投喂频率对花鲈幼鱼胃排空、生长性能和体组分的影响❋
2018-04-13张凯强温海深张美昭李吉方
王 伟, 张凯强, 温海深, 张美昭, 李吉方
(海水养殖教育部重点实验室(中国海洋大学),山东 青岛 266003)
投喂频率对鱼类摄食和饲料转化率具有非常重要的影响。养殖生产过程中,投喂频率过低往往不能满足鱼类生长所需的食物供给,投喂频率过高不仅会增加养殖成本而且会污染养殖水体。因此,为了使养殖鱼类获得正常的生长状态,需要制定适宜的投喂频率。目前,关于鱼类适宜投喂频率主要是通过设置多个不同投喂频率组来进行长期的养殖实验,最后通过比较不同投喂频率组在整个养殖期间所表现出来的生长性能以及食物转化效率等指标来确定的。然而,长时间的养殖实验使鱼类极易受到外界诸如温度、盐度、光照等因素波动的影响从而间接影响鱼类的摄食和生长,对实验结果造成一定偏差;此外,养殖周期过长必将伴随着大量人力物力的投入[1]。由于胃排空实验具有易操作性和快速性等特点,近年来众多学者对多种鱼类胃含物湿重随摄食后时间变化关系进行了研究,建立其各自最佳的胃排空模型(胃排空率)[2-5]。胃排空率这一指标现常被用来评估鱼类的消化功能及饵料的可消化性。另外,由于它的简便性,现也被部分学者用来对鱼类适宜投喂频率的确定进行辅证[6-7]。相关研究报道,通过建立符合鱼类胃排空特征最佳的数学模型,可以进一步了解其食欲恢复程度,在养殖生产实践中这可以为确定饲料投喂量、最佳投喂频率等提供科学的指导。
花鲈(Lateolabraxmaculatus)隶属于硬骨鱼纲(Osteichsthyes)鲈形目(Perciformes)鮨科(Serranidae)花鲈属(Lateolabrax),又称白花鲈、鲈子鱼、板鲈、青鲈、鲈鱼、寨花,主要分布于我国沿海和朝鲜沿海。花鲈因其肉质细嫩、味道鲜美、营养价值高、广温广盐性、抗病力强等特点,深受消费者的欢迎,具有良好的养殖前景。楼宝等[8]对体重为(85.95±2.04)g花鲈幼鱼进行了为期40 d的不同投喂频率养殖实验,与投喂频率研究相似的花鲈鱼种摄食节律的研究也有报道[9-11],但尚未见花鲈胃排空特征研究以及其与养殖实验相结合来确定花鲈幼鱼最佳投喂频率研究的报道。本文以花鲈幼鱼为研究对象,用数学模型拟合所得胃排空实验数据,同时与养殖实验结果进行比对,以期得出花鲈幼鱼最佳的投喂频率,从而为养殖生产提供科学的指导。
1 材料与方法
1.1 实验材料
实验在山东省东营市利津县双瀛水产苗种有限责任公司进行,实验对象为随机挑选的500尾体质健壮、无外伤、体长为(4.78±0.11) cm,体重为(2.02±0.03)g 同一批该育苗场孵化的花鲈幼鱼。先暂养在室内水泥池中,每天饱食投喂2次,待其完全适应后,选取其中的450尾随机放入15个70 L的塑料圆桶(上外径:540 mm下外径:410 mm 高度:460 mm)开始实验。
实验所用饲料为海水仔稚鱼育苗用缓沉型配合饲料(粗蛋白≥53%、粗脂肪≥8%、粗纤维≤3%、粗灰分≤16%、钙≤4%、食盐≤2.5%、总磷≥1.5%、赖氨酸≥2.5%、水分≤9%),粒径1.5 mm。
1.2 实验设计
实验设5个不同的日投喂频率组,分别为1次/2d(F1组)、1次/d(F2组)、2次/d(F3组)、3次/d(F4组)和4次/d(F5组)。每组设3个重复,每个重复放鱼30尾。以白天投喂饵料,具体投喂时间见表1。实验过程中每次将实验鱼投喂至饱食状态,10 min后捞出残饵,70 ℃烘干称重。实验期间记录每天称量的饵料重量和残饵重量。整个养殖过程采用微流水并且24 h连续充气,日换水1次,换水率100%。实验用水为经过砂滤的新鲜海水。实验期间养殖水体各项水质指标为:盐度28~29,温度19~21 ℃,溶解氧5 mg/L以上,pH为7.78~8.24,氨氮及亚硝酸盐含量均小于0.10 mg/L。养殖车间屋顶透光,光照周期为自然的光周期。整个养殖实验持续5周。
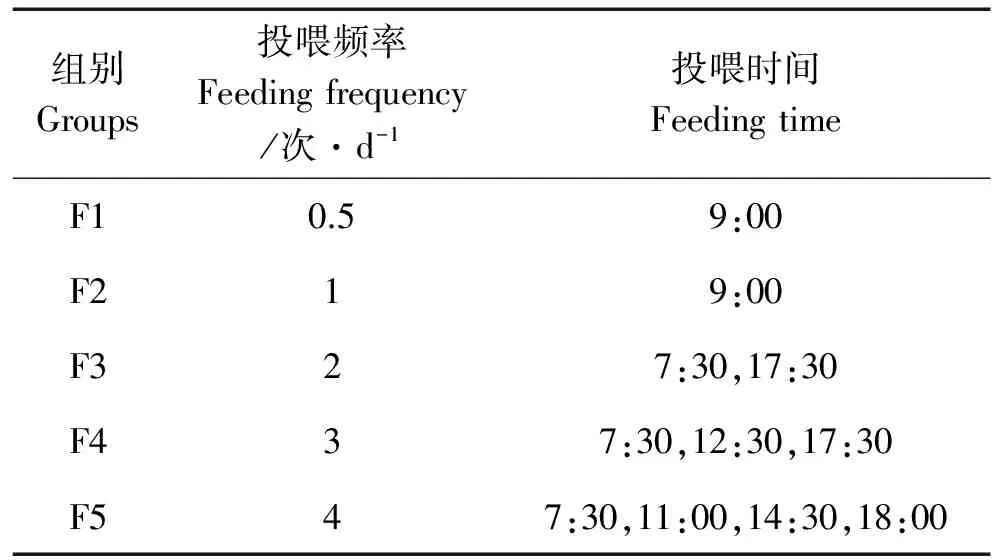
表1 各组花鲈幼鱼投喂时间和投喂频率Table 1 Groups of juvenile Lateolabrax maculatus feeding time and feeding frequency
