棉铃抗氧化酶活性影响棉纤维长度发育的研究
2018-04-18杜海烨康娜娜成莎渠云芳黄晋玲
杜海烨,康娜娜,成莎,渠云芳,黄晋玲
(山西农业大学 农学院,山西 太谷 030801)
棉花是一种栽培历史悠久且经济回报高的作物。我国是世界上最大的棉花生产国和消费国,提高棉花产量与品质面临重大问题。植物抗氧化酶系统中最重要的3种酶SOD、POD和CAT在细胞内抗氧化功能各不相同[1](图1),且其作为清除细胞活性氧自由基(ROS)进而影响植物生长发育的关键调节因素之一[2],近年来被很多人作为切入点研究棉花受水涝干旱胁迫[3]、盐胁迫[4]病害胁迫[5]等的抗逆性。此外抗氧化酶还影响着棉纤维初生壁非纤维素多糖类物质合成[6]。而MDA作为细胞呼吸导致的膜脂过氧化产物,从侧面反映着细胞的呼吸活性与抗逆性[7]。
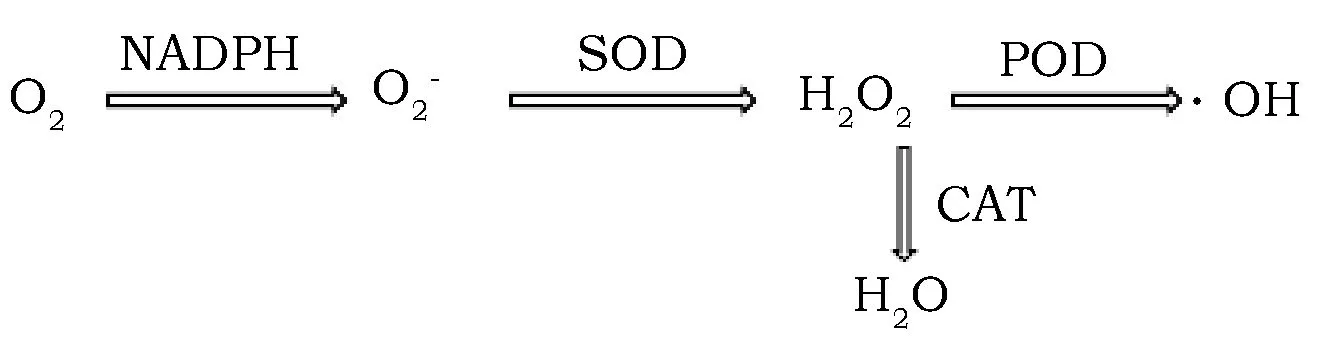
图1 抗氧化酶系统清除ROS过程Fig.1 The cleanning process from the antioxidant enzyme system to ROS
棉铃作为一个有机整体,在铃壳、棉籽和纤维的各器官中进行着物质及信号分子的运输,对整体研究棉花纤维发育具有重要作用[8]。棉纤维的伸长与棉铃的增大相吻合。从开花起,经20~30 d棉花纤维伸长接近最大长度,其中花后5~20 d是纤维伸长的关键阶段,直接影响棉纤维的长度。本文以叶色不同的一对棉花近等基因系为研究材料,以棉铃各器官为研究对象,对其棉纤维伸长阶段抗氧化酶活性以及MDA含量变化进行研究,以期为棉花纤维品质改良提供一定理论依据。
1 材料与方法
1.1 试验材料
本研究选用的材料为叶色不同的一对棉花近等基因系:绿叶、无毛、白絮、棕籽(简称绿叶棉)和红叶、无毛、白絮、棕籽(简称红叶棉)。该对近等基因系来自于亚洲棉(Gossypiumarboreum)×比克氏棉(G.bickii)×陆地棉(G.hirsutum) 3种杂种后代,由山西农业大学棉花育种实验室提供。
参试材料于2016年4月在山西农业大学农作站种植。每个材料种植2行,采用常规的栽培管理措施进行管理。在棉花开花盛期(7月25号)对参试材料同一果枝同一节位当天所开花进行挂牌标记,并于花后10 d与20 d取2种参试材料不同棉株相同部位棉铃各30个,每10个为一次重复,重复3次。每一棉铃分为铃壳、棉籽、棉纤维3部分,液氮速冻后储存于-80 ℃冰箱。
1.2 试验方法
采用NBT(氮蓝四唑)光还原法测定超氧化物歧化酶(SOD)活性[9],在560 nm波长下进行比色测定;采用愈创木酚法测定过氧化物酶(POD)活性[9],在波长470 nm处进行测定,记录吸光度值。酶活性大小以每分钟OD的变化值来表示,其单位为△OD470·g-1FW·min-1;采用高锰酸钾滴定法测定过氧化氢酶(CAT)活性[10],酶活性用每克样品鲜重1 min内分解H2O2的毫克数表示;采用硫代巴比妥法测定丙二醛(MDA)含量[9],在450 nm、532 nm和600 nm处测定其吸光度值,参比为0.5%的硫代巴比妥。
采用分梳法,随机取10粒籽棉将纤维梳直,测定棉纤维长度;取100粒籽棉分别将纤维和棉籽称重计算衣分衣指。
1.3 数据处理
测定数据均采用3次重复均值,使用Excel对数据进行t检验方差分析;使用SPSS对纤维发育指标与测定的各项指标进行Pearson双尾检测相关性分析。
2 结果与分析
2.1 SOD酶活性
棉纤维快速伸长阶段,绿叶棉棉铃各器官中SOD酶活性呈现显著上升趋势,且在铃壳和棉纤维中分别上升了50.6%和287%,达到了极显著差异水平;红叶棉的铃壳和棉籽中SOD酶活性也呈现上升趋势,但在棉纤维中SOD酶活性下降了32%,呈现显著下降趋势(图2)。随着棉纤维的伸长发育,棉铃各器官氧化呼吸作用增强,细胞内产生大量的ROS。因此,棉铃的正常发育需要更高的SOD酶活性。

