新疆农田防护林不同林龄生物量及碳储量
2018-04-24朱玉伟桑巴叶王永红陈启民褚奋飞
朱玉伟,桑巴叶,王永红,刘 康,陈启民,褚奋飞
(新疆林科院 造林治沙研究所,新疆 乌鲁木齐 830063)
随着人类大量使用煤、石油等化石能源排放过量的二氧化碳等温室气体,是造成全球气候变化的根本原因,气候变暖及其影响越来越引起人们的关注,全球碳循环是全球气候变化的核心问题之一,森林生态系统碳循环是全球碳循环的重要组成部分[1-5],在CO2收支平衡、缓解气候变化以及维持全球气候环境方面起着十分重要的作用[6-7],而大量研究表明森林生态系统地上部分和地下部分有机碳储量占陆地总碳储量的80%和40%以上[8-9]。应对气候变化,降低大气中CO2含量主要有两条途径,即工业直接减排和森林碳汇间接减排。短期内我国通过工业直接减排的难度较大,森林碳汇投资少、代价低、综合效益大,更具有经济可行性和现实操作性,森林碳储量的估算对增汇减排和全球气候变化意义重大[10]。人工造林被认为是吸收CO2、减缓气候变暖,增加森林生态系统碳汇功能的重要措施[11-12]。农田防护林主要由人工林组成,它在新疆森林生态系统中占有举足轻重的作用,通过开展新疆农田防护林生物量分布特征研究,进一步分析固碳能力,对森林碳汇间接减排具有十分重要的意义,也可为新疆农田防护林经营提供参考。
农田防护林是由树木组成的具有多种功能的,由带状组成的人工绿色网络系统[13],其最高目标是建立或恢复持续而稳定的高生产力水平、高生态效益的农田生态系统。在增强农田生态系统的抗干扰能力、改善农田小气候、净化空气、碳汇和保障作物稳产高产等方面起到重要作用[14-15]。生物量是体现森林生产潜力及固碳潜力重要而直接的指标[16],研究不同树种、不同林龄农田防护林生物量、碳储量、碳密度的变化规律,对于掌握农田防护林生态系统不同发育阶段的生产力、产量结构、分配特点及固碳增汇能力,揭示不同时期影响防农田护林生产力和生物量的主导因素等均有重要指导意义。由于新疆农田防护林树种单一,杨树占80%以上,过去对杨树生物量方面的研究主要集中在杨树总生物量和各组分生物量变化规律、生物量预测方法、模型以及与其他相关环境因子之间关系方面[17-19],对不同林龄下农田防护林各树种生物量、碳密度、碳储量及其分配特征研究甚少。影响森林生态系统碳储量大小的因子为气候、林龄、林分类型以及森林群落结构,而树种是决定森林生态系统碳储量大小的一个重要生物学因子[20],但对不同造林树种之间碳储量、碳密度的林龄格局方面的研究还少有报道[21]。基于此,本文拟以新疆农田防护林为对象,利用不同林龄下的新疆主要农田防护林树种解析木的实测数据,探讨其生物量、碳密度、碳储量组成、分配以及各组分生物量、碳储量、碳密度随年龄的变化规律,并分析农田防护林主要造林树种和林龄结构下的碳储量和碳密度差异,综合分析和评价新疆农田防护林主要树种不同林龄结构下的固碳功能,为已增加碳汇为目的的人工造林和农田防护林经营管理提供依据。
1 研究地区与研究方法
1.1 研究区概况