1.3 样品采集与测定
1.3.1 花鲈幼鱼胃排空实验和胃排空特征及其模型选择 选取260尾平均体长(5.44±0.32)cm,平均体重(2.75±0.27)g活泼健壮、规格较一致的花鲈幼鱼暂养于2个800 L的长方形塑料槽(157 cm×119 cm×58 cm)中。微流水连续充气,水温20℃,每天饱食投喂2次海水仔稚鱼育苗用缓沉型配合饲料,适应1周后开始进行胃排空实验。实验开始前将所有实验鱼饥饿24 h,使其胃排空,然后随机选取20尾鱼解剖观察到胃内均无饲料残余。然后开始投喂海水仔稚鱼育苗用缓沉型配合饲料,将饲料少量多次投入塑料水槽中,使所有实验鱼达饱食状态(继续投饵时不再摄食),待几分钟后水槽内仍有饲料残余,则停止投喂并捞出残饵。然后随机取15尾鱼解剖记录饱食时胃含物重量(湿重),以后每隔1 h取样一次,每次随机取15尾鱼,用100 mg/L的MS-222麻醉后逐一称量体重,然后解剖取出胃含物称重(湿重)。每次所得胃含物重量的平均值作为该取样时间点胃含物的重量。
目前研究中对鱼类胃排空实验数据进行拟合的数学模型种类较多,但最常用的是指数、线性和平方根这3种,通过模型拟合分析其变化规律。3种模型的数学方程式如下:
指数模型:Y=A×e-Bt;
线性模型:Y=A-B×t;
平方根模型:Y0.5=A-B×t。
式中:Y为胃内含物湿重百分比(%)=胃内含物湿重/总摄食量×100;B为瞬时胃排空率;t为摄食后的时间(h);A为常数。
通过比较各拟合模型的相关系数(R2)、残差平方和(RSS)及残差的标准差(SDR)等指标来选择花鲈最佳胃排空模型
1.3.2 养殖实验样本的采集与计算 养殖实验开始和结束前均饥饿24 h,然后统计各处理组鱼的数量,同时每个处理组3个重复随机各取10尾鱼逐一测量体长、体重,液氮保存用于测全鱼体组分。另每个重复再各取10尾鱼逐一测量体长、体重后解剖取内脏和肝脏分别称重,用以计算花鲈幼鱼的形体指标。实验鱼体水分含量采用105℃恒重法 (GB/T 6435-2006)测定,粗蛋白含量采用凯氏定氮法(GB/T 6432-1994)测定,粗脂肪含量采用索氏抽提法(GB/T 6433-1994)测定,粗灰分含量采用550℃灼烧法(GB/T 6438-1992)测定。
相关计算公式如下:
存活率(SR, %)=100×(Nt-N0) /N0;
摄食率(FR, %)=100×I/ [t×(W0+Wt) / 2];
增重率(WGR, %)=100×(Wt-W0) /W0;
特定生长率(SGR, %/d)=100×(lnWt-lnW0) /t;
饲料转化率(FCE, %)=100×(Wt-W0) /I;
肝体指数(HSI, %)=Wh/Wb×100;
脏体指数(VSI, %)=Wv/Wb×100;
肥满度(CF, g/cm3)=Wb/L3×100。
式中:Nt,N0分别为实验结束和实验开始时鱼的尾数;I为总摄食量( g );W0,Wt分别为实验开始和结束时花鲈幼鱼的体重( g );t为实验天数( d );Wh、Wv分别为肝脏重和内脏重( g );Wb为鱼体重( g ),L为鱼体长( cm )。
1.4 统计分析
胃排空实验数据用Excel进行统计和常规计算,用SPSS17.0对数据进行分析、拟合并获得相关参数。养殖实验结果用平均数±标准差(Mean±SD)表示,用Excel对数据进行常规计算与统计,用SPSS17.0统计软件对数据进行单因素方差分析(One-way ANOVA),再用Duncan法进行多重比较检验各组之间差异显著性,以P<0.05为显著性水平。
2 结果
2.1 花鲈幼鱼的胃排空特征
花鲈幼鱼在20 ℃条件下,其胃含物排空变化见图1。由图1可见,花鲈幼鱼瞬时胃含物湿重随摄食后时间的变化呈现一定的变化趋势,其瞬时胃含物湿重随摄食后时间的变化先逐渐降低,后渐渐趋于平缓直至最后变成0。花鲈幼鱼胃含物湿重在饱食后9 h内下降显著,9 h时胃含物湿重降低了约80%,以后下降逐渐缓慢直至达到完全排空的状态。
2.2 花鲈幼鱼胃排空模型的选择
