外源6-BA对大红三角梅生长、开花及生理的影响
2018-04-24田高飞闫淑君金妍超吴艳芳段嵩岚
田高飞,闫淑君,金妍超,刘 震,吴艳芳,段嵩岚
(福建农林大学 园林学院,福建 福州 350002)
三角梅属(Bougainvillea)又称叶子花属、三角花属、九重葛属、宝巾属等,是紫茉莉科(Nyctaginaceae)的一类常绿攀援灌木,统称为三角梅、叶子花、三角花、九重葛、勒杜鹃、宝巾、刺仔花、南美紫茉莉等[1]。由于花色丰富多样、花期较长、一年四季皆可开花,三角梅在热带和亚热带地区广受欢迎[2],但是在实际应用中经常出现只长叶不开花、株型散乱、花期不一致等现象,影响观赏,国内往往依靠人工修剪、控水、遮光等物理手段调控花期[3-4],而对于化学调控措施即应用生长调节剂调节生长和开花的研究鲜有报道[5-6]。
6-苄基腺嘌呤(6-BA)是细胞分裂素类的生长调节剂,能够控制细胞分裂、调控新陈代谢,在植物的生长发育过程中发挥重要功能[7]。此外,还有研究表明6-BA具有促进植物花芽分化、解除顶端优势、提高花朵品质等作用[8-9]。植物由营养生长转入生殖生长到开花是一连串生理及形态变化的复杂过程,这期间植物受体内外各种因素的共同作用;碳水化合物为植物的能量来源,早期Klebs提出的碳氮比学说即认为植物体内要有较多的碳水化合物才可开花;氮是植物体内重要的营养元素,可溶性蛋白质是重要的含氮化合物,是氮素的主要存在形式,可溶性蛋白质是花器官形态建成的物质基础,是花芽分化进程中的重要生理生化指标[10]。本试验采用不同浓度的外源6-BA对三角梅进行处理,研究其对三角梅生长及开花的影响,以及其在调控过程中与可溶性糖、淀粉和可溶性蛋白质等生理生化指标的内在联系,以便为实际应用管理提供参考依据。
1 材料与方法
1.1 试验材料
供试材料于2016年9月10日选购自福州市建新花卉市场,为1年生大红三角梅(Bougainvilleaspectabilis‘Crimsonlake’)扦插苗,平均株高53.71 cm,采用常规办法进行养护管理,盆土表面干燥时进行浇水,浇水时应浇透,以盆底有水流出为宜。试验地点为福建农林大学园林学院下安3号实验楼楼顶露天天台,位于25°15′-26°39′N,118°08′-120°31′E,属于亚热带海洋性季风气候,温暖湿润,雨量充沛,年平均气温16~20℃,其中1、2月份最冷,7、8月份最热,极端最高气温为42.3℃,极端最低气温为-2.5℃,年均日照1 700~1 980 h,年均降水量900~2 100 mm,年相对湿度约为77%。
