东拉山短丝木犀植物群落土壤养分对其群落空间分布的影响
2018-04-08杨国栋钱慧蓉王贤荣
杨国栋,钱慧蓉,陈 林*,王贤荣
(1 南京林业大学 南方现代林业协同创新中心, 南京 210037;2 南京林业大学 生物与环境学院, 南京 210037)
植被与环境因子之间的关系向来是生态学领域研究的热点[1-2],在诸多环境因子中,土壤作为最重要的水分及养分资源的提供者,对植物的生长和繁殖有重要作用[3],也可以通过调节资源的供给与分配影响生态系统内的物种组成和群落动态, 制约生态系统的演替过程和对环境变化的响应[4]。良好的土壤功能是维持植被群落生态系统稳定的基础,其中的土壤有机质、氮素、磷素和钾素不仅是土壤组成的重要部分,而且是生态系统中极其重要的生态因子[5]。自然条件下的植被群落结构是物种对环境长期适应的结果[6],不同生态适应类型、不同层次的植被对土壤环境响应程度的差异性,导致植被群落结构随土壤功能区分表现出分异格局[7]。研究植被与土壤环境之间的相互作用关系,通过分析其空间分布格局与植被群落演替规律,进而为制定物种多样性保护对策、有效利用植物资源提供科学依据[8]。
短丝木犀(Osmanthusserrulatus)是木犀科(Oleaceae)木犀属(Osmanthus)野生种之一,是中国特有树种,其模式产地即为四川宝兴县。该植物的模式标本由英国植物采集家E. H. Wilson于1910年在四川穆坪(今宝兴)采得,随后由美国植物学家A. Rehder鉴定为一新种,于1916年发表在《Plantae Wilsonianae》一书中[9]。中国植物志曾记载其在四川、广西、福建有分布,但近年来仅在四川的西岭雪山、峨眉山、东拉山等地有所发现,其中东拉山谷集中分布于海拔1 800~2 300 m的短丝木犀种群群落是迄今为止世界上发现的面积最大的自然种群。短丝木犀是木犀属少数春季开花的种类之一,花大而芬芳,花梗长而纤细,极为美丽[10]。20世纪初Wilson等采集到该植物的种子之后,英国皇家植物园,Hiller树木园等处已引种栽培,如今已长成大树,在英国、欧美等国家在园林中已有应用,可是在中国该植物却尚未开发利用。短丝木犀春季开花,有香味,具独特的观赏价值和开发利用前景[11]。
东拉山谷地处四川盆地向青藏高原的过渡地带,其连绵的山脉、纵横的沟谷、蜿蜒的溪流等复杂的地形地貌和湿润多雨的自然气候以及相对封闭的原始自然环境造就了当地极为丰富的动植物多样性,其间的赶羊沟也是最早一批发现野生大熊猫的地方。该区有着“物种避难所”的环境特点,在大尺度上气候对植被分布起着决定性作用,在较小尺度上地形、土壤、生物之间的相互作用等因素也影响着植物群落的分布。本研究对短丝木犀群落的空间分布及其与土壤肥力之间的关系进行分析,以期明确该植被群落演替过程中土壤因子的变化机制,并对短丝木犀群落的空间分布及其与土壤养分含量的关系进行分析,并定量分离各类因素对短丝木犀群落空间分布的解释能力,目的在于为了片段破碎化的短丝木犀群落的保护与恢复,为中国野生木犀属种质资源的保护和利用提供科学依据。
1 材料和方法
1.1 研究区概况
东拉山大峡谷位于龙门山脉邛崃山脉南部的宝兴段,地处四川省雅安市宝兴县陇东镇境内,位于E102.25°~102.95°,N30.21°~30.69°,总面积约348 km2,海拔1 800~5 338 m,集典型的峡谷地貌和雪山草原等自然生态景观于一体。该区气候属亚热带季风气候区,由于受山地海拔影响,垂直变化明显,具有亚热带到永冻带的垂直气候[12]。全境褶皱密集,断裂发育,天然植被保存完好,植被类型为亚热带常绿阔叶林,植被类型有常绿阔叶林、常绿和落叶阔叶混交林、落叶阔叶林、针阔混交林等[13]。
1.2 研究方法
1.2.1样地选择2016年3月在沿短丝木犀分布线路踏查的基础上,兼顾物种群落生境地分布的不同,选择具有代表性、典型性的集中分布地段共设12个20 m×20 m的正方形样地(图1),每个样地内用插值法细分为4个10 m×10 m与16个5×5的小样方。样地海拔在1 950~2 100 m,西南坡,坡度为38°。采用交叉五点取样法,以1 m×1 m区域为网格单元,对240个采样点进行调查采样并GPS定位。

