杂种落叶松家系生长性状变异分析与优良家系选择
2018-04-25贾庆彬杨伟财张含国
李 祺 贾庆彬 杨伟财 张含国
(1. 林口县青山国家落叶松良种基地,黑龙江 牡丹江 157000;2. 吉林省林业科学研究院,吉林 长春 130033;3. 东北林业大学林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040)
落叶松(Larixgmelini)是我国北方地区重要的造林树种之一,相较于其他松科树种,其木材坚实耐腐,具有生长迅速、成林较快的特点[1-3]。目前对于落叶松研究已取得较丰富的科研成果,孙晓梅等[4]对12年生日本落叶松自由授粉家系生长性状进行遗传变异研究时发现,不同家系间胸径、树高、材积、主枝长、主枝粗、冠幅等变异达到显著或极显著水平,且各性状受中度或较强的遗传控制,具有很大的家系选择潜力。杜超群等[5]在对鄂西亚高山区日本落叶松无性系生长性状变异与早期选择进行研究时发现,11年生日本落叶松无性系间胸径、树高和材积变异系数的分布范围分别为8.17%~29.53%、5.13%~23.08%、17.30%~60.42%;不同年龄无性系间胸径、树高和材积生长量均存在极显著差异。罗旭等[6]对16年生杂种落叶松家系生长性状研究中发现,家系间与家系内都存在着丰富的变异,家系间存在极显著差异,高生长量最大的家系与最小的家系相差1.3倍,胸径相差1.4倍,材积相差达到2.3倍。高扬等[7]在落叶松不同杂交组合 (兴 × 日、日 × 长) 优良家系选择的研究中发现,落叶松杂种优势明显,树高、胸径等生长性状指标可以高出当地落叶松生产对照达到62.13%与110.51%。
本研究以12年生杂种落叶松家系试验林为研究对象,利用生长性状数据,对各参试家系进行遗传变异分析、家系间差异性比较,结合材积性状家系育种值计算结果,筛选出杂种落叶松优良家系,为今后落叶松种质资源保护、杂种落叶松良种的推广与应用提供参考。
