长链非编码RNA LINC00467促进非小细胞肺癌细胞的增殖
2018-04-24霍小蕾田向阳张毅强贾建桃韩玲娜
裴 振,霍小蕾,田向阳,张毅强,贾建桃,韩玲娜
(长治医学院:1. 生理学教研室;2. 组织胚胎学教研室;3. 附属和平医院;4. 生化教研室;5.病理生理学教研室,山西长治 046000)
肺癌是世界范围内发病率和死亡率最高的恶性肿瘤,全世界每年约有180万新发病例[1]。肺癌根据病理学形态和临床特征又可分为小细胞肺癌(small cell lung cancer, SCLC)和非小细胞肺癌(non-small cell lung cancer, NSCLC),其中NSCLC患者约占80%~85%[2-3]。目前,NSCLC的治疗手段仍然是以手术治疗为主。随着治疗技术的日臻成熟,尽管治疗方法有较大改进,但是NSCLC的5年生存率仍然不能令人满意,仅为15%[4-5]。因此,亟需一种新的NSCLC治疗靶标,以更好监测其复发和转移等病程。长链非编码RNA(long non-coding RNAs, LncRNA)是一类近年来被发现与多种疾病密切相关的分子[6-7]。LncRNAs可定位于细胞核或细胞质或者特定亚细胞器上,其定位与其行使的功能密切相关[8]。研究显示,LncRNA家族分子LINC00467(long intergenic non-protein coding RNA 467)在调控神经母细胞瘤、结直肠癌和肺腺癌的发生发展中发挥了重要作用[9-11]。然而,LncRNA-LINC00467在NSCLC中的研究鲜有报道。本研究通过检测LINC00467在NSCLC中的表达、定位及分子调控方式,初步探究LINC00467对NSCLC生物学行为的作用机制,为其分子治疗提供理论基础。
1 材料与方法
1.1材料Trizol试剂和逆转录试剂盒(miScriptⅡRT Kit,#218161)购于QIAGEN公司;逆转录试剂盒(reverse transcription system,#A3500)购于Promega公司;LipofectAMINE 2 000购买于Invitrogen公司;细胞周期与细胞凋亡检测试剂盒(Cell Cycle and Apoptosis Analysis Kit)、CCK8试剂购于碧云天生物科技公司;细胞浆/核RNA抽提试剂盒(PARISTMKit Protein and RNA Isolation system)购于Ambion生物公司;CyclinD1、CDK6和GAPDH抗体(Cell Signaling Technology)购于上海优宁维生物科技有限公司;siRNA片段购于广州瑞博生物有限公司。
1.2方法
1.2.1一般资料 收集25例长治医学院附属和平医院肿瘤科自2014年12月至2016年1月NSCLC患者经手术切除的组织,包括癌组织和癌旁组织(癌周3 cm以上)。患者年龄35~75岁,平均(55.23±3.45)岁。患者术前均未进行放化疗和生物治疗、未合并其他肿瘤等,均可完整收集患者的检查、治疗和手术资料以及联系方式等。标本首先置于液氮灌中10 min,然后取出放于-80 ℃保存用于抽提RNA,行qRT-PCR法检测LncRNA-LINC00467的表达情况。
高通量基因表达(GEO)公共数据库中GSE19804和GSE18842数据集在PubMed GEO DataSets主页(http://www.ncbi.nlm.nih.gov/gds/)下载获得,这两个数据集的平台来源为GPL570,其基因表达谱芯片分析均采用Affymetrix Human Genome U133 Plus 2.0 Array,LINC00467对应的探针号为224443_at。GSE19804中包含60对人NSCLC患者癌组织及癌旁组织,GSE18842中包含正常人肺组织45例,NSCLC组织46例。