3种数学模型逐一拟合所得实验数据,发现平方根和线性拟合效果均较好,指数拟合效果较差(见图1)。从鱼类最佳胃排空模型评选指标之一的相关系数R2来看,平方根模型最大,指数模型最小,线性模型介于两者之间,而从残差平方和(RSS)及残差标准差(SDR)指标上看均是平方根模型最小,指数模型最大(见表2),与拟合图结果吻合。综合以上相关指标分析得出平方根模型对花鲈幼鱼胃排空变化的描述效果最佳。由平方根模型公式可计算得出,花鲈幼鱼在饱食后17 h左右胃排空,食欲基本恢复至摄食前的状态,80%胃排空时间为9.38 h。
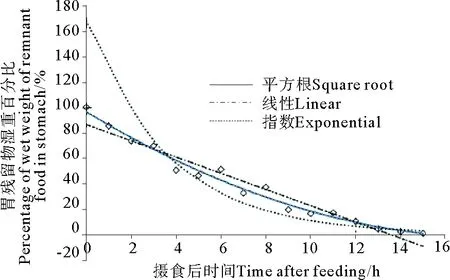
图1 花鲈幼鱼胃排空变化Fig.1 The changes of gastric evacuation of juvenile Lateolabrax maculates

数学模型Model公式EquationR2RSSSDRP线性模型LinearmodelY=86.367-6.383t0.949750.6153.62<0.001指数模型ExponentialmodelY=167.67e-0.273t0.856950.5863.37<0.001平方根模型SquarerootmodelY0.5=9.968-0.586t0.980283.8621.84<0.001
2.3 投喂频率对花鲈幼鱼生长指标的影响
经过5周的养殖实验,花鲈幼鱼终末体重、增重率、摄食率和特定生长率和饲料转化率等指标见表3。由表3可见,从F1组到F3组,花鲈幼鱼的终末体重、增重率、特定生长率均随每天投喂次数的增加而显著升高,且各处理组间差异显著(P<0.05),而F3、F4、F5组间差异均不显著(P>0.05)。除F4和F5组间差异不显著(P>0.05),摄食率在F1到F4组之间呈显著升高的趋势(P<0.05)。饲料转化率在F3组最高,F5组最低,F3组显著高于其他4组(P<0.05),F1组与F2、F5组差异均不显著(P>0.05),而显著低于F3、F4组(P<0.05)。不同投喂频率下,花鲈幼鱼均出现轻微死亡,但各组存活率均在93%以上,且组间差异不显著(P>0.05)。
2.4 投喂频率对花鲈幼鱼形体指标的影响
由表4可知,F5组肝体指数显著高于除F4组的其他各组(P<0.05)。内脏指数随着投喂频率的增加呈逐渐升高的趋势,F5组最高,F1组最低,F5组显著高于除F4组的其他各组(P<0.05),F1到F4组之间无明显差异。肥满度在各不同投喂频率组间差异不显著。
2.5 投喂频率对花鲈幼鱼体组分的影响
不同投喂频率组花鲈幼鱼全鱼营养组分见表5。全鱼水分含量随着投喂频率的增加逐渐降低,F1组水分含量最高,显著高于除F2组外的其他各组(P<0.05),F2、F3、F5组以及F3、F4、F5组之间均无显著性差异(P>0.05)。粗脂肪含量从F1到F4组上升显著(P<0.05),F5与F3、F4组之间差异不显著(P>0.05)。F1、F2组灰分含量显著高于F3、F4和F5(P<0.05),F3、F4和F5组之间无显著性差异(P>0.05)。投喂频率对花鲈幼鱼全鱼粗蛋白含量没有显著性影响(P>0.05)。
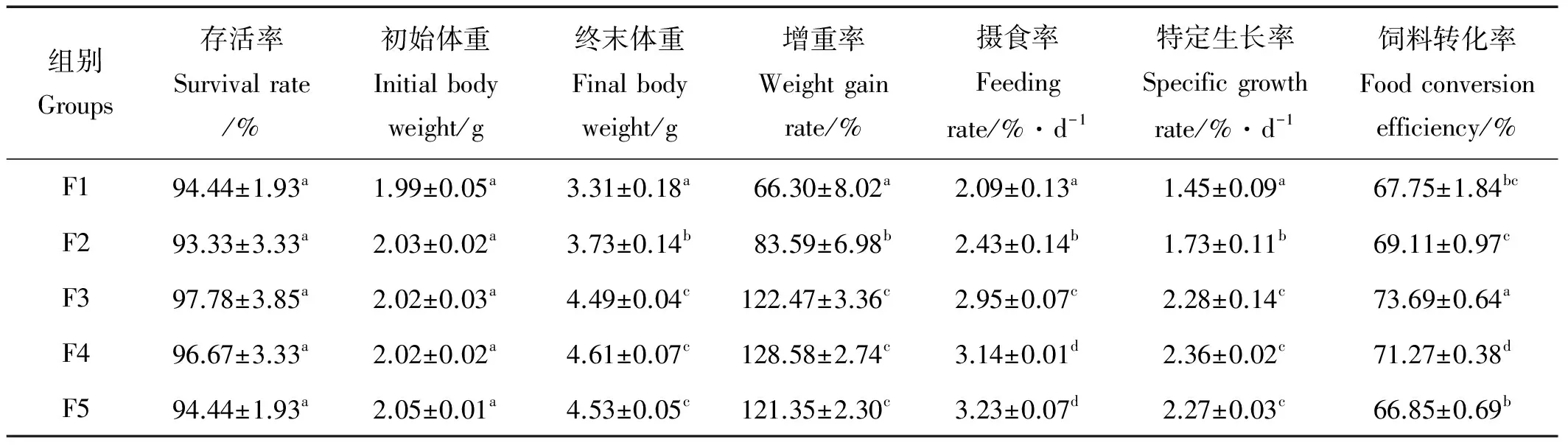
表3 投喂频率对花鲈幼鱼存活率、增重率、特定生长率、摄食率和饵料转化率的影响Table 3 Effects of feeding frequency on survival、weight gain rate (WGR)、specific growth rate (SGR)、feeding rate (FR) and food conversion efficiency (FCE )of Lateolabrax maculatus juvenile
注:同一列中标有不同字母的数值表示差异显著(P<0.05), 下同。
Note: Values within the same column with different letters are significantly different(P<0.05), The same as below.