1 材料与方法
1.1 研究对象概况
研究对象为青山林场12年生杂种落叶松家系试验林。青山林场位于黑龙江省牡丹江市林口县 (45°24′45.49″ N,130°32′55.10″ E),地处张广才岭与完达山脉交汇处,平均海拔405.70 m,年平均气温3.2 ℃,年平均降水量650 mm。试验林造林种子来源于林口县青山林场杂种落叶松种子园,共21个试验处理。其中,杂种落叶松家系15个,落叶松优良种源对照3个 (白刀山和小北湖为长白落叶松种源,乌伊岭为兴安落叶松种源),长白落叶松自由授粉优良家系对照2个 (长73~18与长73~4),杂种落叶松种子园混合种子对照1个。2005年春季播种育苗,2007年春季按完全随机区组设计营建试验林,株行距2.0 m × 1.5 m,设4个区组,每个区组内每个家系40株。
1.2 调查方法
2016年10月,林木停止生长后,对试验林4个区组内的全部个体进行树高、胸径、冠幅测量,采用形数法计算材积,按公式 (1)~(4) 分别计算材积、变异系数、遗传力、遗传增益。
V=fε(H+3)g1.3[8]
(1)
式中:V为材积 (m3);fε为落叶松实验形数 (0.41);H为树高 (m);g1.3为胸高断面积 (m2)。
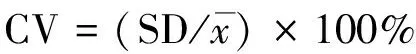
(2)
遗传力:h2=(Mf-Mfb)/Mf
(3)
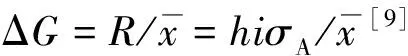
(4)
在数量遗传学中,把决定数量性状的基因加性效应值定义为育种值,在林木繁育过程中,基因加性效应可以相对稳定的遗传给子代,因此,计算育种值在林木遗传育种研究中具有重要意义[10-11]。虽然育种值不能直接度量,但可以通过数学方法进行估计,本研究采用最佳线性无偏估计 (BLUP) 法对杂种落叶松材积性状各家系育种值进行估算,BLUP法的线性混合模型一般公式为:
y=Xβ+Zu+e
(5)
式中:y为材积测量值向量;β为区组效应;u为家系遗传效应;e为随机误差效应;X和Z分别为区组效应β和家系效应u的关联矩阵。
1.3 数据分析
方差分析、多重比较利用PASW Statistics 18软件中一般线性模型进行计算,相关分析利用PASW Statistics 18软件中双变量Pearson相关分析法进行计算,家系材积性状育种值利用R软件中ASReml 3.0数据包进行计算。
2 结果与分析
2.1 杂种落叶松家系生长性状变异分析
杂种落叶松家系生长性状变异分析结果见表1。由表1可知,参试家系生长性状均存在较丰富的变异,各性状家系平均变异系数分别为树高21.23%、胸径28.40%、材积58.87%、冠幅24.7%,这为杂种落叶松家系生长性状的人工选择与遗传改良提供了较好的基础。树高方面,兴10 × 日13、兴7 × 日77-2、日3 × 兴9这3个家系存在较大的变异,其中变异最大的兴10 × 日13家系高出参试家系平均值55.68%,高出混种对照99.70%,高出变异系数最低的日12 × 兴9家系148.12%。胸径方面,日3 × 兴9、日5 × 兴12、日5 × 长78-3这3个家系存在较大的变异,变异最大的日3 × 兴9家系高出参试家系平均值26.44%,高出混种对照38.60%,高出变异系数最低的日12 × 兴9家系66.10%。材积方面,日3 × 兴9、日5 × 兴12、兴7 × 日77-2这3个家系存在较大的变异,3个家系平均变异系数为65.86%,高出参试家系平均变异系数11.87%,高出变异系数最低的兴5 × 兴9家系50.16%。其中,变异最大的日3 × 兴9家系高出参试家系平均值13.71%,高出混种对照26.25%,高出种源对照白刀山、小北湖、乌伊岭分别为14.45%、2.20%和20.40%,高出长白落叶松自由授粉家系对照长73-18与长73-4分别为13.40%、14.29%,高出变异系数最低的兴5 × 兴9家系52.62%。