新疆(34°25′—49°10′N,73°40′—96°23′E) 位于中国的西北边陲亚欧大陆中部,东西长约2 000 km,南北宽1 600 km,面积165 km2,是中国面积最大的省区,约占全国总面积的1/6。新疆有充足的光热和土地资源,具有发展林业得天独厚的优势,但风沙危害严重。2014年新疆耕地面积为512万hm2,农田防护林面积为36.94万hm2,农田防护林面积占耕地面积的7.21%,新疆绿洲农田防护林体系关系到新疆的耕地面积稳定,是保障粮食安全的生态屏障,在改善小气候、防风固沙、保持农业稳产、高产中起着重要作用。在新疆绿洲,杨树是农田防护林的主要树种,在绿洲内部杨树约占农田防护林面积的80%以上,以杨树为主形成的绿色廊带和农田防护林体系长久以来是维系新疆干旱荒漠绿洲生态稳定的主体和保证,也是新疆木材供给的主要来源。研究区农田防护林林分树种单一,纯林面积占总面积的98%,杨树面积占80%以上,为绝对优势树种,其他的树种主要为榆树和沙枣。新疆主要农田防护林树种包括幼龄、中龄、近熟林、成熟林和过熟林5个龄级,近几年随着更新的加快,过熟林面积很少。因此,本文中过熟林生物量按成熟林计算,林分概况见表1。
1.2 研究方法
1.2.1 样地设置 于2013-2014年6-10月在新疆南、北、东疆典型地区和田地区墨玉县、玛纳斯区域150团场、吐鲁番地区托克逊县设立3个主要农田防护林树种不同林龄标准地,要求立地类型及营林措施一致,标准地为长100 m,宽度根据林带宽度而定,每个林龄2个重复,共32个样地,对标准地中的林木测定树高和胸径,并详细调查样地土壤、植被、郁闭度等生境要素。
1.2.2 标准木选择 采用标准地调查数据结合二类资源清查数据,确定不同树种、不同龄级的平均树高和胸径,选取平均标准木作为测定生物量的标准样木。各树种每个龄级选择标准木两株。
1.2.3 林木生物量测定
1)树干生物量测定。解析木伐倒后,在离地径1.3 m处分段,>15 m的树干按2.0 m长度分段,<15 m的树干按1.0 m长度分段;在地径处及每个分段处截取3.0~5.0 cm厚的圆盘,记录圆盘号和直径,并分别测取每一段树干和圆盘的鲜重。截取的圆盘带回室内置于80℃烘箱烘至恒重,推算树干生物量。
2)树枝及树叶生物量测定。对树枝、树叶进行分层、分级调查,从第1活枝起,将树冠等分为上、中、下3层,在各层内以枝基径≤1.0 cm、1.0~2.0 cm、2.0~4.0 cm、>4.0 cm为标准进行分级,统计各层、各等级枝数,每级选取3个标准枝称取带叶枝鲜重,摘净叶后分别称取枝鲜重和叶鲜重;各级分别取100.0~200.0 g的枝、叶样品各1份,称鲜重后装入自封袋做好标记,带回室内烘干(80℃)至恒质量称重,测定水含率并算出干物质生物量。

表1 研究样地林分概况
3)根系生物量测定。以伐根为中心,分不同方向、层次(≤20.0 cm、20.0~40.0 cm、40.0~60.0 cm、>60.0 cm)挖取树木全部根系,用水冲洗,再根据根系直径≤1.0 cm、1.0~2.0 cm、2.0~5.0 cm、5.0~10.0 cm、>10.0 cm 5类分层分类称重,随机抽取1.0 kg样品,装入自封袋做好标记,带回室内进行烘干(80℃),测定干物质生物量。