1.2 试验方法
试验于2016年9月19日开始,选取1年生扦插苗,每盆1株,于2016年9月22日开始施用6-BA(北京索莱宝科技有限公司),用针筒吸取滴于三角梅茎顶,每周2次,共2周。6-BA处理采用200 mg·L-1(A1)、500 mg·L-1(A2)、1 000 mg·L-1(A3)3个浓度梯度,以蒸馏水为对照(CK),每处理3个重复,每重复2株。
1.2.1 形态指标 每株随机选取3个枝条,并在最上端的叶片标记,2016年9月19日至2016年12月26日期间每2周调查1次枝条长度、枝条节数,计算枝条平均生长率和节间长,开花时则调查始花日期(植株第1朵花开放日期)、末花日期(植株最后1朵花凋谢日期),计算始花天数和花期天数,于盛花期(植株50%以上的花朵开放)统计每株的花序数与花朵数,测量盛开花朵苞片以及成熟叶片的长和宽,采用直尺测量,长为基部至顶尖的距离,宽为最宽处的距离。
枝条生长率(cm/d):调查期间标记枝条的总净生长量(cm)/调查天数(d)
节间长(cm):最后一次调查时标记枝条长度(cm)/枝条节数
始花天数(d):始花日期-施药日期
花期天数:末花日期-始花日期
1.2.2 生理生化指标 2016年9月21日至2016年12月28日期间每2周取样1次,取所选枝条中部的功能叶片,测定可溶性糖、淀粉和可溶性蛋白质含量。测定方法参考高俊凤[11]的方法,可溶性糖和淀粉含量采用蒽酮比色法测定,可溶性蛋白质含量采用考马斯亮蓝G-250法测定。
1.3 数据处理
用Excel 2007进行数据整理和图表制作。将6-BA作为单一因素,采用SPSS Statistics 19.0做ANOVA分析,揭示不同浓度6-BA对三角梅生长、开花及生理的影响,用LSD检验指标在各处理间的差异显著性,并用字母标记法表示差异显著性,处理间平均值差异显著水平P<0.05。
2 结果与分析
2.1 不同处理对大红三角梅生长的影响
试验结果表明(表1、图1):A1处理下的枝条生长率和节间长均小于对照,而A2和A3处理下则大于对照,但各处理与对照相比均未达到显著差异水平,说明6-BA对大红三角梅枝条生长和节间长的影响效果不明显;与对照相比,各处理均能增加试验植株叶片的长度,其中A1、A2差异不显著,A3达到显著差异水平,而各处理对于叶片宽度没有显著影响。这表明,6-BA对大红三角梅株型等生长方面的影响作用并不大。