综合各家系生长性状变异结果,发现日3 × 兴9家系在所有参试家系中表现出较丰富的变异,日12 × 兴9家系变异相对较小。
2.2 杂种落叶松家系间生长性状差异比较
杂种落叶松家系各性状方差分析结果见表2。
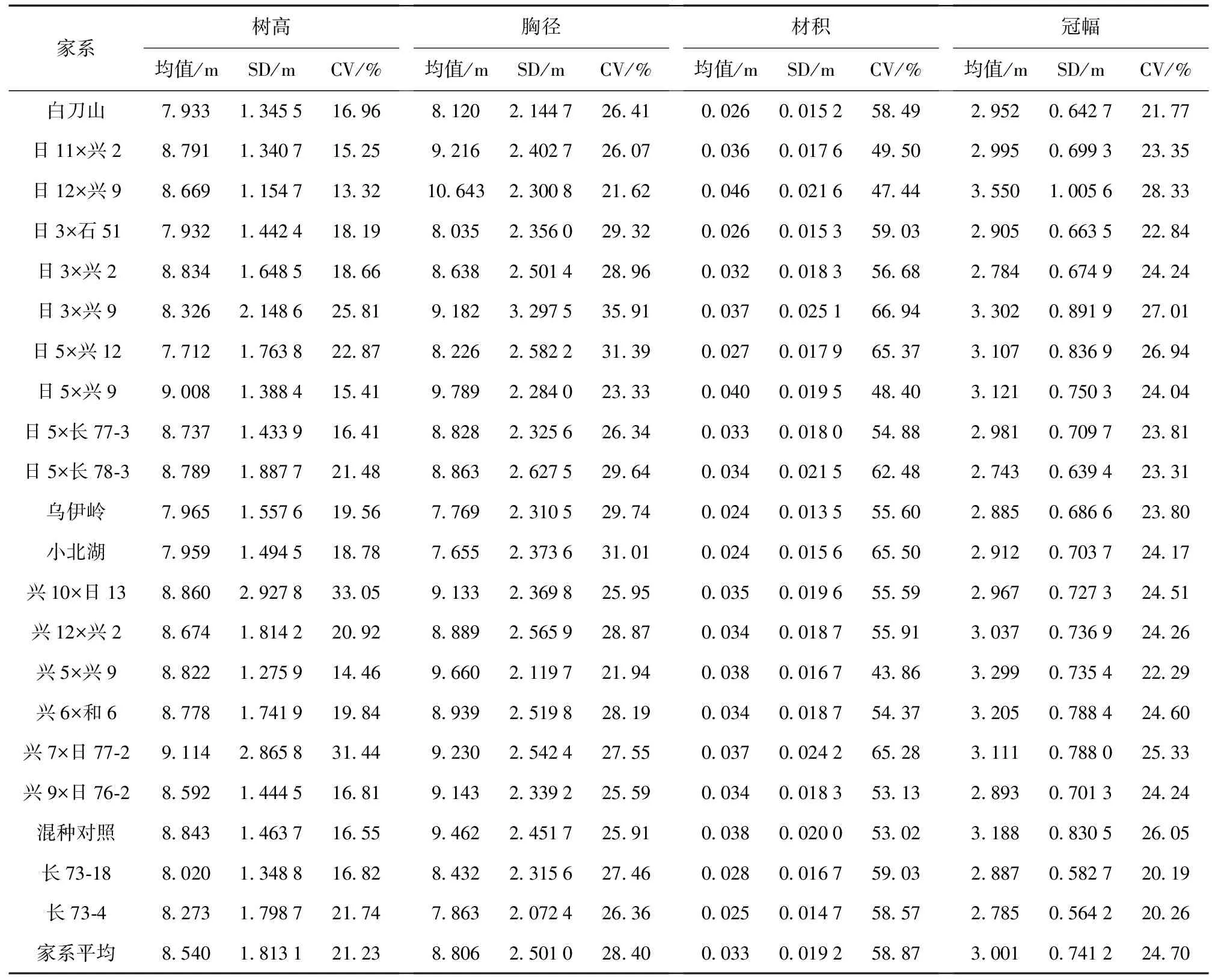
表1 杂种落叶松家系生长性状变异分析Table 1 Variation analysis of growth traits of hybrid L.gmelini families
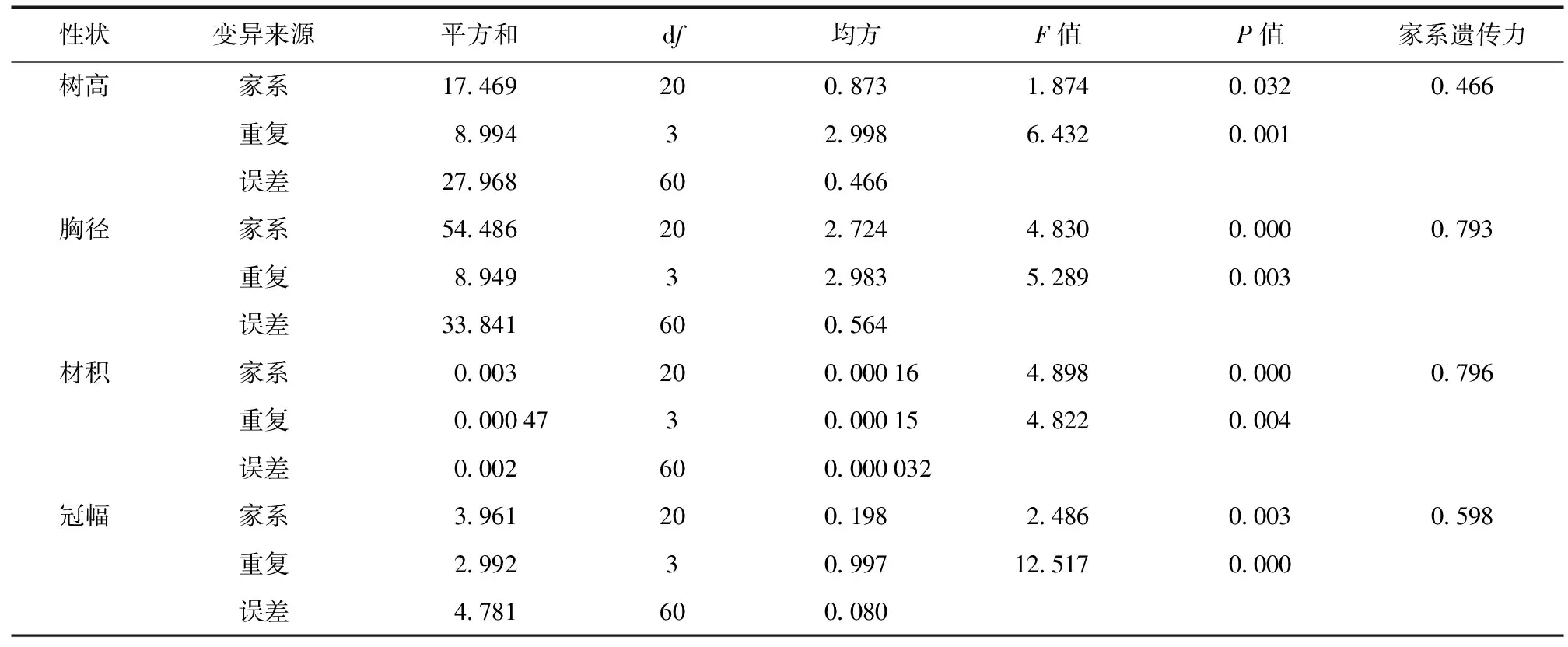
表2 杂种落叶松家系各性状方差分析Table 2 Variance analysis of growth traits of hybrid L.gmelini families
由表2可知,家系间树高性状差异达到显著水平 (P< 0.05),胸径、材积量、冠幅性状差异均达到极显著水平 (P< 0.01),各性状家系遗传力分别为树高0.466、胸径0.793、材积0.796、冠幅0.598。其中,胸径与材积家系遗传力较高,受较强遗传控制,树高与冠幅受中等强度遗传控制,该结果表明,杂种落叶松生长性状表现可以较好的遗传给子代,为今后优良家系的选育与改良提供了较好的基础。
杂种落叶松家系间生长性状多重比较结果见表3。
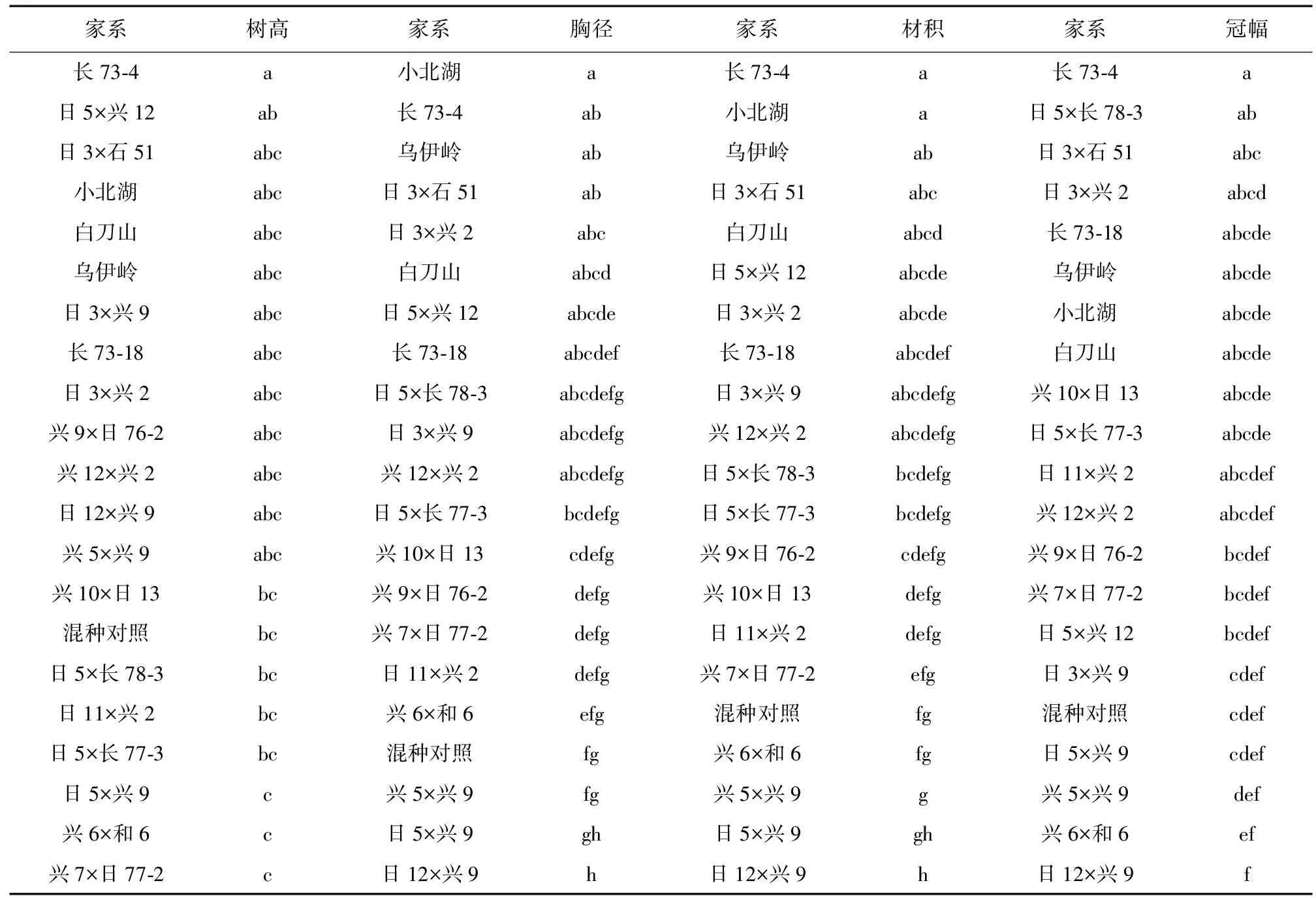
表3 杂种落叶松家系间生长性状多重比较Table 3 Multiple comparisons of growth traits among hybrid L.gmelini families
注:不同小写字母表示差异显著。