图1 短丝木犀群落的12个样点Fig.1 12 samples community of Osmanthus serrulatus
1.2.2植物调查在12个短丝木犀样方中,采用“每木记帐法”记录群落中所有物种的信息,包括:(1)胸径(DBH)2 cm以上的乔木种类的种名、胸径、高度和冠幅,测定其坐标;(2)胸径小于2 cm的灌木种类的种名、平均高度和冠幅;(3)草本植物的种数、多度和平均高度。
1.2.3物种多样性计算方法以物种数(S)作为衡量物种丰富度的参数。
植物重要值计算:重要值(IV) =(相对基盖度+相对频度+相对密度)/3
其中相对基盖度(RCO) =Ci/∑Ci× 100 (Ci表示样方中某植物种基盖度,即植物的胸高断面积,m2);相对频度(RFE) =Fi/∑Fi× 100 (Fi表示样方中某植物种频度,%);相对密度(RDE) =Di/∑Di× 100(Di表示样方中某植物密度,株/m2)
Simpson指数和Berger-Parker指数均为优势度指数,其中前者随一个或几个物种的优势度的增加而增加,后者完全由最大相对多度的物种给出。Shannon-Wiener指数为变化度指数,物种数量越多,其分布越均匀,其值越大。本文采用3个定量指标即Simpson指数、Shannon-Wiener指数和在Shannon-Wiener指数基础上的Pielou均匀度指数分布来测度和分析群落物种多样性与环境的关系[14]。
1.2.4土壤样品处理和分析土壤样品分析方法依据《土壤农化分析方法》[15],在每个样方网格内,用环刀对0~20 cm的土壤进行3点重复取样,混合后密封,带回实验室测定,所有样品经风干、剔除杂质、研磨和过筛后,用于测定沉积物的全氮、全磷和全钾。土壤有机质测定采用浓硫酸-重铬酸钾外加热法,土壤全氮采用浓硫酸消煮-凯氏定氮法,土壤全磷采用浓硫酸-高氯酸消煮钼锑抗比色法,全钾含量采用碱熔-火焰光度法测定。
1.2.5数据处理采用克里格(Kriging)插值法,利用原始数据和半方差函数的结构性,在有限区域内对区域化变化量进行无偏最优估计[16-17],不仅能够将有数据的采样点之间的自相关性量化,还可以预测估算采样点在未知区域范围内的空间如何分布。
式中,Z(xi)为没有数据的采样点四周的已经有数据的采样点的值;λi为第i个已经知道数据的采样点对没有数据的采样点的权重。
地统计中往往要求数据呈正态分布,否则会出现比例效应,对分析结果造成较大误差。在ArcGIS地统计分析模块中,可利用探索性分析模块中的QQPlot图对土壤养分含量数据分布进行初步判断和说明。QQPlot图在直观的对正态分布情况进行表达时,需结合SPSS中K-S检验得出的渐进显著性,即P值,对数据的分布进行正态分布的最终判断,当P<0.05时,数据不符合正态分布,对于不符合正态分布的需要对数据进行相关转换,反之,则符合正态分布。
1.2.6数据源的建立将最终240个采样点的GPS数据、土壤样品养分测定数据等汇总录入Excel,编辑好各样点属性数据,并导入ArcGIS中,建立样点属性数据库。
2 结果与分析
2.1 植物群落结构特征
根据短丝木犀的样方资料统计,4 800 m2的样地中共有维管束植物235种,隶属于77 科143 属,其中蕨类植物8科9属12种,分别占科、属、种总数的10.39%、6.29% 和5.08% 。裸子植物2科5属9种,被子植物67 科129 属214种,占科、属、种总数的87.01%、87.76%和91.06%。
短丝木犀群落镶嵌在常绿和落叶阔叶混交林、落叶阔叶林或针阔混交林中,群落组成种类丰富,科、属组成复杂。该群落主要由常绿和落叶种类混合组成,群落外貌有明显的季相变化。由于海拔较高,气候条件相对较为严峻,群落物种个体较小,群落结构可分为乔木层、灌木层、草本层。乔木层分层明显,共有75种植物,以短丝木犀、云锦杜鹃(Rhododendronfortunei)、猫儿刺(Ilexpernyi)和鹅耳栎(Carpinusturczaninowii)为优势种。灌木层主要由乔木层的幼树、幼苗和其它灌木组成,平均高度0.95 m,共有93种。草本层植物共有70种,其中数量较多的有大叶水冷花(Pileamartinii)、虎耳草(Saxifragastolonifera)、紫萁(Osmundajaponica)和尼泊尔蓼(Polygonumnepalense)等。