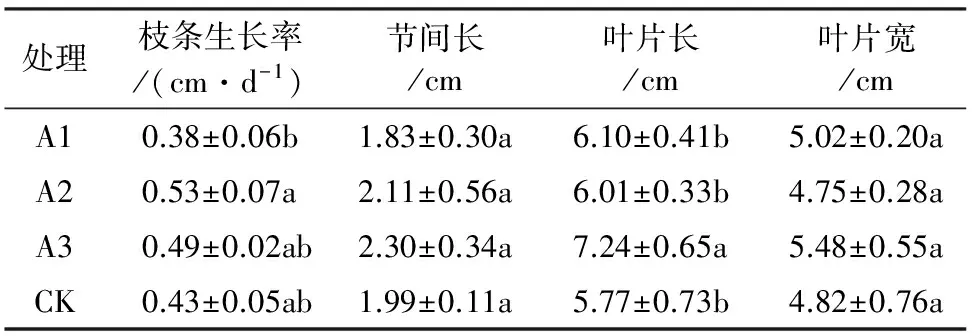
表1 不同处理对大红三角梅生长的影响(平均值±标准差)
注:不同字母表示差异显著(P<0.05)。
2.2 不同处理对大红三角梅开花品质的影响
各处理下始花天数均比对照组少(表2、图1),且随浓度增加而减少,其中A3天数最少(26.00±1.73)d,各处理与对照相比均达到显著差异水平,但各处理间差异不显著,说明6-BA具有促进大红三角梅提前开花的效果;对花期天数的分析表明,与对照相比,低浓度(A1)下花期天数减少了1 d,而高浓度(A2、A3)下则增加了约4 d和5 d,但并未达到显著差异水平;与对照相比(表2和图2),各处理对花苞片长度和宽度的影响有减小的趋势,但差异不显著;各处理下大红三角梅花序数量和花朵数量相比于对照组来看均有所减少,且随着浓度的增加,花序数量和花朵数量逐步减少,A3的花序数量和花朵数量最少,分别为12.50±2.12和20.00±1.41,且均呈显著差异水平。这说明6-BA虽然能够促使大红三角梅提前开花,但使其花苞片长度和宽度有减小的趋势,同时会减少开花数量,特别是在高浓度(A3)下,其花序数量和花朵数量均显著减少。

注:A:200 mg·L-1处理;B:500 mg·L-1处理;C:1 000 mg·L-1处理;D:对照处理。图2同。
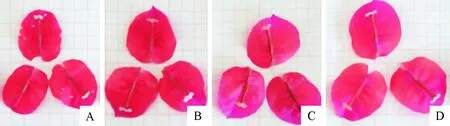
图2 不同处理花苞片大小
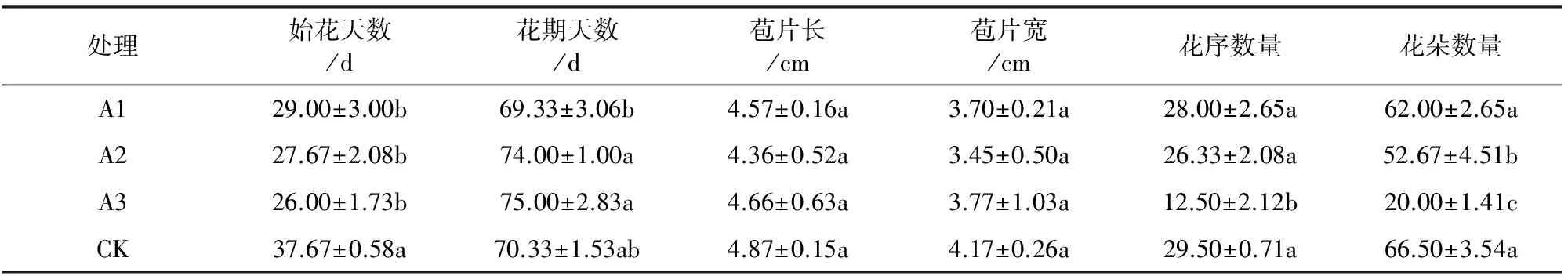
处理始花天数/d花期天数/d苞片长/cm苞片宽/cm花序数量花朵数量A129.00±3.00b69.33±3.06b4.57±0.16a3.70±0.21a28.00±2.65a62.00±2.65aA227.67±2.08b74.00±1.00a4.36±0.52a3.45±0.50a26.33±2.08a52.67±4.51bA326.00±1.73b75.00±2.83a4.66±0.63a3.77±1.03a12.50±2.12b20.00±1.41cCK37.67±0.58a70.33±1.53ab4.87±0.15a4.17±0.26a29.50±0.71a66.50±3.54a
注:不同字母表示差异显著(P<0.05)。
2.3 不同处理叶片可溶性糖含量的变化