由表3可知,日12 × 兴9、日5 × 兴9、兴5 × 兴9、兴6 × 和6家系在胸径与材积性状上均表现较好,与其他多数家系存在显著差异 (P<0.05)。材积方面,4个家系平均值为0.039 5 m3,高出参试家系平均值19.70%,高出混种对照3.95%,高出材积较小的日3 × 石51家系51.92%。其中,材积最高的日12 × 兴9家系高出参试家系平均值39.39%,高出混种对照21.05%,高出种源对照白刀山、小北湖、乌伊岭分别为76.92%、91.67%和91.66%,高出长白落叶松自由授粉家系对照长73-18与长73-4分别为64.29%、84.00%,高出材积最低的日3 × 石51家系76.92%。
2.3 杂种落叶松家系优良家系的选择
杂种落叶松各家系材积性状及育种值见表4。
表4杂种落叶松各家系材积性状及育种值
Table 4 Breeding value in volume of hybridL.gmelinifamilies
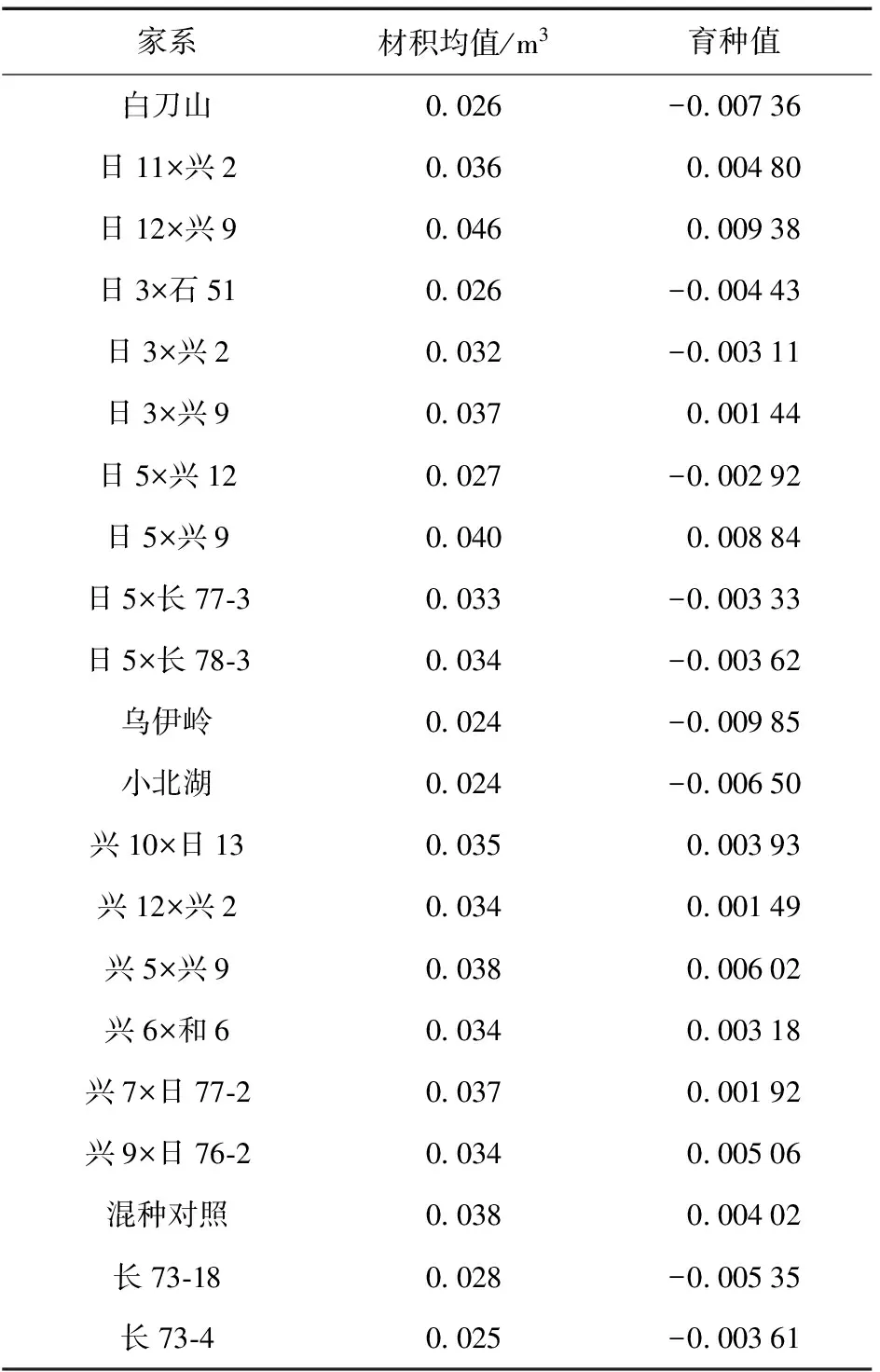
家系材积均值/m3育种值白刀山0 026-0 00736日11×兴20 0360 00480日12×兴90 0460 00938日3×石510 026-0 00443日3×兴20 032-0 00311日3×兴90 0370 00144日5×兴120 027-0 00292日5×兴90 0400 00884日5×长77⁃30 033-0 00333日5×长78⁃30 034-0 00362乌伊岭0 024-0 00985小北湖0 024-0 00650兴10×日130 0350 00393兴12×兴20 0340 00149兴5×兴90 0380 00602兴6×和60 0340 00318兴7×日77⁃20 0370 00192兴9×日76⁃20 0340 00506混种对照0 0380 00402长73⁃180 028-0 00535长73⁃40 025-0 00361