1.2.2细胞系 人正常支气管上皮细胞系HBE及人NSCLC细胞系A549、NCI-H266、NCI-H1299均购自中国科学院上海生命科学研究院细胞资源中心。HBE、A549、NCI-H266和H1299细胞所用培养液为含100 mL/L胎牛血清(FBS)的RPMI1640培基,细胞保持在37 ℃、50 mL/L CO2的潮湿环境中。
1.2.3细胞转染 将生长状态良好的A549细胞接种于6孔板中(按1.5×105个细胞/孔),用对照序列negative control sequence(si-NC)和si-LINC00467(5 μmol/L)干扰试剂进行细胞转染,具体步骤参照LipofectamineTM转染试剂脂质体2 000试剂盒说明书。转染36 h后,收集细胞及上清进行相应检测。相应siRNA片段的序列分别为:si-LINC00467-1:5′-GAAGAAGAGAAGAGAGAAA-3′;si-LINC00467-2:5′-GATAAGAAGTCCACTCACA-3′;si-NC:siN05815122147。
1.2.4qRT-PCR检测LINC00467、CyclinD1和CDK6 mRNA表达采用Trizol法提取各组细胞的总RNA,反转录得到cDNA,进行实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)检测。引物序列如下:LncRNA-LINC00467 mRNA的正向引物为5′-ATTGAAGATGCTGCCAAGGG-3′,反向引物为5′-GCCCAGTTTCAGTCCCTCTT-3′;CyclinD1 mRNA的正向引物为5′-TCGTTGCCCTCTGTGCCACA-3′,反向引物为5′-GCAGTCCGGGTCACACTTGA-3′;CDK6 mRNA的正向引物为5′-CCAGATGGCTCTAACCTCAGT-3′,反向引物为5′-AACTTCCACGAAAAAGAGGCTT-3′;U6(细胞核的参照物) mRNA的正向引物为5′-CTCGCTTCGGCAGCACA-3′,反向引物为5′-AACGCTTCACGAATTTGCGT-3′;18S(细胞质的参照物)mRNA的正向引物为5′-GTGGGCCGAAGATATGCTCA-3′,反向引物为5′-TTGGCTAGGACCTGGCTGTA-3′;GAPDH(细胞质的参照物)mRNA的正向引物为5′-TGCACCACCAACTGCTTAGC-3′,反向引物为5′-GGCATGGACTGTGGTCATGAG-3′。引物根据GenBank序列,用软件Primer Premier 5.0和Oligo 6.22设计,由华大基因生物科技有限公司合成。PCR反应条件:95 ℃ 5 min,随后95 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,重复40个循环。
1.2.5CCK8检测LINC00467 siRNA对细胞增殖活性的影响 各组细胞设置5个复孔,进行相应的处理后。各板加入100 μL混合培养液(90 μL联合培养基+10 μL CCK8溶液/孔),在37 ℃孵育长达2 h,应用SpectraMax 190光吸收酶标仪(美国分子仪器公司)测定450 nm处的吸光度并进行分析。
1.2.6克隆形成实验检测LINC00467 siRNA对细胞增殖活性的影响 各组细胞按每孔1 000个细胞分别接种于6孔板中,培养约15 d后,肉眼可见克隆时即终止培养。PBS磷酸盐缓冲液清洗数次,40 g/L多聚甲醛固定30 min,10 g/L结晶紫染液染色10 min,晾干,拍照。每组设置3个重复。