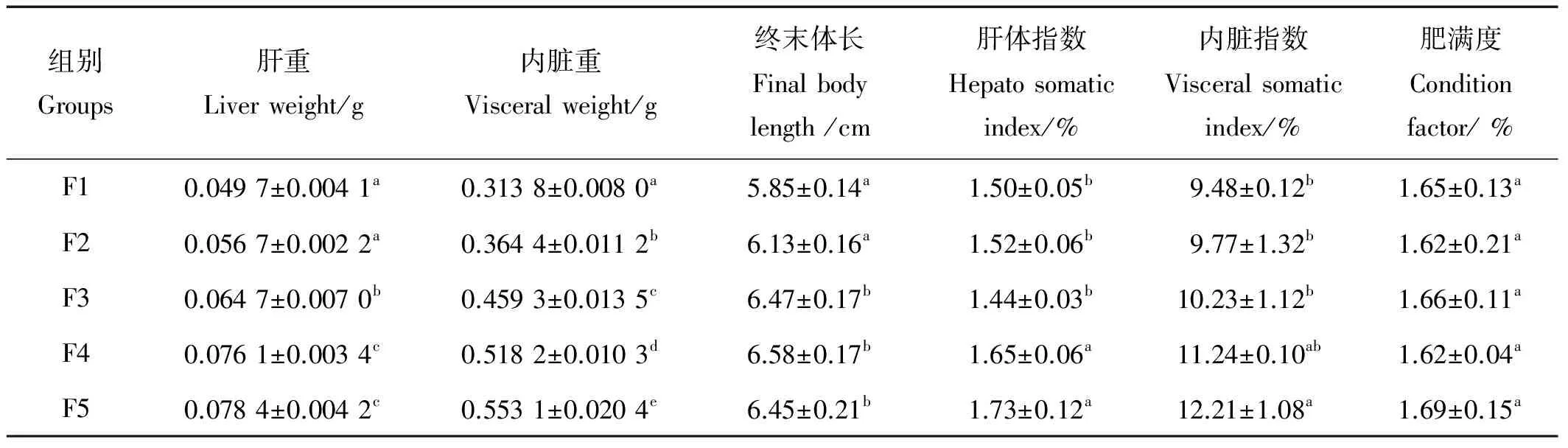
表4 投喂频率对花鲈幼鱼形体指标的影响Table 4 Effects of feeding frequency on body indexes of Lateolabrax maculatus juvenile
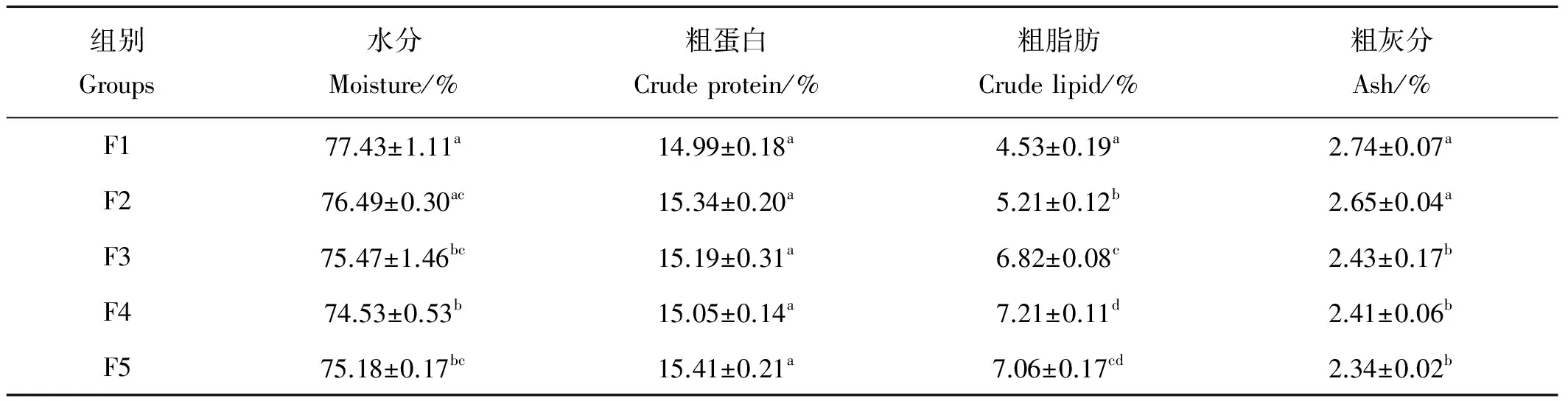
表5 投喂频率对花鲈幼鱼体组分的影响Table 5 Effects of feeding frequency on body composition of Lateolabrax maculatus juvenile
3 讨论
3.1 投喂频率对花鲈幼鱼生长、摄食以及饲料转化率的影响
花鲈幼鱼的摄食率受投喂频率的影响显著。在本实验条件下,从F1组到F4组,花鲈幼鱼的摄食率显著提高,但F4组到F5组,花鲈幼鱼的摄食率虽然继续增加但差异不显著。这与龙虎斑(Hulonggrouper)[12]、条石鲷(Oplegnathusfasciatus)[13]等研究结果相一致。可能与不同投喂频率条件下花鲈幼鱼每天摄食总量不同有关,当其摄食至饱食状态时,继续增加投喂,摄食率无明显提高。
本研究条件下,花鲈幼鱼的终末体重、增重率和特定生长率均随投喂频率的增加不断上升,到F5略有下降,但与F3、F4组间差异不显著。Wu等[14]对金鲳鱼(Trachinotusovatus)、冒树泉等[15]对许氏平鮋(Sebastesschlegeli)幼鱼研究均发现其增重率变化规律与本研究结果非常相似。陈文霞等[16]对凡纳滨对虾(Litopenaeusvannamei)研究发现每天投喂4次的凡纳滨对虾的生长性能显著高于每天投喂2次和3次。这可能是因为投喂频率过低时,鱼类摄食不足,无法满足正常的生长需求,从而导致其生长速度滞后。何利君等[17]研究发现随着投喂频率的增加,南方鲇(Silurusmeridionalis)的特定生长率逐渐增加,但是每天投喂3次的特定生长率较2次时出现下降,这与本实验结论相似,说明适当增加饱食投喂状态下的投喂次数有利于鱼类的生长,但这种频率一旦超过一定的限度,也会导致鱼类因过度消耗能量去摄食从而影响其生长,同时也会增加饲料成本。
部分鱼类的饲料转化率随投喂频率的增加而逐渐降低[15,18]。而有些鱼类在一定范围内其饲料转化率随投喂频率的增加而升高[16,19],但当投喂频率进一步增加时,有些开始下降[20-21],有些则基本保持不变[22]。而何利君等[17]对南方鲇的研究发现,投喂频率对其饲料转化率并无显著性影响,Gwynnc等[23]对大西洋鳙鲽(Hippoglossushippoglossus)的研究结果同南方鲇一样。奥尼罗非鱼(Oreochromiscoaureus)仔稚鱼的饲料转化率随投喂频率的增加先显著降低,而后保持相对恒定[24]。本研究中,花鲈幼鱼的饲料转化率随投喂频率的增加先显著升高,在F3组达到最大,为73.69%,而后逐渐降低,这与斑点叉尾鮰(Ictaluruspunctatus)[21]和黄鳝(Monopterusalbus)[20]的研究结果较一致。原因可能是由于投喂过于频繁会使花鲈胃肠中摄食的食物未经完全消化即被排出体外,影响了其消化吸收;另一方面,也可能是因为在较高投喂频率下花鲈忙于摄食和消化致使分配用于生长的能量占比降低,食物转化率也随之下降。鱼类最适宜的投喂频率因其种类、大小以及饲料营养成分的不同而存在差异。在本实验条件下研究得出,对于2~4.5 g左右的花鲈幼鱼每天投喂2次其生长性能较每天投喂3次组差异不显著,但节省了饲料和劳动力,而且此时的饲料转化率最高。因此,综合养殖成本和生长性能,每天投喂2次是较优的选择。
3.2 投喂频率对花鲈幼鱼形体指标以及体组分的影响
