,恢复辐射导致的小鼠外周血象异常。脾脏作为外周免疫器官, 是体内最大的淋巴器官,兼具免疫和造血功能[12-13]。脾脏对辐射敏感性较高,辐射损伤后脾脏体积缩小或消失,细胞核发生收缩和裂解,形成核质空泡甚至导致脾脏细胞坏死[14]。鸡血藤醇提物及儿茶素可显著升高辐射导致的小鼠脾脏指数降低,说明鸡血藤醇提物和儿茶素对辐射损伤的脾脏具有保护作用,进而调节辐射引起的造血功能障碍和免疫系统失衡。在升高脾脏指数的同时,我们还发现鸡血藤醇提物中、高剂量和儿茶素高剂量给药28 d时,可显著升高小鼠外周WBC和RBC水平,给药21 d时可显著升高血小板水平,但在28 d时各组血小板数量无显著性差异,这可能和血小板寿命和再生周期有关。
深入的机制研究发现,鸡血藤醇提物和儿茶素均可显著上调 JAK/STAT信号通路中mRNA的表达,与模型组相比,儿茶素组上调作用更显著。JAK/STAT途径是机体内普遍存在的信号通路之一,参与机体细胞增殖、分化、存活、凋亡,介导机体免疫失调和肿瘤的形成等过程[15-16]。JAK/STAT通路是造血生长因子信号传导的重要通路,鸡血藤醇提物和儿茶素通过提高JAK2,STAT5a和STAT5b蛋白的磷酸化水平,激活造血因子受体JAK2-STAT5信号通路,促进外周血象的恢复,其作用机理可能与上调JAK/STAT信号通路中相关蛋白磷酸化水平,激活JAK/STAT信号通路有关。同时,鸡血藤醇提物和儿茶素可显著下调Cleaved Caspase-3和Bax的基因表达,上调Bcl-2的基因表达,与模型组比,鸡血藤醇提物组下调基因表达作用更明显。细胞凋亡涉及一系列的基因激活,表达及调控等作用,如Bcl-2家族,Caspase家族等[17-19]。脾脏中Cleaved Caspase-3和Bax的基因表达上调,Bcl-2的表达显著下调,促进小鼠脾脏的凋亡。提示鸡血藤醇提物和儿茶素均能不同程度的抑制辐射引起的小鼠脾脏细胞的凋亡,其抗凋亡作用的产生与下调脾脏细胞中Caspase-3和Bax,上调Bcl-2的表达有关。激活JAK/STAT途径,可使Bcl-2基因表达增加,进而使Cleaved Caspase-3活性降低,进而抑制细胞凋亡[20-21]。
因此,鸡血藤醇提物和儿茶素可能通过对小鼠脾脏中JAK/STAT信号通路的激活作用,抑制脾脏细胞的凋亡,对辐射损伤的小鼠脾脏具有一定修复能力,其修复能力与上调JAK/STAT信号通路中mRNA的表达和相关蛋白磷酸化的水平有关。
[参考文献]
[1] Johnke RM, Sattler JA, Allison RR. Radioprotective agents for radiation therapy: future trends[J]. Future Oncol, 2014, 10(15):2345-2357.
[2] Hosseinimehr SJ. Trends in the development of radioprotective agents[J]. Drug Discov Today, 2007, 12(19-20): 794-805.
[3] 国家药典委员会.中华人民共和国药典(一部)[S].北京:人民卫生出版社,2010:180.
[4] Wischermann K, Popp S, Moshir S, et al. UVA radiation causes DNA strand breaks, chromosomal aberrations and tumorigenic transformation in HaCaT skin keratinocytes [J]. Oncogene, 2008,27(31): 4269-4280.
[5] 刘屏,王东晓,陈桂芸,等.鸡血藤单体化合物对造血祖细胞增殖的调控作用研究[J].中国药理学通报,2007,23(6):741-745.
[6] Moulder JE, Cohen EP. Future strategies for mitigation and treatment of chronic radiation-induced normal tissue injury[J].Semin Radiat Oncol, 2007, 17(2): 141-148.
综上所述,针对于现代柔性制造产业,如航空航天制造业,其所制造的工业零件具有品类多、批量小以及精度要求高等特点,传统的生产制造方式不能够全面满足这些生产需求特点,导致航天航空企业发展的成本较大、产品的质量和响应速度受到影响。因而实现工业机器人的高精度定位方法的优化发展,实现高精度补偿技术,提高柔性制造水平,能够为对航空航天制造领域在内的柔性制造行业提供新的动能,也可以预见,在现代柔性制造行业发展的带动下,工业机器人的精准度较为得到较大程度的提升,而工业机器人将会得到较大的发展空间。
[7] Smina TP, Joseph J, Janardhanan KK. Ganoderma lucidum total triterpenes prevent gamma-radiation induced oxidative stress in Swiss albino mice in vivo[J]. Redox Rep, 2016, 21(6): 254-261.
[8] 易湛苗,张照辉,翟所迪,等.辐射防护与辐射治疗剂临床应用及进展[J].中国药物应用与监测,2009,6(4):239-242.
[9] 李敏,赵德华,王继生.抗辐射药物研究进展[J].实用药物与临床,2017,20(12):1435-1440.
[10] 崔艳君,刘屏,陈若芸.鸡血藤有效成分研究[J].中国中药杂志,2005,30(2):121-123.
[11] 王舟,曾令福,肖元梅,等.绿茶抗辐射损伤作用研究[J].四川大学学报,2003,34(2):303-305.
[12] Wilson PF. Magnification of inter-individual variation in biological responses after low doses and dose-rates of ionizing radiation[J]. Health Phys, 2016, 110(3): 296-298.
[14] 贺许良,卢先州,陈枚,等.免疫器官辐射损伤与防护的研究进展[J].现代生物医学进展,2014,14(5):997-1000.
[15] 张青,张琳琪.JAK/STAT信号通路在细胞凋亡中的研究进展及中药干预研究[J].中医临床研究,2016,8(26):142-145.
[16] 宋丹,张立艳,董海兵,等.JAK-STAT信号通路研究进展[J].中国畜牧兽医,2012,39(6):128-132.
[17] Thulasitha WS, Umasuthan N, Jayasooriya RG,et al. A thioredox in domain-containing protein 12 from black rockfish Sebastes schlegelii: responses to immune challenges and protection from apoptosis, against oxidative stress[J]. Comp Biochem Physiol C Toxicol Pharmacol, 2016, 185-186: 29-37.
[18] Udayakumar TS, Stoyanova R, Shareef MM,et al. Edelfosine promotes apoptosis in androgen-deprived prostate tumors by increasing ATF3 and inhibiting androgen receptor activity[J]. Mol Cancer Ther, 2016, 15(6): 1353-1363.
[19] Himuro T, Horimoto Y, Arakawa A, et al. Activated caspase 3 expression in remnant disease after neoadjuvant chemotherapy may predict outcomes of breast cancer patients[J]. Ann Surg Oncol, 2016, 23(7): 2235-2241.
[20] Schindle C, Plumlee C. Inteferonspen the JAK-STAT pathway[J].Semin Cell Dev Biol, 2008, 19(4): 311-318.
[21] Guerriero ML, Dudka A, Underhill-Day N, et al. Narrative-based computational modeling of the Gp130/JAK/STAT signalling pathway[J]. BMC Syst Biol, 2009, 3: 40.