各处理在试验期间大红三角梅叶片可溶性糖含量的变化情况基本一致(图3),总体呈先上升后下降再上升的趋势。各处理于9月21日至10月19日期间变化幅度很小,10月19日至11月2日,各处理含量急剧增加,于11月2日达最大峰值,之后急剧下降,除CK外其余各处理均于11月16日达到低谷值,而后开始上升,CK则在11月30日达低谷值后开始明显上升,其低谷值出现时间比其余各处理延迟约2周。结合对花期观察可知,可溶性糖含量开始大幅度增加至最大峰值的时间段与大红三角梅初花期及盛花初期的时间段基本吻合,说明在开花初期可溶性糖含量明显增加,以便为成花进程提供能量。
11月2日之前,各处理含量均高于对照,A3、A2含量保持在较高水平,11月2日及之后基本上为A2、A1含量保持在较高水平。方差分析和多重比较表明(表3),在最高峰值各处理间含量差异明显,除A3外,其余各处理与对照相比均达显著差异水平,A1、A2之间以及A2、A3之间也均达显著差异水平,其中A2含量最高,为(30.70±0.76)mg·g-1FW。整体来看,低浓度(A1、A2)下的作用效果比较好,可溶性糖含量整体保持在较高水平。
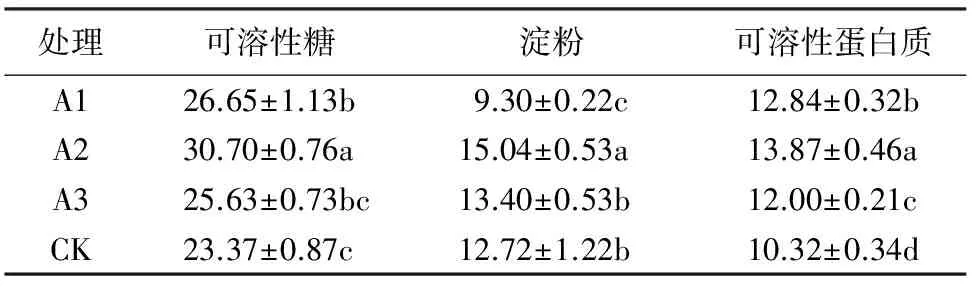
2.4 不同处理叶片淀粉含量的变化
如图4所示,各处理在试验期间大红三角梅淀粉含量的变化情况基本一致,总体呈上升—下降—上升—下降的趋势。各处理含量于9月21日至11月2日期间变化不大,11月2日至12月28日期间,各处理含量变化较为明显,基本呈“M”型变化趋势,A1于11月16日达到最大峰值,其余各处理则于12月14日达到最大峰值。可以看出,大红三角梅淀粉含量在开花前期也出现较大幅度变化,但时间晚于可溶性糖,可能是由于可溶性糖与淀粉二者之间的存储转化机制导致。
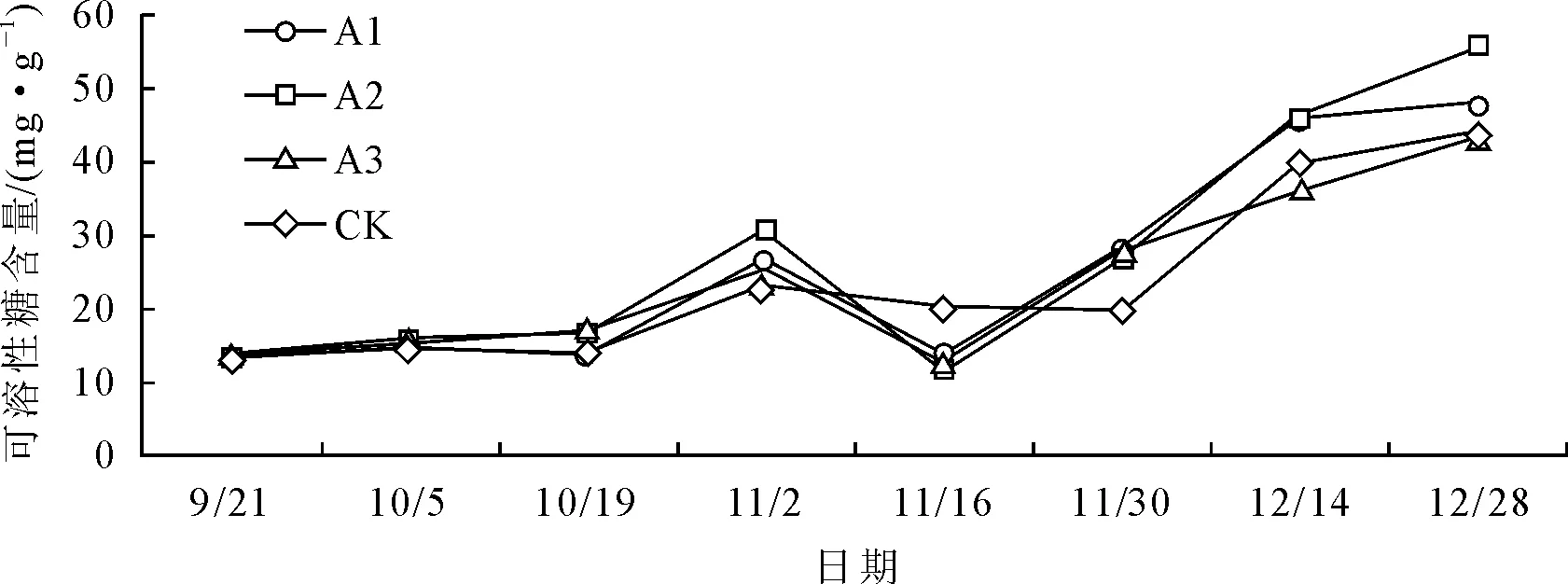
图3 不同处理对叶片可溶性糖含量的影响
mg·g-1
注:不同字母表示差异显著(P<0.05)。
11月16日之前,各处理含量均高于对照,A3、A2含量保持在较高水平,11月16日及之后基本上为A2、A1含量保持在较高水平。方差分析和多重比较表明(表3),在最高峰值A2含量最高,为(15.04±0.53)mg·g-1FW,且与对照相比达到显著差异水平,A1、A2、A3之间也均达显著差异水平。整体来看,低浓度(A1、A2)下的作用效果比较好,淀粉含量整体保持在较高水平。
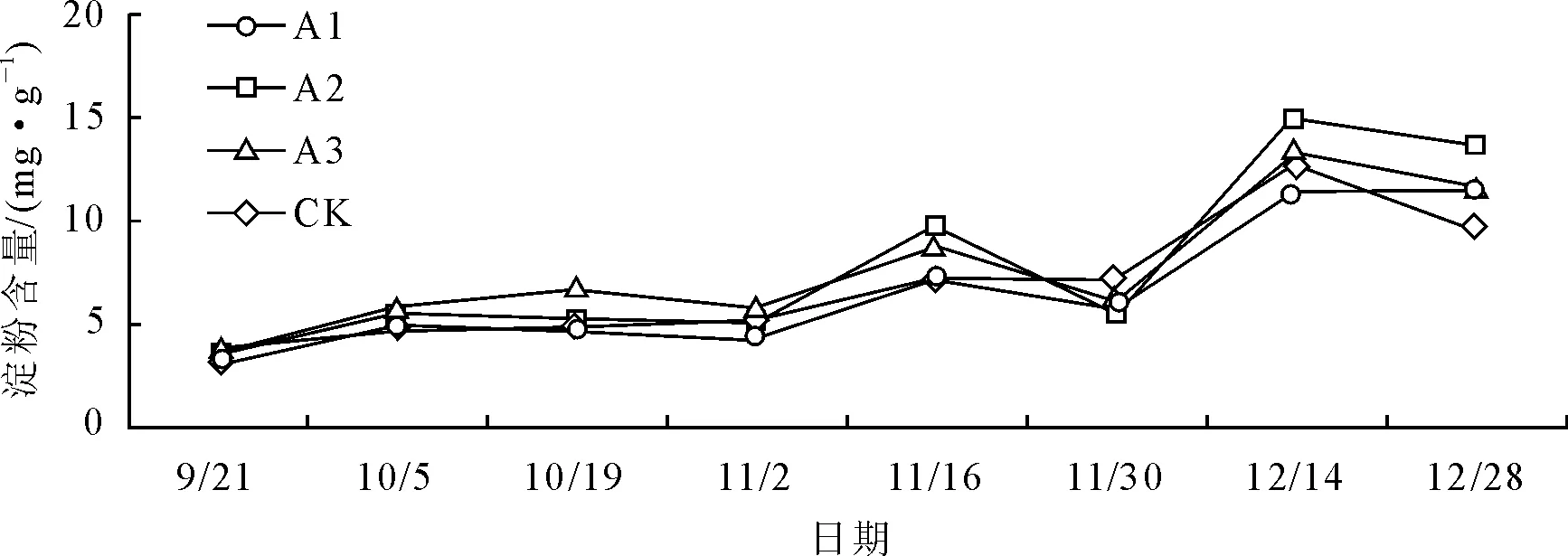
图4 不同处理对叶片淀粉含量的影响
2.5 不同处理叶片可溶性蛋白质含量的变化