1.2.4 有机碳密度测定
1)样品采集。取标准木根、枝、叶、树干各1份,重量200~300 g。
2)有机碳测定。将烘干的植物样品粉碎,用于测定有机碳含量。植物样品采用重铬酸钾-硫酸容量法测定。根据各树种各组分(干、枝、叶、根)生物量与其含碳率的乘积得到各自的碳储量,各树种碳储量为各树种各组分之和。
1.2.5 数据处理 根据野外调查的结果和室内分析的数据,用Excel2010进行数据处理,用SPSS17.0进行显著性和相关性分析。
2 结果与分析
2.1 主要树种不同林龄各组分的生物量及其分配
2.1.1 农田防护林主要树种面积 新疆农田防护林树种主要有杨树、榆树、沙枣及少量软阔和硬阔树种如白蜡、柳树、胡杨、山杏等,主要为杨树、榆树、沙枣,占比超过90%。对以上3个主要树种进行研究,将少量硬阔树种面积归到榆树,软阔树种归到杨树进行统计。从林龄结构上看(表2),3种主要农田防护林树种的面积以幼龄林和中龄林为主,从幼龄林到过熟林逐渐减少。杨树幼龄林和中龄林面积占到总面积的60.54%,榆树幼龄林和中龄林面积占到总面积的96.02%,沙枣幼龄林和中龄林面积占到总面积的84.16%。新疆的农田防护林以杨树为主,杨树面积占总面积的79.62%。由于新疆农田防护林主要树种龄级主要以幼龄林、中龄林、近熟林和成熟林为主,占总面积的96.16%,过熟林面积很小,仅占3.84%,因此,将过熟林面积归到成熟林面积之中计算。
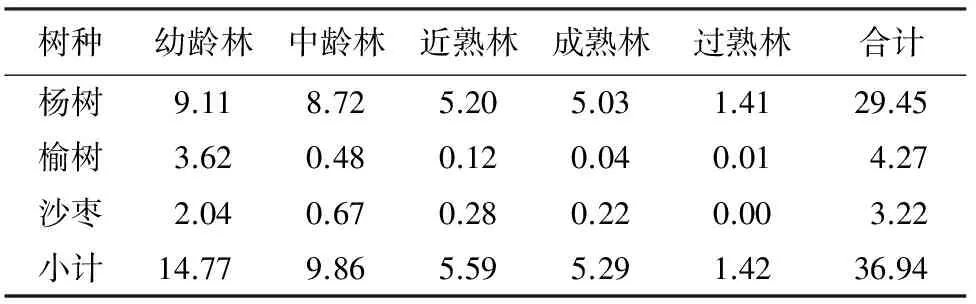
表2 农田防护林主要树种不同林龄面积
2.1.2 主要树种不同林龄各组分生物量分配 主要树种杨树、榆树、沙枣不同林龄各组分生物量及其分配比例不同(表3、图1)。沙枣除幼龄林外不同组分的生物量的大小顺序为树干>根>树枝>树叶,树干居主导地位,体现了其生物量积累的水平,幼龄林不同组分的生物量的大小顺序为树枝>树干>树叶>根,树干和树叶生物量差别不大,原因为幼龄林处于生长初期,光合作用强烈,枝条和叶生长旺盛,营养物质积累迅速,树干生物量积累相对缓慢;杨树不同林龄不同组分的生物量的大小顺序为树干>树枝>根>树叶,树干生物量占绝对优势,说明杨树为速生树种,枝、干生长迅速,生物量积累迅速;榆树不同林龄不同组分的生物量变化规律与上述2个树种有所区别,在4个龄级中,树干生物量占绝对优势,枝和根生物量处于中间,相差不大,叶生物量最小,榆树为深根性树种,根系发达,因此,根系生物量较为庞大。

图1 防护林不同林龄树种各组分生物量垂直变化规律
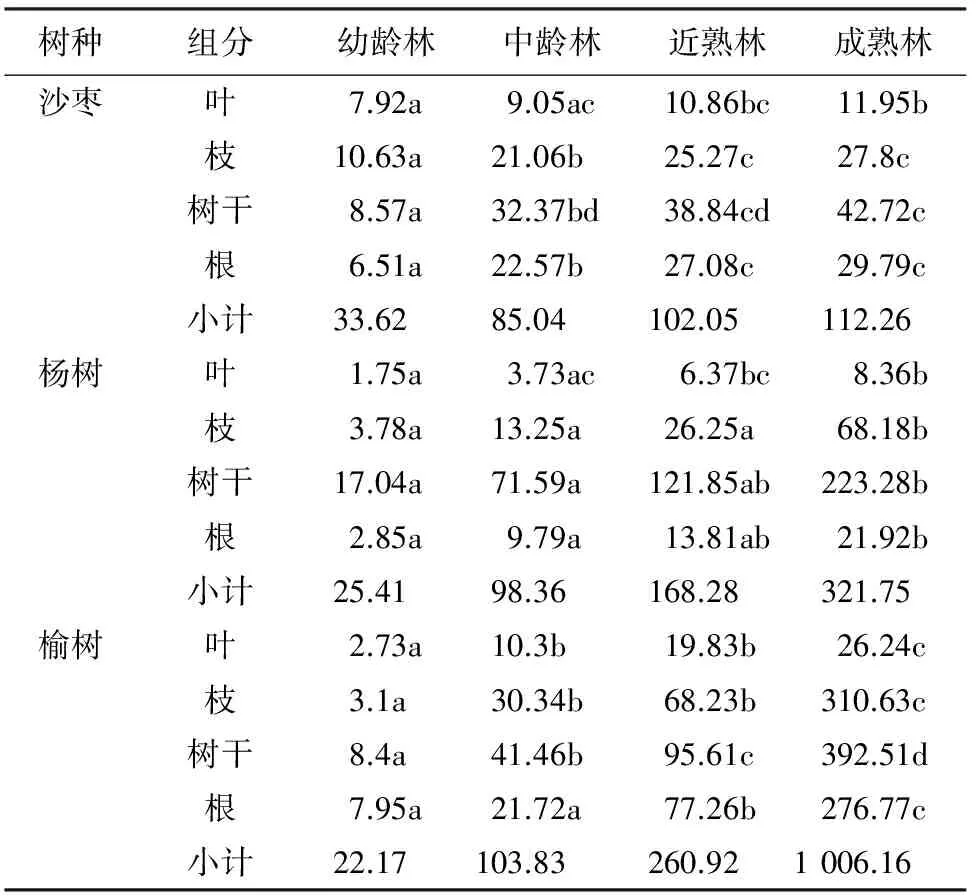
注:均值差的显著性水平为 0.05。