图2 纤维伸长期2种参试材料棉铃各组织SOD酶活性Fig.2 SOD activity of two materials during fiber elongation periods注:图中小写字母表示在5%水平上差异;大写字母表示1%水平上差异,下同Note:Lowercase letters in the figure indicate differences at the 5% level; capital letters represent 1% level difference,The same below
花后10 d,绿叶棉铃壳中SOD酶活性高于红叶棉且未达到差异水平(P>0.05);棉籽和棉纤维中SOD酶活性低于红叶棉,且纤维中酶含量差异显著。花后20 d,绿叶棉铃壳和纤维中SOD酶活性极显著高于红叶棉,棉籽中红叶棉极显著高于绿叶棉(图3)。从花后10 d到20 d,随着棉纤维的快速伸长,绿叶棉的铃壳和纤维中SOD酶活性显著高于红叶棉,这可能是绿叶棉铃壳和纤维的细胞具有比红叶棉更为活跃的呼吸氧化活动。

图3 纤维伸长期棉铃各组织SOD活性的变化Fig.3 Changes of SOD activity in different tissues of cotton bolls during fiber elongation
2.2 POD酶活性
棉纤维快速伸长阶段,2种参试材料棉铃各器官中(除红叶棉纤维外)POD酶活性逐渐上升,且在红叶棉的铃壳中上升了263%,差异极显著(图4)。在棉铃发育的相应时期,SOD酶活性升高导致细胞内超氧阴离子增加,相应升高的POD酶活性提高了对细胞内超氧阴离子的清除能力。