形体指标作为鱼体能量状态的衡量标准在鱼类研究中占有重要的地位[25]。在本研究中,随投喂频率的增加,肝脏指数和内脏指数均升高,这应该是由于摄食量升高导致更多营养物质在肝脏和内脏积累或沉积的缘故[26-27]。而孙丽慧等[28]对星斑川鲽(Platichthysstellatus)幼鱼研究却与本研究结果截然相反,这可能是因为不同的鱼类之间存在差异,也可能与鱼的大小有关。鱼体脂肪含量一般受到诸如养殖水体环境、饲料种类、投喂频率等外界条件的影响。本实验研究结果表明,随着日投喂次数的增加,花鲈幼鱼全鱼水分含量逐渐下降,而脂肪含量则上升显著(P<0.05)。这与大黄鱼(Pseudosciaenacrocea)[29]、尼罗系吉富罗非鱼(Oreochromisniloticus)[7]的研究结果相一致。另一方面,本研究中花鲈幼鱼全鱼脂肪含量的上升可能是因为在高投喂频率下,摄食量的增加使得营养物质以体脂的形式储存[30]。在鱼苗培育过程中,可以适当的提高投喂频率为其越冬储备脂肪;而在成鱼养殖生产上,也可通过改变投喂频率来调控鱼体脂肪含量,进而改善鱼肉品质。
在许多研究中发现,鱼体营养组分中粗蛋白的含量受投喂频率影响不显著[31]。在本研究中也得出了类似的结果。Jarboe等[32]发现,斑点叉尾鮰鱼体灰分含量随投喂频率的增加而下降,与本实验研究结果一致。楼宝等[8]对体重85.95 g花鲈幼鱼不同投喂频率的研究结果表明,水分和蛋白质含量基本不受投喂频率的影响,而粗脂肪含量则随投喂频率的增加明显降低,灰分含量的变化规律则与粗脂肪相反。这一研究结果与本实验结果有所差异,这可能与实验鱼的规格、养殖方式、饵料成分以及环境的不同有关,具体还有待进一步研究论证。
3.3 花鲈幼鱼胃排空特征以及胃排空时间对花鲈幼鱼摄食的影响
不同鱼类因其食性和消化道结构存在差异,胃排空的方式也必然复杂多样。关于其最佳胃排空模型的选择也自然十分困难。即使同种鱼类在不同实验条件下也可能得到不同的数学模型。目前研究中最经常使用的是指数、线性和平方根这3种模型。已经发现许多以浮游生物为食的小型海洋鱼类是以指数模型排空的,这种排空的特征是先快后慢,如赤鼻棱鳀 (Thryssakammalensis)、玉筋鱼 (Ammodytespersonatus) 、斑鰶(Clupanodonpunctatus)和小鳞鱵(Hemirhamphussajori) 这4种鱼类[33];许多肉食性鱼类是以线性模型排空的,其特征是直线下降,如眼斑拟石首鱼(Sciaenopsocellatus)[3]、大眼梭鲈(Stizostedionvitreum)[34]等;而平方根模型的排空特征是刚开时速度较慢,然后速度加快,最后又开始减慢。如南方鲇[4]、俄罗斯鲟(Acipensergueldenstaedtii)幼鱼[2]等。在本研究条件下,对花鲈幼鱼胃排空实测数据分析得出,平方根模型最适合定量描述花鲈幼鱼胃排空变化。
鱼类胃排空速率很大程度上与其食欲的恢复有关,胃排空时间对鱼类最佳投喂频率的确定具有重要的指导意义[7, 19]。Grove等[35]对虹鳟(Oncorhynchusmykiss)研究发现,其胃内残余食物湿重占比低于1/2时,食欲基本恢复到摄食前水平的一半。一般情况下我们把鱼类胃内残余饲料量为零的时间点作为其食欲完全恢复至摄食前水平的标志。也有研究认为,对于一些胃容量较大的鱼类,其胃排空超过80%,即可认定其食欲基本得到恢复[6]。由平方根模型公式Y0.5=9.968-0.586t可得出本实验条件下花鲈幼鱼80%和99%胃排空时间分别为9.38和15.3 h,理论上投喂间隔9.38 h之后,花鲈幼鱼食欲基本得到恢复。投喂频率为4次/d、3次/d时,其投喂时间间隔分别为3.5和5 h,在这期间花鲈幼鱼胃排空程度约为摄食前的37%和50%,此时花鲈食欲尚未完全恢复,再进行投喂必然导致饲料利用效率的降低。这与Gwyther对黄盖鲽(Limandalimanda)研究结果一致[36]。投喂频率为1次/2 d、1次/d,其投喂时间间隔已经达到或超过24 h,花鲈幼鱼胃过度排空,已经处于饥饿状态,这必然伴随着生长性能的降低。投喂频率为2次/d时,其投喂时间间隔为10 h,花鲈幼鱼胃排空超过了80%,食欲基本恢复,特定生长率和食物转化率此时应处于较高水平,生长性能也明显优于其他各组,这与养殖实验结论相符。孙晓峰等[7]研究得出,4.2 g左右的尼罗系吉富罗非鱼幼鱼80% 胃排空为9 h,养殖实验结果同时发现,投喂频率为2次/d (投喂时间间隔为9 h) 其特定生长率较高,饲料转化率最高,此时的鱼种表现出较佳的生长性能,这与本实验对花鲈幼鱼研究结论相似。