农田防护林3个主要树种树干生物量积累最多但占总生物量的比重不同,沙枣不同林龄树干平均生物量占总生物量的37.02%,杨树为66.20%,榆树为36.90%。农田防护林3个主要树种各组分生物量随着林分年龄的增大增长的幅度不同。沙枣树干成熟林是幼龄林、中龄林、近熟林的4.98、1.32倍和1.10倍。成熟林叶、枝、根生物量是幼龄林、中龄林、近熟林的1.50~4.58倍。沙枣不同龄级各个组分生物量增长不明显,除幼龄林外,中龄林、近熟林、成熟林各个组分生物量增长缓慢,这主要与沙枣生物学特性和新疆特殊的生长环境有关,沙枣在新疆大多生长在风沙前沿,造成该树种生长缓慢,生物量积累较慢;杨树树干成熟林是幼龄林、中龄林、近熟林的13.10、3.12倍和1.83倍,成熟林叶、枝、根生物量是幼龄林、中龄林、近熟林的4.78~18.04倍,说明杨树生长迅速,生物量积累幅度大,原因主要是杨树在新疆主要生长在绿洲内部,水分条件优越,生长的立地条件可满足杨树迅速生长的需要;榆树成熟林树干是幼龄林、中龄林、近熟林的46.73、9.47倍和4.11倍,成熟林叶、枝、根生物量是幼龄林、中龄林、近熟林的9.61~100.20倍,榆树在3个树种中生物量积累幅度最大,是因为榆树从幼龄林到成熟林生长长达60 a以上,在新疆主要生长在绿洲内部,水分条件优越,加之本身的生物学特性,是硬阔叶树种中生长较为迅速的,各组分生物量缓慢增长,但增长幅度较大。
2.2 主要树种不同龄组的碳储量和碳密度
2.2.1 主要树种不同龄组的碳储量 林龄结构是森林结构与功能的主要动力因子,农田防护林主要树种林龄与农田防护林生物量、碳储量、碳密度具有密切关系。研究表明,森林生态系统碳储量随着林龄的增长而增长,成熟林仍然能发挥巨大的碳汇作用[22]。由表2可知,幼龄林和中龄林的面积占到总面积的66.68%,从中看出新疆农田防护林大多都处于幼、中龄林阶段,随着时间推移会不断向成熟林过度,因而固碳潜力巨大。
根据测定的农田防护林主要树种各龄级各组分含碳率(表4),计算出主要树种各龄级各组分碳储量(表5)。从表5可以看出,3种主要造林树种中杨树碳储量最大,榆树高于沙枣但相差不大,主要是杨树面积最大,占农田防护林总面积近80%,榆树面积大于沙枣。3个主要农田防护林树种碳储量除榆树根>树枝外,其余均为树干>树枝>根>树叶,树干为主要碳储量组分。3种主要造林树种由于龄级结构不同,造成碳储量排序不同,杨树各龄组碳储量大小依次为成熟林(2 884.10×104Mg)>中龄林(1 576.46×104Mg)>近熟林(1 325.14×104Mg)>幼龄林(333.83×104Mg),分别占总碳储量的47.17%、25.76%、21.65%、5.46%,碳储量与生物量和含碳率密切相关,各龄级在含碳率在相差不大的情况下,生物量的大小决定碳储量的大小,各树种碳储量的大小与各树种单株生物量和面积呈正相关。成熟林占比最大,原因是成熟林面积虽然只占总面积的21.87%,但单株生物量是幼龄林的12.66倍,中龄林的3.27倍,近熟林的1.91倍,中龄林和近熟林相差不大,原因是中龄林面积大于近熟林,近熟林生物量大于中龄林,幼龄林占比最小,原因是面积虽然最大(面积是成熟林的1.41倍),但单株生物量较小(成熟林生物量是幼龄林的12.66倍),前三者占杨树总碳储量的94.54%,说明新疆农田防护林主要树种杨树成熟林、中龄林和近熟林对其碳储量起主要作用;榆树和沙枣各龄组碳储量大小依次为幼龄林(110.19×104Mg;79.24×104Mg)>中龄林(60.01×104Mg;67.02×104Mg)>近熟林(40.02×104Mg;34.59×104Mg)>成熟林(27.42×104Mg;31.47×104Mg),分别占总碳储量的46.37%和37.32%、25.32%和31.57%、16.84%和16.29%、11.54%和14.82%。