中国和越南苔类植物新记录种
——柏氏细鳞苔
2018-04-08韦玉梅唐启明TruongVanDo朱瑞良
韦玉梅,唐启明,Truong Van Do,朱瑞良
(1 广西壮族自治区中国科学院广西植物研究所 广西喀斯特植物保育与恢复生态学重点实验室,广西桂林 541006;2 华东师范大学生命科学学院,上海 200062;3 越南国家自然博物馆 维管束植物分类与多样性保育中心,越南河内 100803)
细鳞苔科(Lejeuneaceae)是苔类植物门(Marchantiophyta)中最大的科,是苔类植物中种类较多、分类十分困难的一个类群[1]。细鳞苔属(Lejeunea)隶属于细鳞苔科,是细鳞苔科的模式属。目前全世界报道的细鳞苔属植物有近400种[2],是细鳞苔科中仅次于疣鳞苔属(Cololejeunea)的第二大属。该属植物主要分布在热带及亚热带地区,温带地区也有少量分布。中国是细鳞苔属植物分布较为丰富的地区之一,目前已报道有43种[3-4],越南分布22种[5]。笔者在对中国和越南的细鳞苔属植物进行研究的过程中,发现了一种在形态上与中国和越南现有报道的细鳞苔属植物有明显区别的植物,经文献资料和模式标本的核查,确认其为中国和越南苔类植物新记录种——柏氏细鳞苔(新拟)[Lejeuneapatersonii(Steph.) Steph.]。现整理报道,以期为未完成的中国苔藓志增加新资料。
1 柏氏细鳞苔 (图1)
Lejeuneapatersonii(Steph.)Steph., Sp. Hepat. 5: 784. 1915. (Fig.1)
EulejeuneapatersoniiSteph., Hedwigia 35: 92. 1896. Type: Indonesia. Java, s.d.,Patersons.n. (holotype: G 12343!).
=LejeuneacrenulataMitt. In Seemann, Fl. Vit.: 415. 1873“1871”. Type: Samoa.Powells.n.
=MicrolejeuneasamoanaSteph., Hedwigia 35: 115. 1896. =Lejeuneasamoana(Steph.) R.M.Schust., J. Hattori Bot. Lab. 26: 249. 1963. Type: Fiji. Ovalau, 1864,Graeffes.n.
=PrionolejeuneacaledonicaSteph., Sp. Hepat. 5: 227. 1913. Type: New Caledonia. Tao, 600 m, Jan. 1910,Franc245.
植物体绿色,干时淡黄色。假根少,无色、透明,簇生于腹叶基部,假根盘缺。茎横切面近圆形,包括7个表皮细胞和6~11个内部细胞,无中轴分化,直径0.75~1.00 mm。植物体长约8~13 mm,带叶宽0.60~0.90 mm;不规则分枝,分枝细鳞苔型。叶覆瓦状排列或毗邻,背瓣卵形,长0.40~0.60 mm,宽0.30~0.45 mm,表面平展,顶端圆形或钝形,稍内弯,边缘具圆齿。叶边缘细胞方形,13~23 × 10~20 μm,中部细胞六边形,25~48 × 20~33 μm,基部细胞相似于中部细胞,35~50 × 25~33 μm,细胞壁薄,角质层平滑,三角体不明显或中等大,中部球状加厚缺。油体未见。油胞和假肋缺。腹瓣小,卵形,长约为背瓣的1/4~2/5,鼓起,近轴边缘略内卷,顶端斜截形,中齿单细胞,角齿不明显,透明疣位于中齿基部的近轴侧,脊拱起。腹叶远生,近圆形,长0.10~0.13 mm,宽0.10~0.17 mm,宽略大于长,宽为茎的1~2倍,顶端2裂达1/2深,裂瓣边缘具明显或不明显的圆齿,基部与茎连接部分呈浅弧形。雄穗未见。雌器苞常生于侧短枝上,具1个新生枝,新生枝叶发生顺序细鳞苔型。雌苞叶背瓣长椭圆形,长约0.45 mm,宽约0.25 mm,顶端圆形或钝圆形,边缘具明显或不明显的圆齿;雌苞叶腹瓣长椭圆形,长为雌苞叶背瓣的2/3,顶端钝形或截形,脊长为雌苞叶腹瓣的1/2~1/3;雌苞腹叶长椭圆形,长约0.40 mm,宽约0.25 mm,顶端2裂达1/2~1/3深,裂瓣边缘具圆齿。蒴萼未见。无性繁殖器官未见。