4 结论
本文以花鲈幼鱼为研究对象,采用传统养殖实验的方法,设计了1次/2 d、1次/d、2次/d、3次/d、4次/d 5个日投喂频率组,对(2.75±0.27)g花鲈幼鱼在养殖期间的生长性能进行了比较研究。结果表明,此阶段花鲈幼鱼在每天投喂2次时生长快、饲料转化率最高,而且养殖成本大大降低;胃排空实验表明的每天投喂2次的投喂时间间隔有助于花鲈幼鱼对饲料进行充分;综合上述结果,每天投喂2次是适宜花鲈幼鱼生长的最佳日投喂频率。
参考文献:
[1] Lee S, Hwang U, Cho S H. Effects of feeding frequency and dietary moisture content on growth, body composition and gastric evacuation of juvenile Korean rockfish(Sebastesschlegeli)[J]. Aquaculture, 2000, 187(3): 399-409.
[2] 张涛, 李伟杰, 庄平, 等. 俄罗斯鲟幼鱼的胃排空特征及其数学模型[J]. 海洋渔业, 2015, 37(2): 151-155.
Zhang T, Li W J, Zhuang P, et al. Analysis of gastric evacuation characteristics and associated mathematical models in juvenileAcipensergueldenstaedtii[J]. Marine Fisheries, 2015, 37(2): 151-155.
[3] 余方平, 许文军, 薛利建, 等. 美国红鱼的胃排空率[J]. 海洋渔业, 2007, 29(1): 49-52.
Yu F P, Xu W J, Xue L J, et al. The gastric evacuation rate of Sciaenopsocellatus[J]. Marine Fisheries, 2007, 29(1): 49-52.
[4] 曾令清, 李凤杰, 曹振东, 等. 南方鲇幼鱼的胃排空特征及其数学模型[J]. 水产学报, 2011, 35(2): 231-237.
Zeng L Q, Li F J, Cao Z D, et al. The analysis of gastric evacuation characteristics and associated mathematical models in juvenile southern catfish (Silurusmeridionalis)[J]. Journal of Fisheries of China, 2011, 35(2): 231-237.
[5] Jobling M. Mathematical models of gastric emptying and the estimation of daily rates of food consumption for fish [J]. Journal of Fish Biology, 1981, 19(3): 245-257.
[6] Riche M, Haley D I, Oetker M, et al. Effect of feeding frequency on gastric evacuation and the return of appetite in tilapiaOreochromisniloticus( L.) [J]. Aquaculture, 2004, 234(1-4): 657-673.