榆树和沙枣幼龄林和中龄林碳储量所占比列较大,二者之和分别占榆树和沙枣总碳储量的71.69%和68.89%,说明新疆农田防护林主要树种榆树和沙枣幼龄林和中龄林对其碳储量起主要作用。主要防护林树种面积和平均碳密度的不同是造成差异的主要原因,杨树各龄级面积相差不是太大,但碳密度相差很大,碳密度从幼龄林的36.66 Mg·hm-2增加到成熟林448.25 Mg·hm-2,增加幅度很大,除中龄林外幼龄林到成熟林,碳储量逐步增加,碳储量中龄林>近熟林,主要是由于中龄林面积大于近熟林。榆树、沙枣幼龄林和中龄林面积很大,分别占到了总面积的96.02%和84.16%,因此,造成榆树和沙枣幼龄林和中龄林碳储量较大。
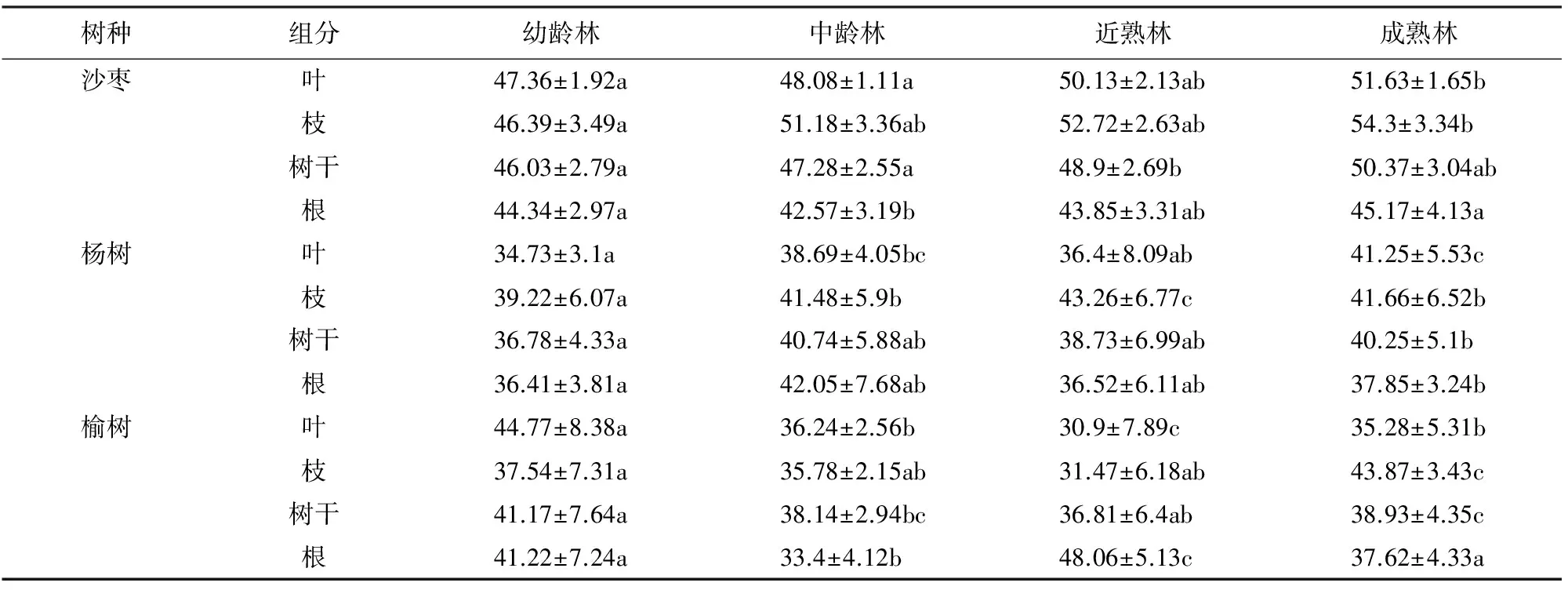
表4 主要树种不同林龄各组分含碳率
2.2.2 主要树种不同龄组的碳密度 从表6可以看出,3个主要农田防护林树种碳密度从幼龄林到成熟林随着林龄的增加而增加,碳密度是单位面积的碳储量,增加的原因主要是不同树种从幼龄林到成熟林生物量的增加,沙枣从幼龄林的38.78 Mg·hm-2增加到成熟林的140.60 Mg·hm-2,杨树从幼龄林的36.66 Mg·hm-2增加到成熟林的448.25 Mg·hm-2,榆树从幼龄林的30.40 Mg·hm-2增加到成熟林的555.15 Mg·hm-2,这表明农田防护林生态系统的碳素在不断的积累。3个主要农田防护林树种不同龄级碳密度均为树干>树枝>根>树叶,表明树干是3个主要农田防护林树种碳密度的主要组分,如沙枣中龄林树干碳密度占沙枣整株碳密度的38.22%、杨树中龄林树干占66.08%、榆树中龄林树干占41.98%,从中可看出树干对整株树种碳密度的贡献。
2.3 主要树种的碳储量和碳密度
由于3种主要农田防护林树种的面积、单位面积生物量、龄组结构等的不同,其碳储量必然存在差异,3种主要农田防护林树种中(表5),杨树的碳储量最高(6 119.53×104Mg),其次为榆树(237.63×104Mg),沙枣为(212.31×104Mg)。3种主要农田防护林树种碳储量的差异主要取决于其面积的大小。
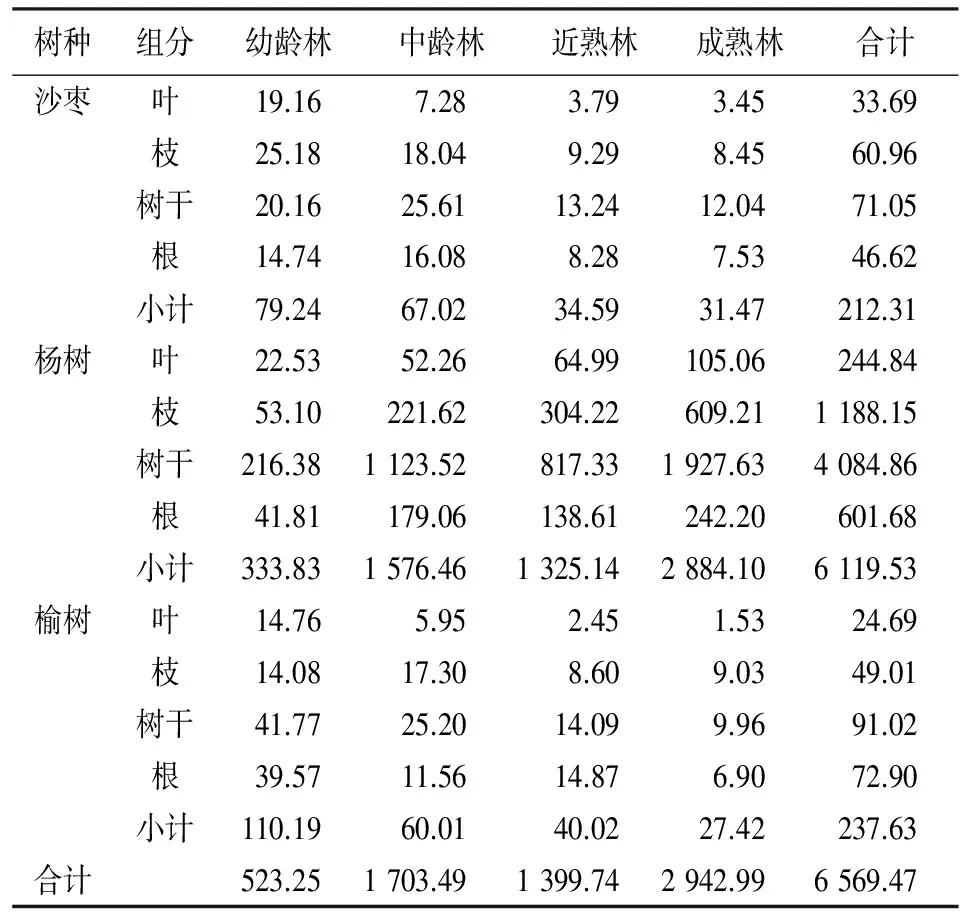
表5 主要树种不同林龄各组分碳储量

表6 主要树种不同林龄各组分的碳密度
3种主要农田防护林树种碳密度也差异明显。其中,杨树平均碳密度最高(207.77 Mg·hm-2),沙枣次之(66.03 Mg·hm-2),榆树最低(55.65 Mg·hm-2)。3种主要树种各龄组的碳密度为幼龄林(35.42 Mg·hm-2)<中龄林(172.69 Mg·hm-2)<近熟林(250.18 Mg·hm-2)<成熟林(442.36 Mg·hm-2)。
3 结论与讨论