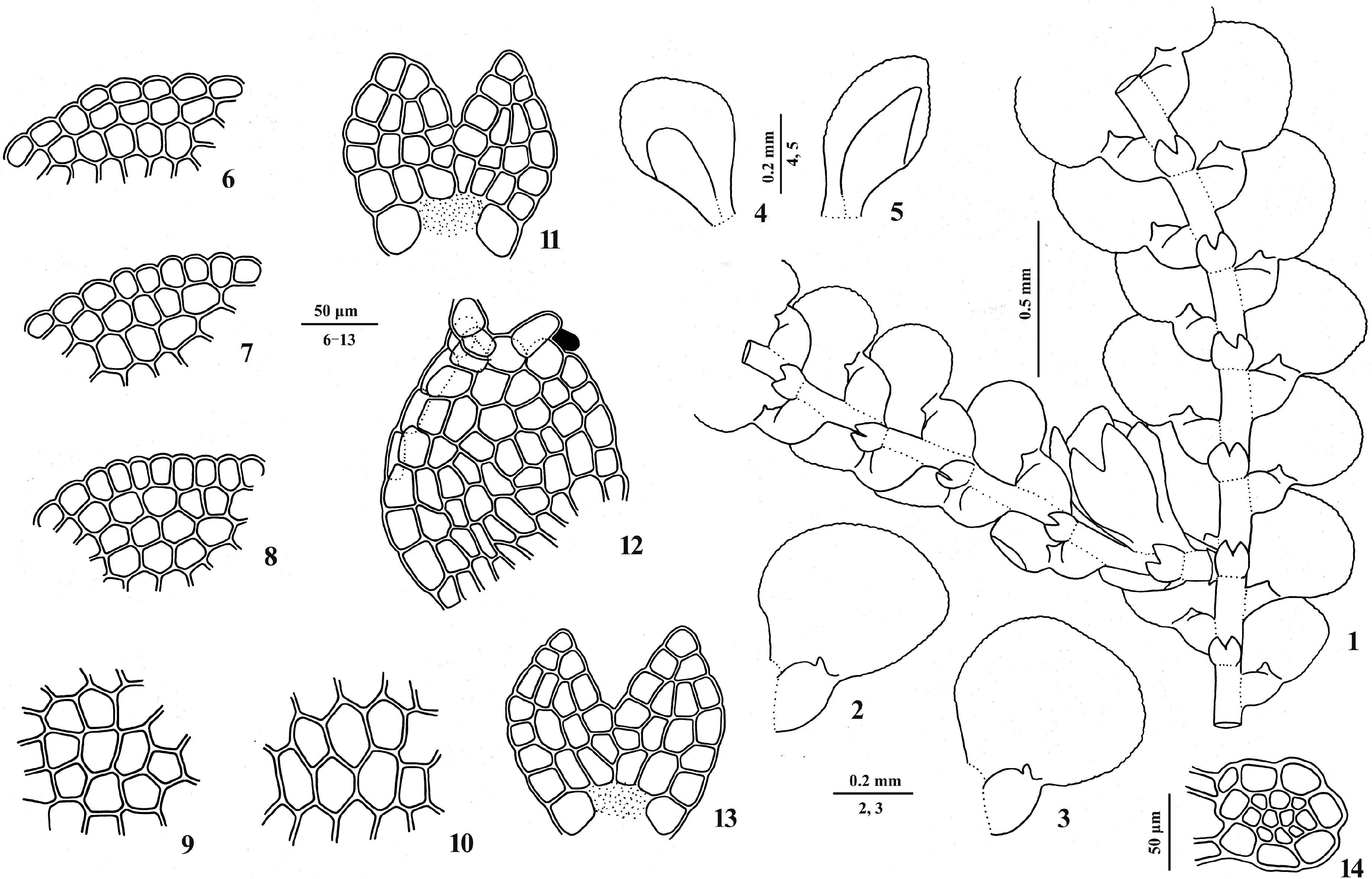
1. 植物体一段带雌枝; 2、3. 叶; 4、5. 雌苞叶; 6~8. 叶边缘细胞; 9. 叶中部细胞; 10. 叶基部细胞; 11、13. 腹叶; 12. 腹瓣; 14. 茎横切面图1 柏氏细鳞苔(绘图标本: 韦玉梅等20141205-23)1. Part of plant with gynoecium; 2, 3. Leaves; 4, 5. Female bracts; 6-8. Margin cells of leaf lobe; 9. Median cells of leaf lobe; 10. Basal cells of leaf lobe; 11, 13. Underleaves; 12. Leaf lobule; 14. Transverse section of stemFig.1 Lejeunea patersonii (Steph.) Steph. (drawn from Wei et al. 20141205-23)
标本鉴定:中国:广西那坡县百都乡达省屯沟谷,23°12′04.15″ N, 105°32′46.38″ E,海拔676 m,韦玉梅等20141205-11 (IBK);地点同前,23°12′04.54″ N, 105°32′44.71″ E,海拔657 m,韦玉梅等20141205-23 (IBK);越南:河江省(HGiang Province,District, Tùng Vi Commune, Thăng Village), 23°03′01.26″ N, 104°52′04.12″ E, 海拔1 099 m, 朱瑞良,韦玉梅等20131103-113 (HSNU)。
生境:树基或岩面薄土生。
分布:印度尼西亚(爪哇),萨摩亚,斐济,新喀里多尼亚,菲律宾[6],马来西亚[7],巴布亚新几内亚[8],密克罗尼西亚(加罗林群岛)[9],塔希提岛[10],泰国[11]。中国和越南为首次记录。
2 讨 论
细鳞苔属植物形态被认为是最具多样化的一个类群[12],因此,其属的界定一直存在争议。近年来,随着分子系统学方法的普及和近期细鳞苔科主要类群的深入研究,一些与细鳞苔属关系长期存在争论的属被归并为细鳞苔属的异名,同时细鳞苔属与其他近缘属之间的关系也得到了澄清[13]。目前,细鳞苔属已能从形态上通过下列特征获得很好的界定[14-16]:植物体分枝细鳞苔型;茎横切面常具7个皮部细胞和3个以上内部细胞;叶不具油胞;腹叶全缘到深二裂;腹瓣顶部具1~2个齿,角齿常退化,透明疣位于中齿基部的近轴侧;油体小,聚合型或均一型;雌器苞新生枝叶发生顺序为细鳞苔型;细胞角质层常具细疣。
2.1 柏氏细鳞苔与相近种的形态比较