1.2.7细胞周期分析检测LINC00467 siRNA对细胞增殖活性的影响 收集细胞并用700 mL/L的冷乙醇固定,4 ℃过夜。碘化丙啶(0.05 mg/mL)和RNA酶(2 mg/mL)常温下染色30 min,流式细胞仪分析细胞周期,细胞周期(G0/G1、S和G2/M期)百分比的评估使用Cell Lab Quanta SC软件。
1.2.8细胞核/质RNA分离及检测LINC00467在细胞核/质定位 收集约1×106细胞数量的A549细胞,用预冷的PBS磷酸盐缓冲液清洗数次,离心,重悬,再将细胞分成2份(一份抽提总RNA,一份抽提核/质RNA)。总RNA的抽提采用Trizol法,细胞质/核RNA的抽提步骤参考胞质/核RNA抽提试剂盒(PARISTM Kit Protein and RNA Isolation system)说明书。将qRT-PCR法测得的细胞核RNA样本中LncRNA-LINC00467、U6、GAPDH、18S基因表达量减去各基因在总RNA样本中的表达量,得到△Ct,计算2-△Ct,进行Lg(2-△Ct)转换即得到最终的表达值[12]。当LINC00467表达量大于0时,即表示该分子主要分布在细胞核中;反之,当LINC00467表达量小于0时,在横坐标下方,即表示该分子主要分布在细胞质中。
1.2.9Western blot蛋白免疫印迹分析检测LINC00467 siRNA对细胞周期分子Cyclin D1和CDK6蛋白表达的影响 处理细胞后,用RIPA裂解液(强)冰上裂解细胞30 min,20 W超声振荡2 min,在4 ℃条件下,12 000 g离心10 min,收集上清,检测蛋白浓度。50 μg待测蛋白在100 g/L的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳 SDS-PAGE进行电泳分离,电泳条件为积层胶80 V 40 min、分离胶120 V 1.5 h 90 min,然后在100 V的条件下湿转膜1.5 h 90 min,室温封闭1 h,一抗4 ℃孵育过夜(Cyclin D1和CDK6一抗的稀释比例1∶500,GAPDH一抗的稀释比例1∶1 000),洗膜,二抗37 ℃孵育1 h,洗膜,显影。用Scion Image软件进行条带的灰度分析。
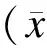
2 结 果
2.1LncRNA-LINC00467在NSCLC组织和正常肺组织中的表达通过提取GEO公共数据库GSE19804和GSE18842数据集的生物信息学资料,分析LINC00467在NSCLC组织和正常肺组织中的表达差异,结果显示:在GSE19804和GSE18842数据集中,LINC00467在NSCLC组织中的表达高于正常肺组织(P均<0.05,图1)。

图1GEO公共数据库分析LINC00467在NSCLC组织和正常肺组织中的表达
Fig.1 The level ofLINC00467 expression in NSCLC tissues and normal lung tissues analyzed by GEO database
2.2LncRNA-LINC00467在NSCLC组织和NSCLC细胞系中的表达及定位为了更进一步明确LncRNA-LINC00467在NSCLC中的表达情况,我们通过qRT-PCR法检测25对人NSCLC组织及其相应的癌旁正常组织。结果显示,相对于癌旁正常组织,LINC00467在NSCLC组织中呈现高表达(n=25,P<0.01,图2);qRT-PCR法检测肺正常支气管上皮细胞系HBE和NSCLC细胞系A549、NCI-H266和NCI-H1299中LINC00467的表达情况,结果显示,LINC00467在NSCLC细胞的表达显著高于正常支气管上皮细胞系HBE(n=3,P<0.05,图3),其中,NSCLC细胞系A549中LINC00467的表达最高。此外,试剂盒抽提分离A549细胞的细胞核RNA和细胞质RNA后,行qRT-PCR法检测相应细胞核/质指标,结果显示LINC00467主要定位于细胞质中(图4),提示其LINC00467可能在细胞质发挥调控某些基因转录后水平或翻译水平的功能。