3种主要农田防护林树种的面积以幼龄林和中龄林为主,杨树、沙枣、榆树幼龄林和中龄林面积之和各占该树种面积的60.54%、84.16%和96.02%。新疆农田防护林以杨树为主,占总面积的79.62%。
农田防护林3个主要树种树干生物量积累最多,并且随着林分年龄的增大而增大,各组分生物量随着林分年龄的增大而增大,但增长的幅度不同。
新疆农田防护林的总碳储量为6 569.47×104Mg,发挥了较强的碳汇功能。主要树种的碳储量大小取决于面积和各树种生物量,且于树种、林龄结构密切相关,杨树的碳储量最高(6 119.53×104Mg),其次为榆树(237.63×104Mg),沙枣为212.31×104Mg。
研究结果表明,3种主要农田防护林树种在增加新疆森林碳储量上发挥了明显作用。新疆农田防护林3种主要树种以幼龄林和中龄林为主,二者共占农田防护林总面积的66.70%,因此,农田防护林固碳潜力巨大。若能对现有的林分加以合理的森林抚育和管理,随着时间的推移,林龄结构将会发生很大的变化,将能更好地发挥森林碳汇功能[22]。3种主要农田防护林树种,除杨树外,其余均以幼龄林的碳储量最大,其次是中龄林。杨树以成熟林的碳储量最大,其次是中龄林,这与杨树面积和林龄有关,与桑巴叶林龄是影响林分碳储量积累的主导因子之一的研究结果相同[23]。3种主要农田防护林树种随林龄增加,碳密度也呈增长趋势,随着时间的推移,碳储量将进一步增加。
我国人工林普遍存在质量低下、树种组成单一及龄组结构不合理等问题[24],新疆农田防护林也存在同样问题。新疆农田防护林碳密度和碳储量增加的潜力巨大,如何估算碳汇增加的潜力及如何根据碳汇潜力的变化调整森林经营管理措施,都是未来应该思考和解决的重要技术问题[25]。未来应加强新疆农田防护林的抚育和管理水平,充分发挥其碳汇功能。随着新疆农田防护林幼、中龄发展成为近、成熟林,需针对不同的树种类型,控制其成熟林的面积比例、促进各龄组碳密度的增长,农田防护林碳储量也将进一步增加。同时应合理增加新造林面积以增加新疆农田防护林碳储量。
参考文献:
[1] 李峰,杨素青,李惠敏,等.林业碳汇项目及其在黑龙江省的发展前景[J].防护林科技,2008(2):72-73.
[2] BUNKER D E,DECLERCK F,BRADFORD J C,etal.Species loss and above ground carbon storage in a tropical forest[J].Science,2005,310(5750):1029-1031.
[3] 胡会峰,刘国华.森林管理在全球CO2减排中的作用[J].应用生态学报,2006,17(4):709-714.
HU H F,LIU G H.Roles of forest management in global carbon dioxide mitigation.[J].Chinese Journal of Applied Ecology,2006,17(4):709-714.(in Chinese)
[4] 冯瑞芳,杨万勤,张健,等.人工林经营与全球变化减缓[J].生态学报,2006,26(1):3870-3877.