群落乔木层和灌木层的物种重要值统计,可以很好地体现主要物种在群落中的地位和作用。根据调查统计,乔木层中重要值在3.0以上的总共有16种(表1),其中短丝木犀和云锦杜鹃的重要值分别为26.43和23.66,在群落中占有较为明显的优势地位,其次为猫儿刺,重要值为11.93,再次为鹅耳栎,重要值为9.75。

表1 东拉山短丝木犀群落乔木层主要树种的特征值

表2 东拉山短丝木犀群落植被多样性指数
注:D.Simpson多样性指数;H.Shannon多样性指数;J.Pielou均度指数;W.Whittaker多样性指数;Jh.Hill多样性指数;F.Alatalo均度指数;G.Molinari均度指数; C.优势度指数;h1.修正的Simpson数;d1.种的丰富度指数;Mc.McIntosh多样性指数
Note: D. Simpson diversity index; H. Shannon diversity index; J. Pielou evenness index; W. Whittaker diversity index; Jh. Hill diversity index; F. Alatalo evenness index; G. Molinari evenness index; C. Dominance index; h1. Modified Simpson number; d1. Richness index of the species; Mc. McIntosh diversity index
样地的3个定量指标即Simpson指数(0.924 1)、Whittaker多样性指数指数(2.221 2)和在Shannon-Wiener指数基础上的Pielou均匀度指数(0.924 7),用来测度和分析东拉山短丝木犀群落物种多样性与环境的关系(表2)。不同群落或某环境梯度上不同点在于优势种的不同,该样地的主要坡向为西南-东北走向,西南坡较之东北坡全年日照时间要长,其物种分布与组成自然存在着差异。挑选群落样地里代表性的4种植物分布图:以短丝木犀和云锦杜鹃为代表的山地常绿阔叶林;以鹅耳枥为代表的山地落叶阔叶林;以铁杉为主的山地针叶林(图2)。短丝木犀主要密集分布方位在东南方位,该群落所处环境较为恶劣,植株生长较为缓慢,实际调查中也发现成年个体相对较多;铁杉分布地都在群落样地的北部,该区海拔较高,地形陡峭;阔叶林散布在群落样地内,分布广泛。乔木层和灌木层的建群种都是短丝木犀,生长旺盛,占据了极大的生存资源空间,优势度明显。
2.2 群落土壤元素含量的统计特征
利用ArcGIS地统计模块对东拉山短丝木犀群落土壤养分含量进行数据分析生成直方图,研究区群落土壤中,全氮、全磷、全钾和有机质含量的经典描述性统计见图3。4个指标含量变化范围分别为6.4~12.5、1.04~1.25、23.5~29.4、211~318 g/kg;最大值分别是最小值的1.94倍、1.21倍、1.25和1.51倍,差异最大的是全氮。几项指标平均值和中值相差不大。从变异系数来看,在土壤成分的空间分布研究中,通常使用变异系数CV表示土壤相关成分含量的空间离散或者变异程度,通常分为三级标准:CV≤10%时,数据间存在空间弱变异性;CV≥100%时,为空间强变异性;当CV介于两者之间时为空间中等变异性[18]。全氮、全磷、全钾和有机质均表现为中等程度的变异性。短丝木犀群落的表层土壤元素的平均值与中值基本相等,说明土壤元素含量分布中心趋于正态分布。

图2 样地内4种指示树种分布图Fig.2 The distribution of four kinds of indicator trees in the community