图2qRT-PCR法检测LINC00467在NSCLC癌组织及癌旁组织中的表达
Fig.2 The expression ofLINC00467 in NSCLC cancer tissues and adjacent normal tissues detected by qRT-PCR (n=25)
2.3验证LncRNA-LINC00467si-RNA在A549细胞中的抑制效率LINC00467 si-RNA瞬时转染非小细胞肺癌A549细胞48 h后,qRT-PCR结果显示,与转染si-NC的细胞相比,转染si-LINC00467-1和si-LINC00467-2组的细胞LINC00467基因的表达水平均受到抑制(n=3,P均<0.05),转染si-LINC00467-1+2组的细胞LINC00467的抑制程度尤为明显,差异有统计学意义(n=3,P<0.01,图5)。

图3qRT-PCR法检测LINC00467在NSCLC癌细胞中的表达
Fig.3 The expression ofLINC00467 in NSCLC detected by qRT-PCR (n=3)
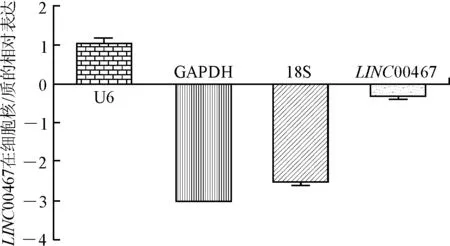
图4qRT-PCR法检测LINC00467在NSCLC细胞系A549中的定位情况
Fig.4 The location ofLINC00467 in NSCLC of A549 cells detected by qRT-PCR
注:表达量在横坐标上方即表示该分子主要分布在细胞核中,表达量在横坐标下方即表示该分子主要分布在细胞质中。

图5qRT-PCR法检测LINC00467si-RNA在NSCLC细胞系A549中的抑制效率
Fig.5 The inhibition efficiency ofLINC00467 si-RNA in NSCLC cell line A549
2.4沉默LncRNA-LINC00467表达对A549细胞增殖的影响LINC00467 siRNA瞬时转染非小细胞肺癌A549细胞48 h后,流式细胞技术结果显示,与对照组si-NC细胞相比,si-LINC00467组处于G0/G1期的细胞比例明显增多(P<0.01),处于S期的细胞比例明显减少,差异有统计学意义(n=3,P<0.01,图6);LINC00467 si-RNA转染处理细胞后培养15 d,细胞克隆形成实验结果显示,转染si-LINC00467细胞组相对于si-NC组,细胞克隆形成明显受到了抑制,且克隆数量明显减少,差异有统计学意义(n=3,P<0.01,图7);此外,CCK8细胞增殖实验结果也显示,转染96 h后,si-LINC00467细胞组相对于si-NC组的细胞增殖活性明显受到抑制,差异有统计学意义(n=5,P<0.01,图8)。提示,沉默LncRNA-LINC00467表达可抑制非小细胞肺癌A549细胞的细胞增殖能力。
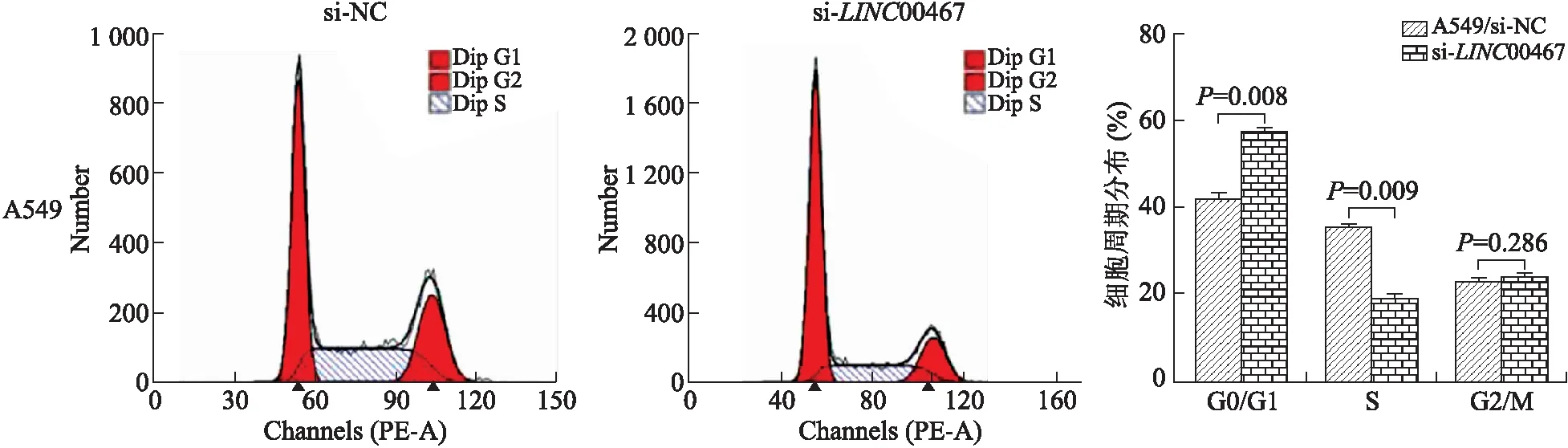
图6流式细胞技术实验检测LINC00467si-RNA对细胞周期分布的影响
Fig.6 Effects ofLINC00467 si-RNA on cell-cycle distribution detected by flow cytometry
图7细胞克隆形成实验检测LINC00467si-RNA对细胞克隆形成的影响