中国和越南新分布的柏氏细鳞苔是典型的细鳞苔属植物,其区别于其他细鳞苔属种类的主要特征是:侧叶顶端圆形或钝形,叶边缘具圆齿;腹叶小,远生,宽为茎的1~2倍;叶细胞角质层平滑,三角体不明显或中等大,中部球状加厚缺。其中,叶边缘具有明显的圆齿是柏氏细鳞苔的一个显著特征,而这样的特征在细鳞苔属植物中并不多见。目前,亚洲及大洋洲分布的细鳞苔属植物中已知的仅5种:Lejeuneaalbescens(Steph.) Mizut.、Lejeuneaasperula(Steph.) Mizut.、LejeuneakinabalensisMizut.、LejeuneamicrolobaTaylor和Lejeuneaumbilicata(Nees) Nees[17-19]。其中,L.albescens和L.kinabalensis具有大型的腹叶(宽可达茎的4倍以上),且腹叶基部呈耳状等特征区别于柏氏细鳞苔;L.asperula和L.microloba侧叶顶端具锐尖或细尖明显不同于柏氏细鳞苔;而典型的L.umbilicata通过腹叶和蒴萼的特征很容易与柏氏细鳞苔进行区分,前者腹叶是茎宽的3~4倍,蒴萼平滑无脊,后者腹叶是茎宽的1~2倍,蒴萼具有5个明显的脊[18]。但值得注意的是,L.umbilicata的蒴萼并不是一个稳定的特征,具脊的情况也是存在的[19],而且在植物体发育不典型的情况下,腹叶有可能会变得很小,极易与柏氏细鳞苔混淆,这种情况下,两者仍可以通过以下特征很好地区分开来:L.umbilicata的叶细胞角质层具细疣,三角体明显,中等大,具中部球状加厚,而柏氏细鳞苔叶细胞角质层平滑,三角体不明显或中等大,中部球状加厚缺。柏氏细鳞苔是目前中国报道的细鳞苔属植物中唯一一种侧叶边缘具有明显圆齿特征的种类,可明显区别于中国其他细鳞苔属植物。
2.2 柏氏细鳞苔的地理分布
柏氏细鳞苔最早报道于印度尼西亚的爪哇岛(Java)上,随后在大洋洲各大岛屿及亚洲其他热带地区陆续出现报道。1983年,Miller等曾对该种的分布进行过详细统计[20],而随后的相关研究中对柏氏细鳞苔的分布记录都是基于他们报道的数据,如最近其新报道于泰国一文[11]中所列出的分布记载。事实上,在Miller等对柏氏细鳞苔的分布记载中,由于他们错误地将Taxilejeuneapatersonii(Steph.) Eifrig (现为里拉细鳞苔Lejeunealeratii(Steph.) Mizut.)等同于柏氏细鳞苔,从而混淆了柏氏细鳞苔与里拉细鳞苔的分布数据[10]。根据笔者对相关文献资料的核实,发现在印度尼西亚的苏门答腊(Sumatra)、婆罗洲(Borneo)、苏拉威西(Celebes)地区[17]以及瓦努阿图(Vanuatu=New Hebrides)[21]的分布记载均来源于对里拉细鳞苔的记录,目前这些地区暂无柏氏细鳞苔的分布报道,因此,笔者建议将上述地区从柏氏细鳞苔的分布区中去除。基于此,本研究重新确认了柏氏细鳞苔的地理分布(图2)。根据柏氏细鳞苔的地理分布,可见该种广泛分布于热带亚洲及大洋洲地区,属于热带亚洲分布类型[22],是一个明显的热带地区种类。而广西中越边境地处中国热带北缘,是北热带和南亚热带交汇处,越南北部与该地区毗邻,柏氏细鳞苔分布区延伸至这些地方也实属情理之中。
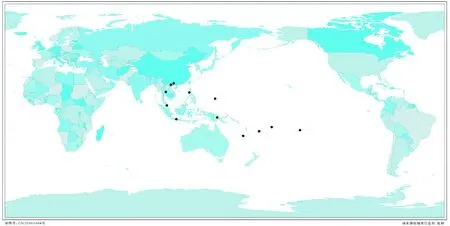
图2 柏氏细鳞苔地理分布图Fig.2 Distribution of Lejeunea patersonii (Steph.) Steph
致谢:感谢海南大学张莉娜老师和河北师范大学李敏老师在论文的修改过程中提出的宝贵意见。
参考文献:
[1]WANG J, ZHU R L, GRADSTEIN S R. Taxonomic revision of Lejeuneaceae, subfamily Ptychanthoideae (Marchantiophyta) in China [M]. Stuttgart: J. Cramer in der Gebr. Borntraeger Verlagsbuchhandlung, 2016: 1-141.