图3 东拉山短丝木犀群落土壤4种养分含量直方图Fig.3 Histogram of four nutrient contents in Dongla Mountains community soil

名称Name范围Range程度Degree描述Describe峰度Kurt3<峰度≤数据个数3
峰度系数(表3)用来度量数据在中心的聚集程度。全氮、全磷和有机质含量分布的峰度系数都小于3,平均值近似中值,说明三者含量分布与正态分布特点相似,三者含量分布曲线中心比正态分布较窄较缓;全钾含量分布的峰度系数大于3,说明分布曲线中心比正态分布较窄较陡。表层土壤元素的含量分布的偏度系数都小于0,则可判断4种养分含量分布成负偏态,在直方图中也可明显看出此特征。在表层土壤元素趋势的分析过程当中,透视面的选择应尽量使采样数据在透视面投影点分布集中,这样通过投影点拟合的趋势方程具有代表性,能有效反映全局趋势。如果拟合曲线平直,则没有全局趋势;如果拟合的曲线为曲线,则存在全局趋势[19]。
2.3 群落土壤元素含量空间分布趋势分析
利用ArcGIS软件中的地统计模块分析东拉山短丝木犀群落的表层土壤含量的趋势(图4),其中每条线的高度代表此点位处的土壤元素含量值,每条线的位置为采样点在东西和南北方向上的投影位置。结果显示,4种养分含量的蓝线和绿线都呈微弱的U型趋势,说明表层土壤养分含量的在东西和南北两方向上都有变化。全钾的蓝线在由北至南的方向上呈逐渐降低的趋势,而且趋势较明显,有较明显的U形趋势,说明东拉山短丝木犀群落的全钾含量高于南部区域;绿线由西至东的方向上呈先上升后下降的趋势,趋势不明显,说明群落的东部与西部总体含量差异不明显。
群落内全氮、全磷和有机质的含量蓝线和绿线都呈微弱的U型趋势,说明三者含量的在东西和南北两方向上都有变化,且在南北方向上,呈两头高,中间低的特点,即先升高后降低的趋势;在东西方向上呈两头低的特点,即中间高的先降低后升高的趋势,但总体呈降低趋势。结合趋势线及整体趋势图,群落土壤全磷含量在东西和南北方向上没有十分显著的变化趋势,整体变化较平稳,说明短丝木犀植物群落样方内的含量很均匀。
2.4 检验群落土壤中营养元素含量数据分布
通过图5可明显看出4种土壤养分含量分布数据接近正态分布,但不完全符合正态分布特点,且经SPSS对4种土壤养分含量数据分析得出的P值(K-Stest)分别为0.041(有机质)、0.036(全氮)、0.019(全磷)和0.022(全钾),小于0.05,不符合正态分布,与QQPlot图结果一致。
因此,通过对4种土壤养分含量数据进行转换后的正态分布检验,结果(图6)表明转换后的数据QQPlot分布图呈直线分布,符合正态分布特点。再利用SPSS软件进行检验,经K-S正态分布检验后得出P(K-Stest)值分别为0.613(有机质)、0.425(全氮)、0.164(全磷)和0.423(全钾),大于0.05,数据符合正态分布,说明经对数转换后的数据服从正态分布,则后续应选用对数正态克里格法进行空间插值,对4种土壤养分含量的分布做出预测。

蓝色线表示南北方向上群落土壤养分含量的变化趋势;绿色线表示东西方向上群落土壤养分含量的变化趋势特征;X轴表示群落正东方向,Y轴表示群落正北方向,Z轴表示点位测量值的大小。A. 有机质含量;B. 全氮含量;C. 全磷含量;D. 全钾含量图4 短丝木犀群落土壤4种养分含量趋势分析图The blue line indicates the change trend of the soil nutrient content of the community south-north; The green line indicates the change trend of the soil nutrient content of the community east-west; The X-axis represents the community due east; The Y-axis represents the community due north; The z-axis is the size of a point measurement. A. Organic matter content; B. Total nitrogen content; C. Total phosphorus content; D. Total potassium contentFig.4 Analysis on the trend of four kinds of nutrient in community soil