Fig.7 Clone formation experiment to detect the effect ofLINC00467 si-RNA on cell clone formation
2.5沉默LncRNA-LINC00467表达对A549细胞周期分子表达的影响LINC00467 siRNA瞬时转染非小细胞肺癌A549细胞48 h后收集相应的RNA和蛋白质,qRT-PCR法检测结果显示,与对照组si-NC细胞相比,si-LINC00467细胞组中LINC00467、G1/S-特异性周期蛋白-D1(CyclinD1)和细胞周期分子细胞周期素依赖激酶(CDK6)的表达水平均受到了不同程度的抑制,差异有统计学意义(n=3,P均<0.05,图9);Western blot法检测结果也同样表明,si-LINC00467细胞组相对于si-NC组CyclinD1和CDK6的蛋白表达量减少,灰度分析显示差异具有统计学意义(n=3,P均<0.05,图10)。

图8CCK8实验检测LINC00467si-RNA对细胞增殖活性的影响
Fig.8 Effects ofLINC00467 si-RNA on cell proliferation activity detected by CCK8 experiment

图9qRT-PCR法检测LINC00467si-RNA对细胞周期分子的影响
Fig.9 Effects ofLINC00467 si-RNA on CyclinD1 and CDK6 detected by qRT-PCR

图10Westernblot法检测LINC00467si-RNA对细胞周期分子的影响
Fig.10 Effects ofLINC00467 si-RNA on CyclinD1 and CDK6 detected by Western blot
3 讨 论
NSCLC是全球范围内最常见的肺部恶性肿瘤,具有极高的发病率和死亡率,居所有恶性肿瘤之首[13-14]。近来年,我国的NSCLC发病率和死亡率均呈上升趋势,其治疗也面临着严峻的挑战[15]。NSCLC的发病过程涉及多基因、多步骤、多阶段,参与其中的基因及环境影响因素广泛而复杂,因此NSCLC的确切发生机制尚不完全清楚。随着分子生物学技术在疾病的应用逐渐深入,寻找合适、有效的分子标志物和治疗靶点是当前NSCLC治疗的新策略。
LncRNAs是在真核生物中发现的一类转录本长度在200~100 000 nt、缺少完整功能性的开放阅读框(open reading frame, ORF)、仅编码少量功能性短肽的RNAs分子[16-18]。LncRNAs的异常表达或功能缺失与多种疾病有关,如肿瘤、银屑病、糖尿病、冠心病等[6-7]。LncRNA-LINC00467位于人类染色体的lq32.3的位置,有4个转录本,全长3 508 bp,属于基因间的LncRNAs。近年来的研究发现,LINC00467在多种肿瘤中处于紊乱表达状态,武丽娜等[10]检测67例结直肠癌患者组织中的LINC00467,发现结直肠癌组织中LINC00467的表达水平明显高于相应的癌旁组织,但是LINC00467的表达水平与结直肠癌的分级、分期关系不大;类似的,丛壮壮等[11]报道肺腺癌组织中LINC00467的表达水平明显高于相应的癌旁组织,尽管LINC00467的表达水平与肺腺癌的分期无关,但是与肿瘤大小和血管浸润有关。GEO是基于大量累积的基因表达谱的数据而形成的高通量基因表达数据库,为疾病相关基因的分析提供了快捷、高效的途径[19-20]。本研究分析GEO公共数据库GSE19804和GSE18842数据集中LINC00467在NSCLC组织和正常肺组织中的表达差异以及检测25对NSCLC患者癌组织和癌旁组织中的LINC00467表达水平。结果显示,LINC00467在GEO公共数据库NSCLC组织中的表达高于正常肺组织(P<0.05),且在NSCLC癌组织中LINC00467的表达显著高于癌旁正常组织的表达(P<0.05);此外,NSCLC细胞系A549、NCI-H266、H1299中LINC00467的表达也高于肺正常上皮细胞系HBE(P<0.05)。这些结果表明,NSCLC患者的LINC00467表达升高,可能是判断NSCLC发生的潜在标志物。