[2]SÖDERSTRÖM L, HAGBORG A,etal. World checklist of hornworts and liverworts [J].PhytoKeys, 2016, 59: 1-828.
[3]高谦, 吴玉环. 中国苔纲和角苔纲植物属志 [M]. 北京: 科学出版社, 2010: 437-463.
[4]贾渝, 何思. 中国生物物种名录(第一卷 苔藓植物) [M]. 北京: 科学出版社, 2013: 398-407.
[5]SHU L, XIANG Y L,etal. New liverwort and hornwort records for Vietnam [J].Cryptogamie,Bryologie, 2017,38(4): 411-445.
[6]HERZOG T. Hepaticae Philippinenses a cl. C. J. Baker lectae [J].AnnalesBryologici, 1931, 4: 79-94.
[7]INOUE H. Studies on oil bodies of some Malayan liverworts [J].JournaloftheHattoriBotanicalLaboratory, 1967, 30: 54-70.
[8]GROLLE R. Einige ostmalesische Lebermoose [J].NovaHedwigia, 1968, 16: 147-159.
[9]SWANSON E S, MILLER H A. Hepaticae from Palau, Caroline Islands, II. Enumeration [J].Micronesica, 1969,5(1): 139-149.
[10]HÜRLIMANN H. Hepaticae aus dem Gebiete des südlichen Pazifik. XII [J].Bauhinia, 1993,11(1):3-17.
[12]WIGGINTON M J E W. Jones’s liverwort and hornwort flora of West Africa [J].ScriptaBotanicaBelgica, 2004, 30: 1-443.
[13]HEINRICHS J, DONG S,etal. Molecular phylogeny of the leafy liverwortLejeunea(Porellales): evidence for a Neotropical origin, uneven distribution of sexual systems and insufficient taxonomy [J].PLoSOne, 2013,8(12):e82547.
[14]ZHU R L, SO M L. Epiphyllous liverworts of China[M]. Stuttgart: J. Cramer in der Gebr. Borntraeger Verlagsbuchhandlung, 2001, 126-162.
[15]GRADSTEIN S R. A classification of Lejeuneaceae based on molecular and morphological evidence [J].Phytotaxa, 2013,100(1): 6-20.
[17]EIFRIG H. Monografische Studienüber die Indomalayischen Arten vonTaxilejeunea[J].AnnalesBryologici, 1937, 9: 73-114.
[18]MIZUTANI M. Lejeuneaceae, subfamilies Lejeuneoideae and Cololejeuneoideae, from Sabah (North Borneo) [J].JournaloftheHattoriBotanicalLaboratory, 1970, 33: 225-265.
[19]LEE G E. A systematic revision of the genusLejeuneaLib. (Marchantiophyta: Lejeuneaceae) in Malaysia [J].Cryptogamie,Bryologie, 2013,34(4): 381-484.
[20]MILLER H A, WHITTIER H O, WHITTIER B A. Prodromus Florae Hepaticarum Polynesiae. Catalogue of Hepaticae and Anthocerotae[M]. Stuttgart: J. Cramer in der Gebr. Borntraeger Verlagsbuchhandlung, 1983: 1-423.
[21]TIXIER P. Bryophytes exotiques [J].Botanique, 1973, 10: 73-86.
[22]吴征镒, 孙航, 周浙昆, 等. 中国种子植物区系地理 [M]. 北京: 科学出版社, 2010: 109-319.