2.5 群落土壤营养含量的空间分布
基于对土壤元素含量的空间分布特征的分析,利用ArcGIS对东拉山短丝木犀土壤养分含量进行克里格空间插值,绘制土壤养分含量空间插值预测图(图7)。从图中可看出群落中部土壤有机质含量明显高于其他区域,呈较大范围的岛状分布,同时,分布地的北、南及西北部均有小范围岛状分布区域。这些有机质岛状分布区域是东拉山短丝木犀分布的中心位置,具有较为稳定的植被格局,由于形成较早,土壤扰动较少,环境稳定,自然环境代谢物质在土壤中大量积累,有机质丰富,因此有很多自然生长状况良好的大胸径成年大树,表现出较为稳定的群落格局。
另外,分布区的全磷和全钾西部明显高于其他区域,全磷和全钾含量最高值的土壤样点也在此范围,但在分布图中不影响该区域全磷和全钾含量整体水平。分布区南部有全磷和全钾含量较高的分布区域,但是岛屿面积较分布区的北部和东部较大范围区域全磷和全钾含量都较小。研究区沉积物中全氮含量自南向北逐渐增加,全氮含量的低值出现在东南部,高值出现在西北部,并形成了几个比较明显的岛状高值区域。
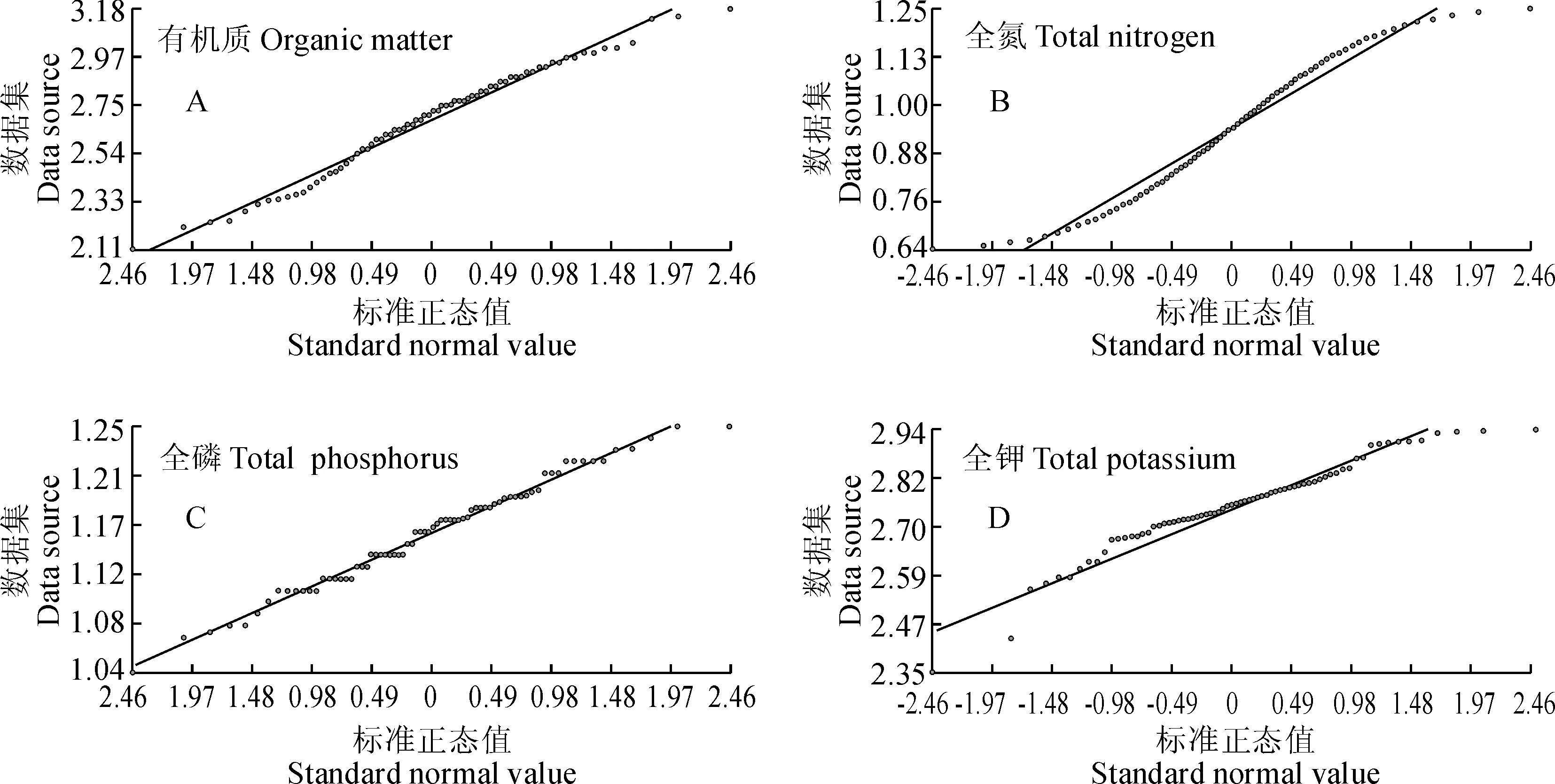
图5 短丝木犀群落土壤4种养分含量QQPlot图Fig.5 Four nutrients contents in soil community QQPlot map
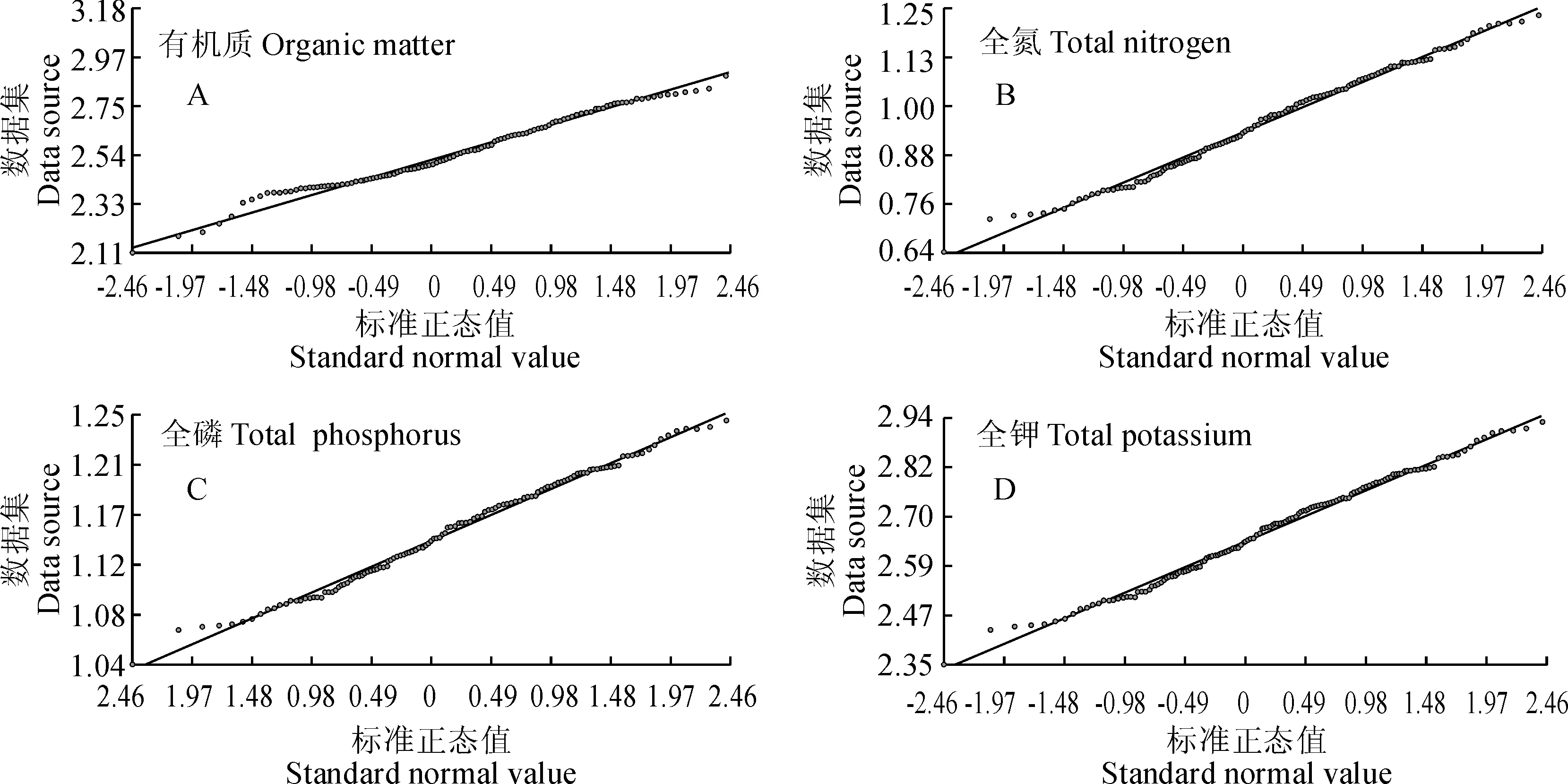
图6 对数转换后的短丝木犀群落土壤4种养分含量QQPlot图Fig.6 The four nutrients in the soil community QQPlot view after the digital conversion

图7 短丝木犀群落土壤4种养分含量的克里格插值预测图Fig.7 Nutrients of the four kinds of soil communities Kriging forecast map

图8 短丝木犀群落的克隆繁殖苗Fig.8 Clonal propagation of the community of O. serrulatus
3 讨 论
物种分布及其多样性与环境因子的关系受到研究尺度的强烈影响[20-21],从大的尺度上来说,气候和地形因子对植物群落的构建都是主导因素;从小的范围来说,土壤理化特征与植物组成则存在一定的耦合作用关系,一方面共同受到地形因素的制约,另一方面则相互产生影响。土壤是陆生植被栖息分布的基质,为植物提供必需的水分和营养物质,是植被分布的重要影响因子之一[22-23],而山地土壤中,有机质以及氮、磷、钾等土壤理化因子与植被群落组成和分布都有显著的关系[24-25]。
土壤的营养元素是土壤肥力的基本属性,也是植物群落的重要环境因子。研究表明,在同一山体成土母质一致的条件下,由于地上植物群落的差异,土壤营养元素的含量亦不相同。在自然条件下,植被生产力及植物群落的物种组成大都受到养分的限制,由于氮素和磷素在植物物质合成方面的重要作用,氮素和磷素在退化植被中的限制作用更为明显[26-27]。
郝日明等[14]研究表明,野生桂花群落地分布在石灰岩山地,群落内有裸露的岩石,土层薄。短丝木犀的生境地多为山体活跃、土层较浅的地区,土层的营养物质保证了成年株短丝木犀的生长。短丝木犀群落土壤中有机质含量高,土壤肥力高,这与林下有大量苔藓植物生存有关。这些植物一方面分泌多种酸性物质,加速成土母质的风化、分解,另一方面这些植物的残体、凋落物进入土壤,增加了土壤有机质的含量。