由图5可知,各处理在试验期间大红三角梅叶片蛋白质含量的变化情况基本相同,总体呈先上升后下降再上升的趋势。9月21日至10月5日期间,各处理含量基本不变;10月5日后各处理含量显著增加并于11月2日达到峰值,之后各处理含量开始急剧下降,除CK外的其余各处理于11月16日出现低谷值之后开始逐渐上升,而CK则于11月30日出现低谷值后开始逐渐上升,其低谷值出现时间比其余各处理延迟约2周。结合花期观察同样可以看到,在大红三角梅初花期及盛花初期可溶性蛋白质含量出现急剧上升趋势,说明可溶性蛋白质含量在成花初期大幅度增加并参与花器官形态的构建,为花器官进一步分化和成熟提供物质基础。
11月2日之前,各处理含量均高于对照,A3、A2含量保持在较高水平,11月2日及之后基本上为A2、A1含量保持在较高水平。方差分析和多重比较表明(表3),在最高峰值各处理间含量差异明显,所有处理间均达显著差异水平,其中A2含量最高,为(13.87±0.46)mg·g-1FW。整体来看,低浓度(A1、A2)下的作用效果比较好,可溶性蛋白质含量整体保持在较高水平。
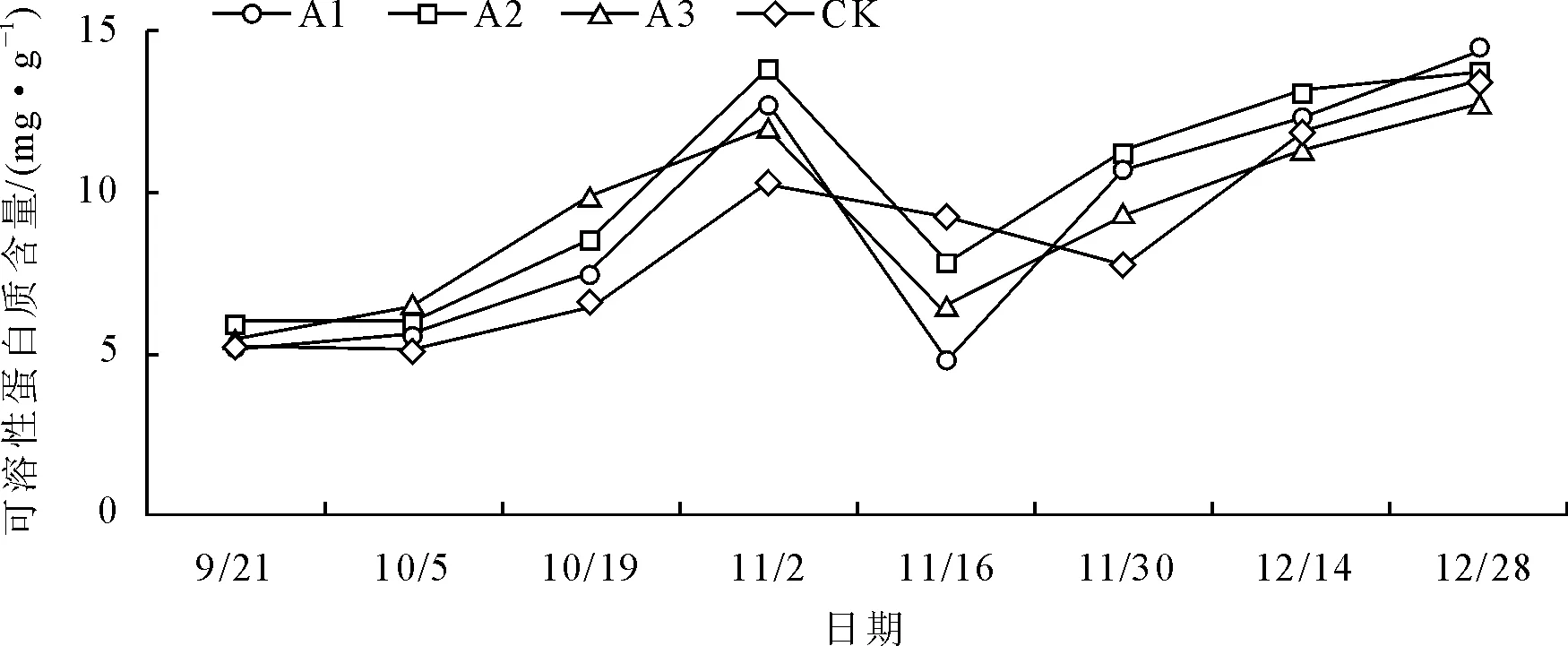
图5 不同处理对叶片可溶性蛋白质含量的影响
3 结论与讨论
适量施用植物生长调节剂可以提高植株观赏效果,调节生长和开花、提高品质[12],这作为观赏植物研究领域的一个重点历来都受到相关研究者的重视[8-9]。本研究结果显示,外源6-BA仅对大红三角梅叶片长的影响较为明显,而对于叶片宽、枝条生长和节间长影响均不显著,这与王贵元[13]等、孔德政[9]等的研究结果差异较大,说明6-BA对植物生长的影响效果可能因植物种类的不同而有差异。开花方面,黎维诗[14]等的研究表明,6-BA能够促使春石斛花芽分化,起到催花的效果,但对其花期方面的影响不显著;孔德政[9]等则发现,6-BA可以促进碗莲提前开花,这些与本试验中6-BA能显著促进大红三角梅提前开花的研究结果相类似;但黎维诗[14]等的研究中还指出,6-BA能够增加春石斛的开花数量,孔德政[9]等对碗莲的研究中也有类似结果,这与本试验的相关研究结果不同,可能是不同种植物之间作用效果不同,亦或作用效果的程度不同所造成,也有可能是施用的浓度过高而产生药害所致,后续试验可以较低浓度的6-BA作为研究重点。