由表4可知,日12 × 兴9、日5 × 兴9、兴5 × 兴9家系育种值相对较大,分别为0.009 38、0.008 84、0.006 02,在所有参试家系中表现较好。结合家系间材积性状差异性比较结果,最终选择出日12 × 兴9、日5 × 兴9、兴5 × 兴9家系作为杂种落叶松优良家系。按14%入选率,选择强度为i=1.59,则遗传增益分别为树高0.230 4、胸径0.402 1、材积0.835 1。
2.4 杂种落叶松家系生长性状间相关性分析
杂种落叶松家系生长性状间相关分析见表5。
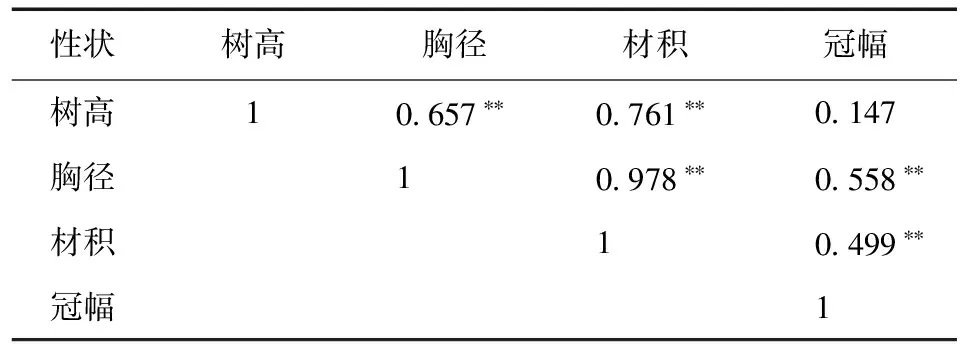
表5 杂种落叶松家系生长性状间相关分析Table 5 Correlation analysis of growth traits of hybrid L.gmelini families
注:**表示相关性极显著。
由表5可知,生长性状间均为正相关关系。其中,冠幅与树高相关性未达到显著水平,与胸径、材积存在极显著正相关关系 (P< 0.01),相关系数分别为0.558和0.499,说明冠幅对杂种落叶松的径生长具有积极的促进作用,对高生长的影响弱于对径生长的影响。
3 结论与讨论
本研究中,杂种落叶松家系生长性状存在较丰富的变异,各性状家系平均变异系数均在20%以上,材积性状变异最大,为58.87%,这为杂种落叶松家系生长性状的人工选择与遗传改良提供了较好的基础。在各参试家系中,日3 × 兴9家系多数性状变异系数较大,表现出丰富的变异,说明该家系内个体间优劣分化较为明显,在以家系均值为标准的比较中,其优势未得到充分显现,在今后进一步人工选育的过程中,对日3 × 兴9家系可重点进行家系内选择,以获取一定数量的优良单株。孙晓梅等[12]在短轮伐期日本落叶松家系生长性状遗传参数变化的研究中也发现,生长性状不仅在家系水平上存在着极显著的变异,家系内个体间变异也很丰富,具有很大的遗传改良潜力,家系和家系内个体选择是落叶松改良的主体。