图4 纤维伸长期2种参试材料棉铃各组织POD酶活性Fig.4 POD activity of two materials during fiber elongation periods
棉花开花后10 d和20 d两个阶段,绿叶棉棉铃各器官中POD酶活性均高于红叶棉同一器官,且铃壳中POD酶活性在花后10 d达到极显著差异,在花后20 d达到显著差异水平;棉纤维中POD酶活性仅在花后20 d达到极显著差异水平;而棉籽中POD酶活性差异在两个时期均没有达到显著水平(图5)。POD参与植物体内多种生理活动,是H2O2的分解剂和解毒物,防止植物细胞受到氧胁迫,绿叶棉中较高的POD酶活性可能与绿叶棉较高的抗逆性有关。

图5 纤维伸长期棉铃各组织POD活性的变化Fig.5 Changes of POD activity in different tissues of cotton bolls during fiber elongation
2.3 CAT酶活性
棉纤维快速伸长阶段,两种参试材料棉纤维中CAT酶活性均呈现下降趋势,且在红叶棉纤维中下降了36%,差异极显著;在铃壳和棉籽中变化趋势不同:绿叶棉铃壳与红叶棉的棉籽呈现下降趋势;绿叶棉的棉籽与红叶棉的铃壳中呈现上升趋势(图6)。

图6 纤维伸长期2种参试材料棉铃各组织CAT酶活性Fig.6 CAT activity of two materials during fiber elongation periods
花后10 d,绿叶棉的铃壳与纤维中的CAT酶活性均显著高于红叶棉且在铃壳中达到极显著差异水平,棉籽中红叶棉CAT酶活性极显著高与绿叶棉;花后20 d,绿叶棉的棉籽与纤维中CAT酶活性显著高于红叶棉,且棉籽中达到极显著差异水平,铃壳中红叶棉CAT酶活性极显著高与绿叶棉(图7)。

图7 纤维伸长期棉铃各组织CAT活性的变化Fig.7 Changes of SOD activity in different tissues of cotton bolls during fiber elongation
2.4 MDA含量
与花后10 d相比,花后20 d绿叶棉铃壳和纤维中MDA含量分别下降了26%和53%,呈现显著降低,而棉籽中MDA含量则略有升高。棉纤维伸长时期,红叶棉棉铃同一器官中MDA含量变化趋势与绿叶棉相同,但红叶棉纤维显著降低,而棉籽中MDA含量显著升高(图8)。棉籽中升高的MDA含量推测与棉籽发育过程中较强烈的氧化呼吸作用导致膜脂过氧化程度升高有关联。

图8 纤维伸长期2种参试材料棉铃各组织MDA含量Fig.8 MDA content in two different materials during fiber elongation periods
棉纤维伸长发育的不同阶段,绿叶棉棉铃各器官MDA含量均低于红叶棉同一阶段的相同器官的含量。 2种材料除花后10 d的铃壳与纤维中MDA含量差异显著外,其他器官之间MDA含量差异均没有显著差异,且MDA含量在铃壳和棉籽中含量较高,纤维中含量较低(图9)。MDA含量是植物细胞膜脂过氧化程度的直接反映,棉纤维伸长的不同阶段,红叶棉棉铃各器官中MDA含量高于绿叶棉,由此可推测红叶棉的膜脂过氧化强度较高。

图9 纤维伸长期棉铃各组织MDA含量的变化Fig.9 Changes of MDA content in different tissues of cotton bolls during fiber elongation
2.5 2种参试材料的品质考察
参试的2种棉花,除百粒籽重外,绿叶棉的百粒籽重、衣指、衣分、纤维的均长均高于红叶棉(表1)。

表1 棉花品质测定Table 1 Determination of cotton quality
2.6 棉铃各项酶活指标与纤维发育相关性分析
对棉铃不同发育阶段各器官的酶活性与纤维发育指标进行相关性分析(表2),结果表明棉纤维伸长发育与SOD、POD和CAT酶活性以及MDA含量具有显著的相关性,但棉铃不同器官以及棉纤维发育的不同时期各器官的酶活指标与纤维发育的相关性大小不同。铃壳与棉纤维中各项酶活指标与棉纤维的伸长发育显著相关,且铃壳中花10 d的SOD酶活性和花后20 d的CAT酶活性以及花后10 d棉纤维中的SOD酶活性与棉纤维的伸长发育呈显著负相关;花后20 d棉籽中的SOD酶活性和CAT酶活性与纤维发育显著相关,且CAT酶活性与纤维发育呈显著负相关;棉铃各器官中MDA含量与棉纤维发育均呈现负相关,且花铃期纤维中达显著水平。