[7] 孙晓锋, 冯 健, 陈江虹, 等. 投喂频率对尼罗系吉富罗非鱼幼鱼胃排空、生长性能和体组成的影响[J].水产学报, 2011, 35(11): 1677-1683.
Sun X F, Feng J, Chen J H, et al. Effects of feeding frequency on gastric evacuation, growth benefit and body composition of juvenile genetic improved farmed tilapia strain of Nile tilapia(Oreochromisniloticus)[J]. Journal of Fisheries of China, 2011, 35(11): 1677-1683.
[8] 楼宝, 史会来, 毛国民, 等. 饲喂频率对鲈鱼生长及体生化成分的影响[J]. 宁波大学学报(理工版), 2007, 20(4): 455-458.
Lou B, Shi H L, Mao G M, et al. Effects of feeding frequency on growth and body biochemical composition ofLateolabraxjaponicus[J]. Journal of NingBo University(NSEE), 2007, 20(4): 455-458.
[9] 竺俊全, 李明云, 吴锡科. 花鲈仔稚鱼的生长发育与摄食节律研究[J]. 浙江海洋学院学报(自然科学版), 2002, 21(3): 210-215.
Zhu J Q, Li M Y, Wu X K. Study on the growth and feeding rhythm of larval and juvenile ofLateolabraxjaponicus[J]. Journal of Zhejiang Ocean University (Natural Science), 2002, 21(3): 210-215.
[10] 肖雨, 刘红. 花鲈鱼种日摄食节律的初步研究[J]. 水产科技情报, 1997(3): 99-103.
Xiao Y, Liu H. Preliminary study on daily feeding rhythm ofLateolabraxjaponicusfingerlings[J]. Fisheries Science & Technology Information, 1997(3): 99-103.
[11] 王晓龙, 温海深, 张美昭, 等. 花鲈初孵仔鱼饥饿不可逆点的确定及摄食节律研究[J]. 中国海洋大学学报(自然科学版), 2017(5): 57-64.
Wang X L, Wen H S, Zhang M Z, et al. Determination of irreversible starvation point and feeding rhythm ofLateolabraxmaculatuslarvae[J]. Periodical of Ocean University of China (Natural Science), 2017(5): 57-64.
[12] 吴永丽, 黄建彪, 马细兰, 等. 投喂频率对虎龙斑摄食、生长和饲料转化率的影响[J]. 安徽农业科学, 2016, 44(27): 108-109.
Wu Y L, Huang J B, Ma X L, et al. Effects of feeding frequency on feeding, growth and feed conversion rate ofHulonggrouper[J]. Journal of Anhui Agriculture Science, 2016, 44(27): 108-109.
[13] Oh S Y, Maran B A V. Feeding frequency influences growth, feed consumption and body composition of juvenile rock bream (Oplegnathusfasciatus)[J]. Aquaculture International, 2015, 23(1): 175-184.
[14] Wu Y, Han H, Qin J, et al. Effect of feeding frequency on growth, feed utilization, body composition and waste output of juvenile golden pompano (Trachinotusovatus) reared in net pens[J]. Aquaculture Research, 2015, 46(6): 1436-1443.
[15] 冒树泉, 邹明妤, 王春生, 等. 许氏平鲉幼鱼适宜投喂频率的研究[J]. 动物营养学报, 2014, 26(8): 2379-2385.
Mao S Q, Zou M Y, Wang C S, et al. Optimal feeding frequency forSebastesschlegeliJuveniles[J]. Chinese Journal of Animal Nutrition, 2014, 26(8): 2379-2385.
[16] 陈文霞, 申玉春, 李再亮, 等. 投喂频率对凡纳滨对虾生长、消化酶和免疫酶活力以及氮收支的影响[J]. 海洋科学, 2013, 37(9): 49-53.
Chen W X, Shen Y C, Li Z L, et al. The effect of feeding frequency on growth, digestive enzymes, immune enzyme and N budget ofLitopenaeusvannamei[J]. Marine Sciences, 2013, 37(9): 49-53.
[17] 何利君, 谢小军, 艾庆辉. 饲喂频率对南方鲇的摄食率、生长和饲料转化效率的影响[J]. 水生生物学报, 2003, 27(4): 434-436.
He L J, Xie X J, Ai Q H. Effects of feeding frequency on feeding rate, growth and feed conversion efficiency for the southern catfish,SilurusMeridionalis[J]. Journal of Aquatic Biology, 2003, 27(4): 434-436.
[18] Liu F G, Liao C I. Effect of feeding regimen on the food consumption, growth and body composition in hybrid striped bassMoronesaxitilis×M.chrysops[J]. Fisheries Science, 1999, 64(4): 513-519.
[19] 覃志彪, 赵海祥, 赵华林, 等. 投饲频率对瓦氏黄颡鱼幼鱼胃排空、生长效益及体组成影响[J]. 海洋与湖沼, 2011, 42(4): 506-511.
Qin Z B, Zhao H X, Zhao H L, et al. Effects of feeding frequency on gastric evacuation, growth performance and body composition of juvenile darkbarbel catfish (PelteobagrusvachelliRichardson)[J]. Oceanologia Et Limnologia Sinica, 2011, 42(4): 506-511.
[20] 杨帆, 张世萍, 韩凯佳, 等. 投喂频率对黄鳝幼鱼摄食、生长及饵料利用效率的影响[J]. 淡水渔业, 2011, 41(3): 50-54, 82.
Yang F, Zhang S P, Han K J, et al. Impact of feeding frequency on feeding, growth and feed utilization for juvenileMonopterusalbus[J]. Freshwater Fisheries, 2011, 41(3): 50-54, 82.