肿瘤是一类渐进性增殖异常的疾病[21],其发生发展常常伴随着癌细胞的恶性增殖。研究显示,NSCLC中LINC00467的高表达可能与其恶性增殖、侵袭转移和血管生成密切相关,ATMADIBRATA等[9]研究发现,在神经母细胞瘤细胞中转录因子N-Myc可结合至LINC00467的启动子区从而抑制其转录,而LINC00467可通过减少抑癌蛋白DKK1的表达从而增强神经母细胞瘤细胞的存活;此外,武丽娜等[10]报道LncRNA-LINC00467沉默后可抑制结直肠癌细胞的增殖和侵袭转移,而丛壮壮等[11]研究显示,LncRNA-LINC00467沉默后可抑制HUVEC的血管生成能力。本研究通过流式细胞检测技术、CCK8和细胞克隆形成实验,检测在NSCLC细胞系A549中通过siRNA沉默LINC00467的表达后细胞的增殖情况,与对照组si-NC细胞相比,流式细胞检测技术结果显示si-LINC00467组处于G1期的细胞比例明显增多(P<0.05),处于S期的细胞比例明显减少(P<0.01);细胞克隆形成实验结果显示,si-LINC00467细胞组细胞克隆形成明显受到了抑制,且克隆数量明显减少(P<0.05);CCK8细胞增殖实验结果显示,si-LINC00467细胞组的细胞增殖活性明显受到抑制(P<0.05)。这些结果表明,在NSCLC细胞中沉默LINC00467的表达可明显抑制其恶性增殖的生物学表型。
细胞周期蛋白通常包括G1期、S期和M期蛋白3种,G1/S期转化过程中的重要事件之一是CyclinD1与CDK6特异性结合,形成Cyclin D1-CDK6复合体,推进G1期向S期的进程[22-23]。肿瘤细胞的恶性增殖常与细胞周期蛋白紊乱有关,尤其是G1期向S期转化过程中的蛋白异常表达。本研究通过qRT-PCR和Western blot实验检测,结果显示:与对照组si-NC细胞相比,si-LINC00467细胞组中CyclinD1和CDK6的表达水平均受到了不同程度的抑制(P<0.05)。同时,我们在LncRNAs核质定位实验中发现LINC00467主要定位在细胞质中。上述结果显示,LINC00467可在细胞质中通过调控周期相关蛋白的表达从而影响周期进程,进而促进了NSCLC的增殖。本研究通过NSCLC临床样本的检测、细胞表型实验和分子技术检测等研究策略从LncRNA-LINC00467在NSCLC中的表达、定位及分子调控方式,并在体外水平探究了siRNA沉默LINC00467的表达水平后对NSCLC生物学行为的影响,这为后续深入研究NSCLC发生发展的作用机制提供理论基础,并且为LINC00467在NSCLC的治疗提供新的策略。
参考文献:
[1] SIEGEL RL, MILLER KD, JEMAL A. Cancer statistics, 2015[J]. CA Cancer J Clin, 2015, 65(1):5-29.
[2] SEVE P, REIMAN T, DUMONTET C. The role of beta III tubulin in predicting chemoresistance in non-small cell lung cancer[J]. Lung Cancer, 2010, 67(2):136-143.
[3] 高莹,田美丽,宋丽萍. SPHK-1在非小细胞肺癌H460细胞耐药株中的作用[J]. 西安交通大学学报(医学版), 2017, 38(2):172-175.