以可溶性糖和淀粉为主的碳水化合物以及可溶性蛋白质是植物体内重要的营养物质,对植物的生长和新陈代谢至关重要,其含量的动态变化可以反映出植物的生长发育状态[10]。可溶性糖和淀粉作为植物体内能量的载体对植物花芽的分化形成和开花具有重要意义,在花芽分化前,首先是碳水化合物在叶片内的积累[15]。张姝媛[16]等对华北紫丁香在花芽分化期间营养物质含量变化的研究显示,华北紫丁香在花芽生理分化期间可溶性糖和淀粉等含量达到最大值,而在花芽形态分化期间含量下降后又逐步上升,说明花芽分化前期碳水化合物的积累有利于花芽的孕育,而在花芽形态分化期间碳水化合物的消耗则为花芽进一步的分化和发育提供能量基础。李亚杰[17]的研究表明,牡丹在花期内可溶性糖和淀粉含量于开花前期含量逐渐增加,盛花期后开始下降。本试验中大红三角梅叶片可溶性糖含量在11月2日之前总体处于上升趋势,在11月2日达到最大峰值之后又开始下降;淀粉的变化稍有不同,其最大峰值出现时间较可溶性糖晚,推测与可溶性糖和淀粉之间的存储转化机制有关。11月2日至11月16日基本上处于大红三角梅初花期和盛花初期,这一时段可溶性糖和淀粉含量大幅度增加说明植物体内在开花前期通过碳水化合物的积累为花器官的进一步分化和成长提供能量基础,这与前人的研究结果具有相似之处;11月16日后其叶片可溶性糖和淀粉含量总体保持上升趋势,这与前人的研究中开花后期可溶性糖和淀粉含量开始下降的结果不同,推测是11月16日之后气温开始较大幅度下降,大红三角梅需要较高含量的可溶性糖和淀粉提供能量以维持自身的生命活动所致。
可溶性蛋白质作为衡量植物代谢水平的重要指标,在花芽分化和开花过程中占有重要的位置[18]。谢利娟[19]等在对毛棉杜鹃花芽分化期间营养物质的变化研究中发现,可溶性蛋白质含量在花芽分化期间呈先急剧下降后缓慢上升的趋势,这可能是花芽形成需要可溶性蛋白质一定程度的积累,之后急剧下降则与花芽形态分化相关。本试验中可溶性蛋白质含量的变化趋势与可溶性糖的基本一致,各处理均于11月2日达到最大峰值,说明开花前期可溶性蛋白质的积累有利于其参与花器官等的构建以及维持其正常的新陈代谢活动,是促使植物加快向生殖生长方向转变的重要因素,这与前人的相关研究结果基本相同;11月16日之后,各处理含量总体呈上升趋势,其原因前面已讨论过,可能是受环境温度降幅较大所致。
6-BA作为一种应用广泛的植物生长调节剂,其在调节植物生长开花过程中与植物体内各生理生化指标的含量变化密切相关。周鑫[20]的研究表明,施用6-BA等植物生长调节剂可增加彩色马蹄莲开花过程中可溶性糖和蛋白质的含量;朱广慧[21]等的研究表明,6-BA可以使东方百合切花可溶性蛋白质含量维持较高水平,并延缓其降解;孔德政[9]等的研究显示,6-BA能够显著提高碗莲花期内SOD和CAT的含量;刘杜玲[22]等则于核桃雌花盛花期施用6-BA,发现能明显提高叶绿素含量。