[21] 董桂芳, 胡振雄, 黄峰, 等. 投喂频率对斑点叉尾鮰幼鱼生长、饲料利用和鱼体组成的影响[J]. 渔业现代化, 2012, 39(2): 48-53.
Dong G F, Hu Z X, Huang F, et al. Effect of feeding frequency on growth, feed utilization and whole body composition of juvenile channel catfish (Ictaluruspunctatus)[J]. Fishery Modernization, 2012, 39(2): 48-53.
[22] 席寅峰, 张东, 施兆鸿. 投喂频率对雌雄分化后灰海马生长发育、饵料转换效率及消化酶活力的影响[J].海洋渔业, 2013, 35(1): 77-85.
Xi Y F, Zhang D, Shi Z H. Effects of feeding frequency on the growth, food efficiency and digestive enzyme activity ofHippocampuserectusafter sex differentiation[J]. Marine Fisheries, 2013, 35(1): 77-85.
[23] Schnaittacher G, King W V, Berlinsky D L. The effects of feeding frequency on growth of juvenile Atlantic halibut,HippoglossushippoglossusL.[J]. Aquaculture Research, 2015, 36(4): 370-377.
[24] 强俊, 王辉, 李瑞伟, 等. 饲喂频率对奥尼罗非鱼仔稚鱼生长、体成分和消化酶活力的影响[J]. 广东海洋大学学报, 2009, 29(4): 79-83.
Qiang J, Wang H, Li R W, et al. Effects of feeding frequency on growth, body biochemical composition and digestive enzymes of larvae and juvenile of hybrid tilapia(Oreochromisniloticus×O.aureus)[J]. Journal of Guangdong Ocean University, 2009, 29(4): 79-83.
[25] Daudpota A M, Abbas G, Kalhoro I B, et al. Effect of feeding frequency on growth performance, feed utilization and body composition of juvenile Nile tilapia,Oreochromisniloticus(L.) reared in low salinity water[J]. Pakistan Journal of Zoology, 2016, 48(1): 171-177.
[26] Azaza M S, Khiari N, Dhraief M N, et al. Growth performance, oxidative stress indices and hepatic carbohydrate metabolic enzymes activities of juvenile Nile tilapia,OreochromisniloticusL., in response to dietary starch to protein ratios[J]. Aquaculture Research, 2015, 46(1): 14-27.
[27] 乐贻荣, 杨弘, 徐起群, 等. 饲料蛋白水平对奥尼罗非鱼(Oreochromisniloticus×O.aureus)生长、 免疫功能以及抗病力的影响[J]. 海洋与湖沼, 2013, 44(2): 493-498.
Yue Y R, Yang H, Xu Q Q, et al. Effects of dietary protein level on growth performance, immunity function and disease resistance for hybrid tilapia(Oreochromisniloticus×O.Aureus)[J]. Oceanologia Et Limnologia Sinica, 2013, 44(2): 493-498.
[28] 孙丽慧, 王际英, 丁立云, 等. 投喂频率对星斑川鲽幼鱼生长和体组成影响的初步研究[J]. 上海海洋大学学报, 2010, 19(2): 190-195.
Sun L H, Wang J Y, Ding L Y, et al. Effects of feeding frequency on growth and body composition of juvenilePlatichthysstellatus[J]. Journal of Shanghai Ocean University, 2010, 19(2): 190-195.
[29] 孙瑞健, 张文兵, 徐 玮, 等. 饲料蛋白水平与投喂频率对大黄鱼生长、体组成及蛋白质代谢的影响[J]. 水生生物学报, 2013, 37(2): 281-289.
Sun R J, Zhang W B, Xu W, et al. Effects of dietary protein level and feeding frequency on the growth performance, body composition and protein metabolism of juvenile large yellow croakers,PseudosciaenaCroceaR.[J]. Journal of Aquatic Biology, 2013, 37(2): 281-289.
[30] Cho S H, Lim Y S, Lee J H, et al. Effects of feeding rate and feeding frequency on survival, growth and body composition of Ayu post-larvaePlecoglossusaltivelis[J]. Journal of the World Aquaculture Society, 2003, 34: 85-91.
[31] Baloi M, Carvalho C V A, Sterzelecki F C, et al. Effects of feeding frequency on growth, feed efficiency and body composition of juveniles Brazilian sardine,Sardinellabrasiliensis(Steindacher 1879)[J]. Aquaculture Research, 2016, 47(2): 554-560.
[32] Jarboe H H, Grant W J. Effects of feeding time and frequency on growth of channel catfishIctaluruspunctatusin closed recirculating raceway systems[J]. Journal of The World Aquaculture Society, 1996, 27(2): 235-239.
[33] 孙耀, 刘勇, 张波, 等. 渤、黄海4种小型鱼类摄食排空率的研究[J]. 海洋与湖沼, 2002, 33(6): 679-684.
Sun Y, Liu Y, Zhang B, et al. Gastric evacuation rates of 4 small-size fish species in BoHai and Yellow seas[J]. Oceanologia Et Limnologia Sinica, 2002, 33(6): 679-684.
[34] Swenson W A, Smith L L. Gastric digestion, food consumption, feeding periodicity and food conversion efficiency in walleye,Stizostedionvitreumvitreum[J]. Journal of The Fisheries Research Board of Canada, 1973, 30(9): 1327-1336.
[35] Grove D J, Loizides L G, Nott J. Satiation amount, frequency of feeding gastric emptying rate inSalmogairdneri[J]. Journal of Fish Biology, 1978, 12(5): 507-516.
[36] Gwyther D, Grove D J. Gastric emptying inLimandalimanda(L.) and the return of appetite[J]. Journal of Fish Biology, 1981, 18(2): 245-259.