表2 各项指标与纤维发育相关性分析Table 2 Correlation analysis between fiber quality and each index of cotton
注:表中*表示在5%水平上差异;**表示1%水平上差异。
Note:* Indicates a difference at the 5% level;**indicates a 1% level difference.
3 讨论
3.1 抗氧化酶活性与棉纤维发育相关性分析
氧作为植物体内重要的电子受体参与植物多种生命活动,但其接受来自叶绿体和线粒体电子传递链泄露的电子则会形成ROS[11]。活性氧自由基有着比氧更为活泼的化学反应性,会对植物细胞产生氧化胁迫性[12]。长期以来,ROS被视为胁迫代谢的有害产物[13]。但是近年来众多研究发现ROS还能作为信号分子行使调控作用[14],尤其是H2O2可以轻易穿越细胞膜且半衰期较长的特性,使得H2O2在植物发育的许多过程中充当信号分子[15,16]。植物体内,在ROS清除的过程中SOD酶将超氧阴离子(O2-)还原成过氧化氢(H2O2);POD和CAT在细胞内分别将过氧化氢还原成羟基自由基和水,这说明POD和CAT竞争共同底物过氧化氢,并且ROS在引起生物毒性的同时也促进着多糖的代谢[17]。而羟基自由基被认为可以增加细胞壁延伸性,使细胞伸长生长[18];同时羟基自由基也介导生长素的产生,间接促进细胞伸长生长[19]。
本研究中,在棉纤维伸长发育阶段,一方面,绿叶棉纤维中具有POD活性上升,CAT活性下降的趋势;红叶棉纤维中则呈现POD酶活性和CAT酶活性共同缓慢下降趋势。绿叶棉纤维中较高的POD酶活性有利于羟基自由基的积累,而较高的羟基自由基含量对于纤维细胞的伸长生长具有促进作用,从而影响棉纤维长度。另一方面棉纤维的发育是铃壳、棉籽和纤维协同发育的共同结果。铃壳中POD酶活性与棉纤维长度呈现显著正相关[8]。2种参试材料铃壳中SOD与POD酶活性的上升,而CAT变化趋势的不同,这可能导致绿叶棉羟基自由基在棉铃中更多的积累。而由考种数据可知,与红叶棉相比,绿叶棉的纤维较长,这可能与其在棉纤维伸长发育时期较高的羟基自由基含量有关。对棉铃中活性氧代谢产物含量变化的后续研究,将为阐明抗氧化酶对棉花纤维长度的影响机制提供依据。
3.2 MDA含量与棉铃发育相关性分析
活性氧不仅是有氧代谢的副产物,而且在环境胁迫下也能大量形成[20]。植株在逆境胁迫下产生大量活性氧,导致各种生物大分子遭到破坏,改变膜的通透性和流动性,引发膜脂的过氧化反应,影响植物正常的代谢。MDA是膜脂过氧化反应的产物之一,对膜和一些生物大分子如蛋白质、核酸等有较强的破坏性。植物体内的MDA的含量能够反应植物抵御逆境的能力以及细胞膜脂的过氧化程度[21]。
本研究中,在棉纤维伸长发育阶段,绿叶棉棉铃各器官中MDA含量均低于红叶棉,由此推测绿叶棉细胞膜的膜质过氧化程度较低。绿叶棉较高的抗氧化酶系统活性,降低了细胞中MDA含量,促进了纤维素的积累[8]。因而,绿叶棉的衣指与衣分均高于红叶棉。
4 结论
通过一对棉花近等基因系棉铃抗氧化酶活性研究结果表明,棉纤维快速伸长阶段棉铃抗氧化酶活性与棉纤维的伸长发育密切相关。绿叶棉中较高的抗氧化酶活性促进棉纤维的伸长发育,特别是相对红叶棉较高的POD酶活性有利于羟基自由基的形成,促进棉纤维的伸长发育。
[1]Székely G. Oxidative stress in plants[J]. Studia Universitatis Babes-Bolyai, Biologia, 2009, 10(11):19.
[2]张晋龙. 棉花细胞质雄性不育系晋A与亚棉A活性氧代谢研究[D].太谷:山西农业大学,2016.
[3]田又升,王志军,于航,等. 干旱胁迫对不同抗旱性棉花品种抗氧化酶活性及基因表达的影响[J]. 西北植物学报,2015,35(12):2483-2490.
[4]李向前. 棉花耐盐与抗氧化的相关性及H2O2对抗氧化系统的调控[D].乌鲁木齐:新疆大学,2010.
[5]郭海军, 董志强, 林永增,等. 黄萎病对棉花叶片SOD、POD酶活性和光合特性的影响[J]. 中国农业科学, 1995, 28(6):40-45.