FENG R F,YANG W Q,ZHANG J,etal.Artificial forest management for global change mitigation.[J].Acta Ecology Sinica,2006,26(1):3870-3877.(in Chinese)
[5] GIBBS H K,BROWN S,NILES J O,etal.Monitoring and estimating tropical forest carbon stocks:making REDD a reality[J].Environmental Research Letters,2007,2(4):1-13.
[6] 刘国华,傅伯杰,方精云.中国森林碳动态及其对全球碳平衡的贡献[J].生态学报,2000,20(5):733-740.
LIU G H,FU B J,FANG J Y,etal.Carbon dynamics of Chinese forests and its contribution to global carbon balance[J].Acta Ecology Sinica,2000,20(5):733-740.(in Chinese)
[7] 王邵军,阮宏华.全球变化背景下森林生态系统碳循环及其管理[J].南京林业大学学报:自然科学版,2011,35(2):113-116.
WANG S J,RUAN H H.Review on carbon cycle of forestry ecosystem and its management under the global changes[J].Journal of Nanjing Forestry University(Natural Science Edition),2011,35(2):113-116.(in Chinese)
[8] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳储量和碳平衡[J].植物生态学报,2000,24(5):518-522.
ZHOU Y R,YU Z L,ZHAO S D,etal.Carbon storage and budget of major chinese forest types[J].Acta Phytoecologica Sinica,2000,24(5):518-522.(in Chinese)
[9] STICKLER C M,COE M T ,COSTA M H,etal.Dependence of hydropower energy generation on forests in the Amazon Basin at local and regional scales[J].Proceedings of the National Academy of Sciences ,2013,110(23):9601-9606.
[10] 方精云,朱江玲,王少鹏,等.全球变暖、碳排放及不确定性[J].中国科学:地球科学,2011,41(10):1385-1395.