本研究中,生长性状家系间差异均达到显著水平 (P< 0.05),多数杂种落叶松家系的材积量显著高于对照 (P< 0.05),生长优势明显。家系遗传力显示胸径与材积家系遗传力较高,受较强遗传控制,树高与冠幅受中等强度遗传控制,这与罗旭等[6]在杂种落叶松生长表现及遗传增益的研究中所得结论相近,其16年生的杂种落叶松子代测定林,树高、胸径、立木材积家系遗传力均达到0.65以上,受较强的遗传控制。因此,家系间差异主要来源于遗传因素,通过人工筛择,杂种落叶松生长性状具有较大的遗传改良潜力。本研究以材积量为主要指标,筛选出日12 × 兴9、日5 × 兴9、兴5 × 兴9家系作为杂种落叶松优良家系,树高、胸径、材积遗传增益分别达到0.230 4、0.402 1和0.835 1,人工选择增益明显,对今后落叶松良种的应用与推广具有积极意义。
杂种落叶松生长性状间均为正相关关系,冠幅与树高相关性未达到显著水平,与胸径、材积存在极显著正相关关系 (P< 0.01),说明冠幅对杂种落叶松的径生长具有促进作用,对高生长的影响弱于径生长。同时,在杂种落叶松林生长到一定阶段时,通过适当疏伐以增加冠幅,可以有效加快林木径生长,对增加材积生长量具有积极的促进作用。这与贯春雨等[13]在杂种落叶松F1代生长性状、木材性状的变异及相关分析,毛桃[14]在马尾松优树子代测定林生长和材质的遗传分析及联合选择的研究中所得结论相似,冠幅与生长性状相关紧密,冠幅越大,树木的长势越好。
[参 考 文 献]
[1] 尹绍鹏, 赵国辉, 夏辉, 等. 长白落叶松半同胞子代测定研究[J]. 西南林业大学学报, 2016, 36(1): 63-68.