[6]杜雄明, 潘家驹, 汪若海. 棉纤维细胞分化和发育[J]. 棉花学报, 2000, 12(4):212-217.
[7]高京草, 王慧霞, 李西选. 可溶性蛋白、丙二醛含量与枣树枝条抗寒性的关系研究[J]. 北方园艺, 2010(23):18-20.
[8]宋世佳,孙红春,张永江,等. 彩色棉抗氧化系统生理特征及纤维素累积对纤维品质的影响[J]. 中国农业科学,2015,48(19):3811-3820.
[9]高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006: 58-66.
[10]叶宝兴,朱新产. 作物科学基础实验[M]. 北京: 高等教育出版社,2007: 447.
[11]del Río L A, Pastori G M, Palma J M, et al. The activated oxygen role of peroxisomes in senescence[J]. Plant Physiology, 1998, 116(4): 1195-2000.
[12]陈俊. 碱地肤幼苗抗氧化酶系统对盐碱混合胁迫的生理响应特点[D].长春:东北师范大学,2006.
[13]Miller G, Shulaev V, Mittler R. Reactive oxygen signaling and abiotic stress[J]. Physiologia Plantarum, 2008, 133(3):481-89.
[14]Mittler R, Vanderauwera S, Gollery M, et al. Reactive oxygen gene network of plants[J]. Trends in Plant Science, 2004, 9(10):490-498.
[15]Bienert G P, Schjoerring J K, Jahn T P. Membrane transport of hydrogen peroxide[J]. Biochimica et Biophysica Acta (BBA)-Biomembranes, 2006, 1758(8): 994-1003.
[16]Vandenbroucke K, Robbens S, Vandepoele K, et al. Hydrogen peroxide-induced gene expression across kingdoms: A comparative analysis[J]. Molecular Biology and Evolution, 2008, 25(3): 507-516.
[17]Liu N, Lin Z, Guan L, et al. Antioxidant enzymes regulate reactive oxygen species during pod elongation in Pisum sativum and Brassica chinensis[J]. Plos One, 2014, 9(2):e87588.
[18]Liszkay A, Van der Zalm E, Schopfer P.Production of reactive oxygen intermediates (O2.-, H2O2,and.OH) by maize roots and their role in wall loosening and elongation growth[J]. Plant Physiology, 2004, 136(2):3114-3123.
[19]Schopfer P, Liszkay A, Bechtold M, et al. Evidence that hydroxyl radicals mediate auxin-induced extension growth[J]. Planta, 2002, 214(6):821-828.
[20]薛鑫, 张芊, 吴金霞. 植物体内活性氧的研究及其在植物抗逆方面的应用[J]. 生物技术通报,2013 (10): 6-11.
[21]谭永安, 柏立新, 肖留斌, 等. 绿盲蝽危害对棉花防御性酶活性及丙二醛含量的诱导[J]. 棉花学报, 2010, 22(5): 479-485.