本试验结果显示,施用6-BA后各处理在初花期之前3种生理生化指标含量整体高于对照,A3处理的含量保持在较高水平,使其促进提前开花的效果最为明显,但初花期之后则基本保持在较低水平,而A2、A1处理的含量则基本保持在较高水平。推测A3处理前期各营养物质的积累加速了花芽分化和生长的进程,使其提前开花,但后期营养物质含量不足,无法满足开花枝条的营养需求从而无法诱导分化更多花芽,甚至使部分花芽产生消蕾,导致花序和花朵数量明显减少。从该试验的整体结果来看,低浓度(A1、A2)的6-BA对大红三角梅叶片各生理生化指标含量的变化影响较为明显,使其总体保持较高水平,其中A2处理效果最好。
综上所述,施用外源6-BA可以影响植物开花过程中相关生理生化指标含量的变化,进而达到调控生长和开花的效果。生长方面,6-BA对大红三角梅生长的影响效果不明显;开花方面,6-BA能够提前大红三角梅的开花时间,但随着浓度的增加而会使花序数量和花朵数量显著减少,花苞片长和宽也有减小的趋势;从提前开花时间方面看,各处理均能显著提前开花时间,1 000 mg·L-1处理提前效果最好;从延长花期方面看,500、1 000 mg·L-1处理能够适度延长花期,且1 000 mg·L-1处理效果好于500 mg·L-1处理,但二者与对照相比均为达到显著差异水平;从开花数量方面看,各处理均能使开花数量减少,其中1 000 mg·L-1处理开花数量最少,且与对照相比达到显著差异水平。因此,这要求实际生产应用者根据观赏目的的不同做出合理选择,以便达到最好的观赏效果。综合看来,500 mg·L-1处理的效果最好,其各项生理生化指标含量基本维持在较高水平,既能使其开花时间显著提前,适当增加花期天数,又不至于使开花数量严重减少,维持较高的开花品质,满足观赏效果。
植物的生长开花和体内各生理代谢活动是一个复杂的过程,同时受体内外各种因素的影响,除了受植物激素的影响外还可能受植物品种、当地气候环境等因素的影响。本试验仅对大红三角梅进行了研究,其结果是否具有普遍性还需要进一步研究。此外,本试验施用的6-BA浓度可能较高,后续试验可以较低浓度的6-BA作为研究重点。
参考文献:
[1] 田高飞,焦航,段嵩岚,等.三角梅属(Bougainvillea)研究进展[J].北华大学学报:自然科学版,2017,18(2):248-254.