东拉山地区属于典型的石灰岩山地,由于石灰岩易受雨水冲刷,山体较为活跃,形成石灰岩碎石坡,继而在地表形成沟壑。所以群落内土壤的保水力、持肥力都较低,只有像短丝木犀这样耐瘠薄、干旱的物种才能够正常生长,完成生活史。
东拉山短丝木犀群落土壤中全钾含量在群落西南方位的陡坡含量相对较低,在西部地区由北向南呈现递减的趋势,主要因为土壤全钾含量受土母质影响较大,且向下淋溶易被土壤粘粒吸附,不具有表聚性,所以陡坡上含量低。群落的西北地区沟壑纵横,雨水冲刷严重,山体活动频繁,短丝木犀的枝条易受外力作用而被土壤和碎石积压。而土壤中钾含量是促进根部生长的主要元素,所以被土壤和地被植物掩盖的短丝木犀枝条逐渐形成不定根,慢慢形成新植株(图8),这种克隆繁殖方式也是很多野生桂花的繁衍更新方式[28]。
4 结 论
短丝木犀物种自身独特的生长习性与生境条件影响着种群的更新方式和种群结构及其遗传结构,进而影响种群的稳定性和物种的生存状态。相对于一般常绿阔叶林树种, 短丝木犀耐贫瘠的能力较强,可适应山体活跃、土层较浅的生境,形成优势纯林。在没有外界干扰的条件下,短丝木犀林下土壤在其地上植被的干扰下继续发育,土壤中有机质含量升高,土壤的保水力、持肥力不断增强,它已经能够适应包括常绿阔叶林树种的多种物种的生存。但是由于短丝木犀的种子在自然生境下存在隔年萌发的现象[29],幼苗的生长不耐荫性等因素,群落自身的发育受到限制。
东拉山短丝木犀植物群落的分布明显与群落土壤中有机物、氮、磷和钾的含量相关,群落土壤的有机质含量和山地气候的敏感关系,决定了短丝木犀的分布地范围,全N、P和K等营养元素则直接决定了短丝木犀群落的更新繁衍方式。短丝木犀植被群落通过改变林下凋落物的质与量和非生物环境等方式影响群落生态系统内土壤养分资源的循环和分布,土壤养分资源的量及空间分布也会反过来影响短丝木犀群落结构及分布。最终的结果是群落土壤作为整个短丝木犀群落生态系统的一部分在生态系统中达到物质能量的平衡, 使得短丝木犀群落在地理隔离间形成适用自身群落生长的小生态系统的“孤岛”。
研究短丝木犀群落的土壤有机质、全氮、全磷、全钾的空间分布规律不仅有助于探明短丝木犀独特生境的生态过程和功能,而且为合理利用和保护破碎化的短丝木犀群落提供科学依据。
参考文献:
[1]EL-GHANI M M A, AMER W M. Soil-vegetation relationships in a coastal desert plain of southern Sinai, Egypt[J].JournalofAridEnvironments, 2003,55(4): 607-628.
[2]GUISAN A, ZIMMERMANN N E. Predictive habitat distribution models in ecology[J].EcologicalModelling, 2000,135(2-3): 147-186.
[3]CRITCHLEY C N R, CHAMBERS B J, FOWBERT J A,etal. Plant species richness, functional type and soil properties of grasslands and allied vegetation in English Environmentally Sensitive Areas[J].Grass&ForageScience, 2002,57(2): 82-92.
[4]胡忠良, 潘根兴, 李恋卿, 等. 贵州喀斯特山区不同植被下土壤C、N、P含量和空间异质性[J]. 生态学报, 2009,29(8):4 187-4 195.
HU Z L, PAN G X, LI L Q,etal. Changes in pools and heterogeneity of soil organic carbon, nitrogen and phosphorus under different vegetation types in Karst mountainous area of central Guizhou Province, China[J].ActaEcologicaSinica, 2009,29(8): 4 187-4 195.