FANG J Y,ZHU J L,WANG S P,etal.Global warming,human induced carbon emissions,and their uncertainties[J].Science China:Earth sci.,2011,41(10):1385-1395.(in Chinese)
[11] PIAO S L,FANG J Y,CIAIS Petal.The carbon balance of terrestrial ecosystems in China[J].Nature,2009,458(7241):1009-1013.
[12] 杨晓菲,卢绍伟,饶良懿,等.中国森林生态系统碳储量及其影响因素研究进展[J].西北林学院学报,2011,26(3):73-78.
YANG X F,LU S W,RAO L Y,etal.Advances in the researches of carbon storage of ecology and related factorsin China[J].Journal of Northwest Forest University,2011,26(3):73-78.(in Chinese)
[13] 邓荣鑫,张树文,李颖.基于田间尺度的东北农田防护林防护效应分析[J].生态学杂志,2009,28 ( 9 ):1756-1762.
DENG R X,ZHANG S W,LI Y,etal.Protective effects of faim land shelterbelts in Northeast China:a field scale analysis[J].Chinese Journal of Ecology,2009,28 ( 9 ):1756-1762.(in Chinese)
[14] WILLIAMS-GUILLEN K,PERFECTO I,VANDER MEER J.Bats limit insects in a tropical agroforestry system[J].Science,2008,320:70.
[15] NAIR P,KUMAR B M,NAIR V D.Agroforestry as a strategy for carbon equestration[J].Journal of Plant Nutrition and Soil Science,2009,172:10-23.
[16] 汪森.森林生态系统碳循环研究进展[J].安徽农业科学,2013,41(4):1560-1563.
WANG S.Research progress of carbon cycling of forest ecosystem[J].Journal of Anhui Agri,2013,41(4):1560-1563.(in Chinese)
[17] 范志平,曾德慧,朱教君,等.农田防护林生态作用特征研究[J].水土保持学报,2002,16( 4):130-140.
FAN Z P,ZENG D H,ZHU J J,etal.Advance in characteristics of ecological Effects of farmland shelterbelts[J].Journal of Soil Water Conservation,2002,16( 4):130-140.(in Chinese)
[18] 姜凤岐,朱教君,曾德慧,等.防护林经营学[M].北京:中国林业出版社,2003:52-58.
[19] 赵天锡,陈章水.中国杨树集约栽培[M].北京:中国科学技术出版社,1994:346-353.
[20] 郭超,周志勇.太岳山森林碳储量随树种组成的变化规律[J].生态学杂志,2014,33(8):2012-2018.
GUO C,ZHOU Z Y.The alteration of carbon stock of forest ecosystem by tree species composition in Taiyue Mountain[J].Chinese Journal of Ecology,2014,33(8):2012-2018.(in Chinese)
[21] 李奇,朱建华,冯源,等.中国主要人工林碳储量与固碳能力[J].西北林学院学报,2016,31(4):1-6.
LI Q,ZHU J H,FENG Y,etal.Carbon stocks and carbon sequestration capacity of the main plantations in China[J].Journal of Northwest Forest University,2016,31(4):1-6.(in Chinese)
[22] 顾丽,郑小贤,龚直文.长白山森林植被碳储量与碳汇价值评价[J].西北林学院学报,2015,29(6):46-51.
GU L,ZHENG X X,GONG Z W.Dynamic change of forests carbon storage and corbon sink evaluation in the over-cuting region of Changbai Mountains[J].Journal of Northwest Forest University,2015,29(6):46-51.(in Chinese)
[23] 桑巴叶,朱玉伟,陈启民,等.新疆杨不同林龄农田防护林的碳储量[J].南方农业学报,2015,46(8):1455 -1461.
SANG B Y,ZHU Y W,CHEN Q M,etal.Carbon storage ofPopulusalbavar.pyramidalisabunge farmland shelter forest with different ages[J].Jourmal of Southerm Agriculture,2015,46(8):1455-1461.(in Chinese)
[24] 徐济德.我国第八次森林资源清查结果及分析[J].林业经济,2014(3):6-8.
XU J D.The 8th forest resources inventory results and analysis in China[J].Forestry Economics,2014(3):6-8.(in Chinese)
[25] 魏晓华,郑吉,刘国华,等.人工林碳汇潜力的新概念及应用[J].生态学报,2015,35(12):1-8.
WEI X H,ZHENG J,LIU G H,etal.The concept and application of carbon sequestration potentialsin plantation forests[J].Acta Ecology Sinica,2015,35(12):1-8.(in Chinese)