[2] 徐夙侠,王亮生,舒庆艳,等.三角梅属植物的生物学研究进展[J].植物学通报,2008,25(4):483-490.
[3] 李旺南.三角梅落地栽植花期控制技术[J].福建热作科技,2012,37(3):46-48.
[4] 唐玉贵,朱积余,黄亚光,等.宝巾花花期调控技术研究[J].西部林业科学,2006,35(1):36-39.
[5] 赵家昱,潘远智,李永红.外源ABA对叶子花开花及内源ABA合成关键酶的影响[J].园艺学报,2014, 41(10):2085-2093.
ZHAO J Y,PAN Y Z,LI Y H.Effects of abscisic acid on flowering and some key enzymes ofBougainvilleaglabra[J].Acta Horticulturae Sinica,2014,41(10):2085-2093.(in Chinese)
[6] 陈庭,王运华,王爱敏,等.多效唑对簕杜鹃生长及耐旱性的影响[J].西南农业学报,2014,27(1):296- 302.
[7] 李欣欣,廖红,赵静.GA3、ABA和6-BA对大豆根系生长的影响[J].华南农业大学学报,2014,35(3):35-40.
[8] 钱桦,刘燕,郑勇平,等.施用6-BA对春石斛花芽分化及内源激素的影响[J].北京林业大学学报,2009,31(6):27-31.
[9] 孔德政,申雪莹,孟伟芳,等.外源激素对碗莲开花及酶活性的影响[J].东北林业大学学报,2015,43(3):79-82.
[10] 韦莉,彭方仁,王世博,等.蝴蝶兰‘V31’花芽分化的形态观察及几种代谢产物含量的变化[J].园艺学报,2010,37(8):1303-1310.
[11] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:140-144.
[12] 李艳,杨群力,原雅玲,等.木本曼陀罗北方引种后的栽培技术探讨[J].西北林学院学报,2013,28(2):96-100.
[13] 王贵元,李冰.不同浓度6-BA和GA3处理对枇杷实生苗生长的影响[J].安徽农学通报,2015,21(7):28-29,32.
[14] 黎维诗,柯海丽,邓小果,等.6-BA和GA3处理对春石斛催花的影响[J].热带作物学报,2011,32(6):1016-1019.
[15] 张建铭,谈锋,陈京.大花栀子花芽生理分化期内源激素和碳氮比的动态变化[J].西南师范大学学报:自然科学版,1999,24(2):219-224.
[16] 张姝媛,郭金丽,秦永生,等.华北紫丁香花芽分化期营养物质与核酸含量的变化[J].华北农学报,2008,23(4):179-183.
[17] 李亚杰.牡丹花期生理生化特性研究[D].洛阳:河南科技大学,2012:19-21.
[18] 赵秋芳,陈娅萍,顾文亮,等.香草兰花芽分化期蛋白质及碳水化合物变化研究[J].热带作物学报,2015,36(6):1053-1058.
[19] 谢利娟,王定跃,孙敏.毛棉杜鹃花芽分化期叶片C、N、P质量分数的变化[J].东北林业大学学报,2009,37(9):45-47.
[20] 周鑫.GA3、6-BA、NAA对彩色马蹄莲生长的影响[D].成都:四川农业大学,2012:27-30.
[21] 朱广慧,唐蓉,邓波,等.6-BA对东方百合切花采后生理的影响[J].南方农业学报,2012,43(11):1754-1757.
[22] 刘杜玲,焦普生.生长调节剂对核桃座果率及叶绿素含量的影响[J].西北林学院学报,2006,21(2):87-89.