[4] RAUNGRUT P, WONGKOTSILA A, LIRDPRAPAMONGKOL K, et al. Prognostic significance of 14-3-3 gamma overexpression in advanced non-small cell lung cancer[J]. Asian Pac J Cancer Prev, 2014, 15(8):3513-3518.
[5] MILLER KD, SIEGEL RL, LIN CC, et al. Cancer treatment and survivorship statistics, 2016[J]. CA Cancer J Clin, 2016, 66(4):271-289.
[6] 何笑云,欧春麟,周素娴. 长链非编码RNAs在糖尿病中的研究进展[J]. 中华糖尿病杂志,2016,8(3):187-189.
[7] BEERMANN J, PICCOLI MT, VIERECK J, et al. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches[J]. Physiol Rev, 2016, 96(4):1297-1325.
[8] CHEN LL. Linking long noncoding RNA localization and function[J]. Trends Biochem Sci, 2016, 41(9):761-772
[9] ATMADIBRATA B, LIU PY, SOKOLOWSKI N, et al. The novel long noncoding RNA linc00467 promotes cell survival but is down-regulated by N-Myc[J]. PLoS One, 2014, 9(2):e88112.
[10] 武丽娜,彭小忠,鲁重美. 长链非编码RNA Linc00467在结直肠癌中的表达及影响[J]. 基础医学与临床, 2016, 36(4):520-525.
[11] 丛壮壮,郭仲,秦涛,等. 长链非编码RNA Linc00467在肺腺癌中的表达及功能[J]. 医学研究生学报, 2017, 30(8):834-838.
[12] HUARTE M, GUTTMAN M, FELDSER D, et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response[J]. Cell, 2010, 142(3):409-419.
[13] FERLAY J, SOERJOMATARAM I, DIKSHIT R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012[J]. Int J Cancer, 2015, 136(5):E359-E386.
[14] 党晓敏,孙忠民,杨岚,等. 铁碳纳米粒介导顺铂对肺癌细胞化疗敏感性的影响[J]. 西安交通大学学报(医学版), 2017, 38(4):517-521.
[15] CHEN W, ZHENG R, BAADE PD, et al. Cancer statistics in China[J]. CA Cancer J Clin, 2016, 66(2):115-132.
[16] 何笑云,欧春麟,肖艳华,等. 沙格列汀对高糖诱导内皮细胞中LncRNA-MALAT1表达的影响[J]. 医学研究生学报, 2016, 29(9):902-905.
[17] BAZZINI AA, JOHNSTONE TG, CHRISTIANO R, et al. Identification of small ORFs in vertebrates using ribosome footprinting and evolutionary conservation[J]. EMBO J, 2014, 33(9):981-993.
[18] 陈葳,薛梅. 肿瘤相关长非编码RNA的表达及调控机制的研究进展[J]. 西安交通大学学报(医学版), 2014, 35(2):143-146.
[19] DAVIS S, MELTZER PS. GEOquery: A bridge between the Gene Expression Omnibus (GEO) and BioConductor[J]. Bioinformatics, 2007, 23(14):1846-1847.
[20] BARRETT T, TROUP DB, WILHITE SE, et al. NCBI GEO: Mining tens of millions of expression profiles—database and tools update[J]. Nucleic Acids Res, 2007, 35(Database issue):760-765.
[21] BALKWILL F, MANTOVANI A. Cancer and inflammation: Implications for pharmacology and therapeutics[J]. ClinPharmacol Ther, 2010, 87(4):401-406.
[22] SEMCZUK A, JAKOWICKI JA. Alterations of pRb1-cyclin D1-cdk4/6-p16(INK4A) pathway in endometrial carcinogenesis[J]. Cancer Lett, 2004, 203(1):1-12.
[23] SCHWARTZ GK, SHAH MA. Targeting the cell cycle: a new approach to cancer therapy[J]. J Clin Oncol, 2005, 23(36):9408-9421.