[6]张金屯. 数量生态学[M]. 北京:科学出版社, 2011.
[7]PORAZINSKA D L, BARDGETT R D,etal. Relationships at the aboveground-belowground interface: plants, soil biota, and soil processes[J].EcologicalMonographs,2003,73(3): 377-395
[8]汪敬忠, 吴敬禄, 曾海鳌, 等. 河套平原表层土壤元素含量与空间分布特征[J]. 沉积学报, 2014,32(4): 677-683.
WANG J Z, WU J L, ZENG H A,etal. Element content and spatial distribution of surface soil in Hetao area[J].ActaSedimentologicaSinica, 2014,32(4): 677-683.
[9]向其柏, 刘玉莲. 桂花资源的开发与应用现状及发展趋势[J]. 南京林业大学学报(自然科学版), 2004,28(s1): 104-108.
XIANG Q B, LIU Y L. The present and development strategy of produetion, development, popularization and utilization of sweet osmanthus[J].JournalofNanjingForestryUniversity(Natural Sciences Edition), 2004,28(s1): 104-108.
[10]刘龙昌, 向其柏. 樨属植物的研究进展[J]. 南京林业大学学报(自然科学版), 2003, (2): 84-88.
LIU L C, XIANG Q B. Research progress ofOsmanthusLour.[J].JournalofNanjingForestryUniversity(Natural Sciences Edition), 2003, (2): 84-88.
[11]臧德奎, 向其柏, 郝日明. 木犀属植物的分布与开发利用[J]. 西南林学院学报, 2004, (1): 23-26.
ZANG D K, XIANG Q B, HAO R M. Study on distribution and utilization of plants in genusOsmanthus[J].JournalofSouthwestForestryUniversity, 2004, (1): 23-26.
[12]吴晓娜, 何飞, 刘兴良, 等. 川西地区种子植物种级水平植物区系地理研究[J]. 四川林业科技, 2009,30(5): 35-43.
WU X N, HE F, LIU X L,etal. Research on flora of species of seed plants in western Sichuan, China[J].JournalofSichuanForestryScienceandTechnology, 2009,30(5): 35-43.
[13]陈俊华, 何飞, 李建彬, 等. 东拉野桂花群落物种多样性及乔木优势种生态位初步研究[J]. 四川林业科技, 2007,28(4): 48-51.
CHEN J H, HE F, LI J B,etal. Primary research on species diversity and niche characteristics of dominant arbor species inOsmanthusserrulatuscommunity[J].JournalofSichuanForestryScienceandTechnology, 2007,28(4): 48-51.