[2] 黄鑫春. 辽东落叶松人工林生长规律研究[J]. 西南林业大学学报, 2016, 36(1): 79-83.
[3] 李艳霞. 长白落叶松优树子代生长与材质的遗传变异及多性状联合选择[D]. 哈尔滨: 东北林业大学, 2012.
[4] 孙晓梅, 张守攻, 王卫东, 等. 日本落叶松自由授粉家系形质性状遗传变异的研究[J]. 北京林业大学学报, 2004, 26(3): 41-45.
[5] 杜超群, 许业洲, 孙晓梅, 等. 鄂西亚高山区日本落叶松无性系生长性状变异分析与早期选择[J]. 华中农业大学学报, 2015, 34(3): 19-23.
[6] 罗旭, 王祥岐, 张含国, 等. 杂种落叶松的生长表现及遗传增益[J]. 东北林业大学学报, 2005, 33(6): 8-9.
[7] 高扬, 王有菊, 杨世桢, 等. 杂种落叶松优良家系的选择[J]. 中南林业科技大学学报, 2013, 33(10): 57-60.
[8] 孟宪宇. 测树学[M]. 北京: 中国林业出版社, 2006.
[9] 沈熙环. 林木育种学[M]. 北京: 中国林业出版社, 2006.
[10] 刘天颐, 杨会肖, 刘纯鑫, 等. 火炬松基因资源的育种值预测与选择[J]. 林业科学, 2014, 50(8): 60-67.
[11] 孙晓梅, 杨秀艳. 林木育种值预测方法的应用与分析[J]. 北京林业大学学报, 2011, 33(2): 65-71.
[12] 孙晓梅, 张守攻, 侯义梅, 等. 短轮伐期日本落叶松家系生长性状遗传参数的变化[J]. 林业科学, 2004, 40(6): 68-74.
[13] 贯春雨, 张含国, 王福森, 等. 杂种落叶松F1代生长性状、木材性状的变异及相关分析[J]. 防护林科技, 2012(5): 24-26.
[14] 毛桃. 马尾松优树子代测定林生长和材质的遗传分析及联合选择[D]. 南京: 南京林业大学, 2007.