[14]MA K P. Measurement methods of community biodiversity I within-habitat diversity measure party[J].BiodiversityScience, 1994,3(sw): 231-239.
[15]WANG F. Responses to fertilization of rice on cold-waterlogged paddy fields developed from different landforms[J].Cold-waterloggedPaddyField, 2013,28(8):802-806.
[16]雷志栋, 杨诗秀, 许志荣, 等. 土壤特性空间变异性初步研究[J]. 水利学报, 1985, (9): 12-23.
LEI Z D, YANG S X, XÜ Z R,etal. A preliminary study on spatial variability of soil properties[J].JournalofHydraulicEngineering, 1985, (9): 12-23.
[17]李海东, 张波, 沈渭寿, 等. 苏南山丘区小流域土壤养分特性空间分布[J]. 长江流域资源与环境, 2009,18(9): 831-836.
LI H D, ZHANG B, SHEN W S,etal. Spatial variability of soil nutrients at a catchment of hilly area in southern Jiangsu Province[J].ResourEnvironYangtzeBasin, 2009,18(9): 831-836.
[18]王绍强, 朱松丽, 周成虎. 中国土壤土层厚度的空间变异性特征[J]. 地理研究, 2001,20(2): 161-169.
WANG S Q, ZHU S L, ZHOU C H. Characteristics of spatial variability of soil thickness in China[J].ActaGeographicaSinica, 2001,20(2): 161-169.
[19]汤国安. ArcGIS地理信息系统空间分析实验教程[M]. 北京:科学出版社, 2012.
[20]KUNIN W E. Extrapolating species abundance across spatial scales[J].Science, 1998,281(5 382): 1513.
[21]孟斌, 王劲峰, 张文忠, 等. 基于空间分析方法的中国区域差异研究[J]. 地理科学, 2005, (4): 11-18.
MENG B, WANG J F,etal. A review on the methodology of scaling with geodata[J].ActaGeographicaSinica, 2005, (4): 11-18.
[22]韩冰, 欧阳志云, 徐卫华, 等. 我国西南地区物种多样性保护效果[J]. 生态学报, 2008, (9):4 589-4 593.
HAN B, OUYANG Z Y, XÜ W H,etal. Soil carbon sequestration and its potential by cropland ecosystems in China[J].ActaEcologicaSinica, 2008, (9): 4 589-4 593.
[23]张广帅, 邓浩俊, 俞伟, 等. 泥石流频发区山地土壤环境因子与植被群落垂直梯度及其关系分析——以云南小江流域为例[J]. 应用与环境生物学报, 2014,20(4): 646-654.
ZHANG G S, DENG H J, YU W,etal. Vertical gradient analysis of soil environmental factors and vegetation community in mountain areas of frequent debris flow: a case study in the Xiaojiang Watershed, Yunnan[J].ChineseJournalofAppliedandEnvironmentalBiology, 2014,20(4): 646-654.
[24]冯云, 马克明, 等. 辽东栎林不同层植物沿海拔梯度分布的DCCA分析[J]. 植物生态学报, 2008,32(3): 568-573.
FENG Y, MA K M,etal. DCCA analysis of plant species distributions in different strata of oak (Quercusliaotungensis) forest along an altitudinal gradient in Dongling mountain, China[J].JournalofPlantEcology(Chinese Version), 2008,32(3): 568-573.
[25]于顺利, 马克平, 徐存宝, 等. 环境梯度下蒙古栎群落的物种多样性特征[J]. 生态学报, 2004,24(12): 2 932-2 939.
YU S L, MA K P, XÜ C B,etal. The species diversity characteristics comparison ofQuercusmongolicacommunity along environmental gradient factors[J].ActaEcologicaSinica, 2004,24(12): 2 932-2 939.
[26]CHEN J, STARK J M. Plant species effects and carbon and nitrogen cycling in a sagebrush-crested wheatgrass soil[J].SoilBiology&Biochemistry, 2000,32(1): 47-57.
[27]DIX J, LEITE J, SATOH K. Soil nitrogen transformation and cycling in forest ecosystem[J].ChineseJournalofEcology, 2004,23(5): 126-133.
[28]赵宏波, 郝日明, 胡绍庆. 中国野生桂花的地理分布和种群特征[J]. 园艺学报, 2015,42(9): 1 760-1 770.
ZHAO H B, HAO R M, HU S Q. Geographic distribution and population characteristics ofOsmanthusfragrans[J].ActaHorticulturaeSinica, 2015,42(9): 1 760-1 770.
[29]杨国栋, 王贤荣, 等. 中国特有香花植物短丝木犀种子萌发研究[J]. 农业科技与信息: 现代园林, 2015, (10): 761-765.
YANG G D, WANG X R,etal. A study on seed germination of an aromatic plant that endemic in China-Osmanthusserrulatus[J].ModernLandscapeArchitecture, 2015, (10): 761-765.
