贵妃鸡多趾性状遗传规律分析及多趾候选基因的表达
2018-04-04王香南郭红威郭亚苹闫峰宾王彦彬孙桂荣康相涛韩瑞丽
王香南,郭红威,郭亚苹,闫峰宾,王彦彬,孙桂荣,康相涛,韩瑞丽
(河南农业大学牧医工程学院,郑州 450002)
多趾(指)是在人、鼠、鸡等物种中常见的肢体发育畸形,鸡与人、鼠有着相似的表型。在人上,该畸形可单独发生(非综合征型),又可以合并多种其他疾病同时发生(综合征型)[1]。根据受累指(趾)的位置,L.G.Biesecker等又将非综合征型多指(趾)分为轴前多指(Preaxial polydactyly,PPD)和轴后多指(Posterior polydactyly,PAP)两大类[2-3],且每种类型的畸形都具有丰富的解剖形态[4]。鸡通常表现为四趾,鸡的四趾是高等动物第五趾丢失的结果,但鸡的多趾不是丢失第五趾的恢复,而是重新在另一端长出了新趾。鸡多趾性状有不同的表现形式,既有趾骨的增加也有趾数目的增加[5]。比较著名的多趾品种有北京油鸡、丝羽乌骨鸡、贵妃鸡等,而对贵妃鸡多趾性状的研究却相对较少。贵妃鸡是法国著名品种,其多趾性状与国内品种的多趾性状有着不同的进化模式,其遗传规律和主要控制基因也可能存在差异[6]。
多指(趾)表型的变异是由于遗传和环境共同作用的结果,遗传是主要的原因。以往的研究表明,非综合征型多指(趾)的遗传方式多为常染色体显性遗传,也有常染色体隐性遗传[7]。早期的杂交试验结果显示,鸡的多趾性状虽为常染色体显性遗传,但多趾性状的外显率却不同[8-11]。另外,J. B. Hutchinson等[12-16]认为,双亲都为多趾的个体后代出现四趾个体是由于遗传修饰,而表现出不同的外显率,这和Lmbr1基因与多趾性状不完全连锁亦有关。邓学梅[17]在对丝羽乌骨鸡和白洛克肉鸡杂交系的研究中,将趾数基因定位在2号染色体的45 cM处。另外,K.Yang等[18]通过对北京油鸡多趾突变与染色体2p区域的关联分析,将多趾位点定位于染色体2p区域的ABR0004~MCE0184之间。Y.F.Sun等[19]发现,Lmbr1基因是多趾性状的重要候选基因。Y.Q.Huang等[20]对Lmbr1基因进行了SNP分析,发现G1255A位点与多趾性状极显著相关。曲鲁江等[21]公开了鸡多趾性状相关的SNP(Lmbr1 C4645A)分子标记及其应用的专利,可以通过SNP的方法准确的分别出丝羽乌骨鸡和北京油鸡的多趾与非多趾的基因型。近期C.He等[22]通过北京油鸡与石歧杂鸡的杂交试验,并对F2代群体的多趾性状进行了全基因组关联分析,重测序中发现,rs80659072的G/T突变与多趾性状高度相关,且证明了多趾性状属于完全显性遗传,而不是以前猜想的不完全外显或遗传修饰。
本试验以不同趾型的贵妃鸡为研究对象,通过杂交、测交方式和多趾候选基因的表达分析,旨在进一步探索贵妃鸡多趾性状的遗传规律和主要候选基因,是对鸡多趾性状研究的重要补充,也能为人类多趾(指)研究提供参考。
1 材料与方法
试验于2016年5月-2017年3月在河南农业大学种质资源场和河南省家禽种质资源创新工程研究中心完成。
1.1 试验材料
1.1.1试验动物试验所用贵妃鸡由河南农业大学家禽种质资源场提供。根据贵妃鸡左脚和右脚的脚趾数目的不同,将贵妃鸡的趾型分为四四趾、四五趾和五五趾3种表型。
1.1.2胚胎组织试验用到的胚胎组织采自胚胎发育的第5~10天,其双亲均为五五趾。
1.2 试验方法
1.2.1趾型的检测通过肉眼观察的方法对所有试验动物进行趾型分型。如图1所示,左图是四趾,右图为五趾。

左图为四趾,右图为五趾The left is four toes and the right is five toes图1 贵妃鸡的趾型Fig.1 The toe types of Royal chicken
1.2.2杂交组合选择贵妃鸡四五趾公鸡6只、母鸡30只,贵妃鸡五五趾公鸡10只、母鸡60只。假设五趾性状对四趾为显性,五五趾个体的基因型为AA,四五趾个体的基因型为Aa,四四趾个体的基因型为aa。3种杂交组合方式分别为四五趾♂×四五趾♀、四五趾♂×五五趾♀和五五趾♂×五五趾♀。3个组合的饲养管理条件相同,同期进行人工授精,连续收集10 d种蛋。每个组合的种蛋分开放置、做好记录、严格标记,在同一时期进行孵化。
1.2.3测交组合选择贵妃鸡表型为五五趾的公鸡9只、表型为四四趾的母鸡36只。分别进行编号,1只公鸡配4只母鸡,饲养管理条件、人工授精方
式、种蛋的收集等与上述组合相同。做好孵化期间的数据记录,用尼龙网兜将测交组不同父本的种蛋相互分开,防止出壳无法辨认。
1.2.4趾型判定出雏当天通过肉眼观察确定每只个体的趾型,分为四四趾型、四五趾型、五五趾型。
1.2.5胚胎样品的采集选择五五趾♂×五五趾♀的种蛋,剔去不合格的种蛋,当天16:00点入孵。在孵化第5天的相同时间采集肢芽部组织样品,以后每隔24 h取样一次,每天采集5个样品。
1.2.6候选基因扩增引物的设计根据文献报道[7]选择影响多趾性状的6个主要候选基因Lmbr1、SHH、RNF32、NOM1、MNX1、Gli3,根据GenBank中提供的上述各基因的序列,利用Primer5.0软件进行引物设计,由上海生物工程科技服务公司合成。内参基因β-actin引物由河南农业大学家禽中心实验室提供,引物信息见表1。
表1荧光定量PCR引物序列
Table1Amplifyingprimersofcandidategenes
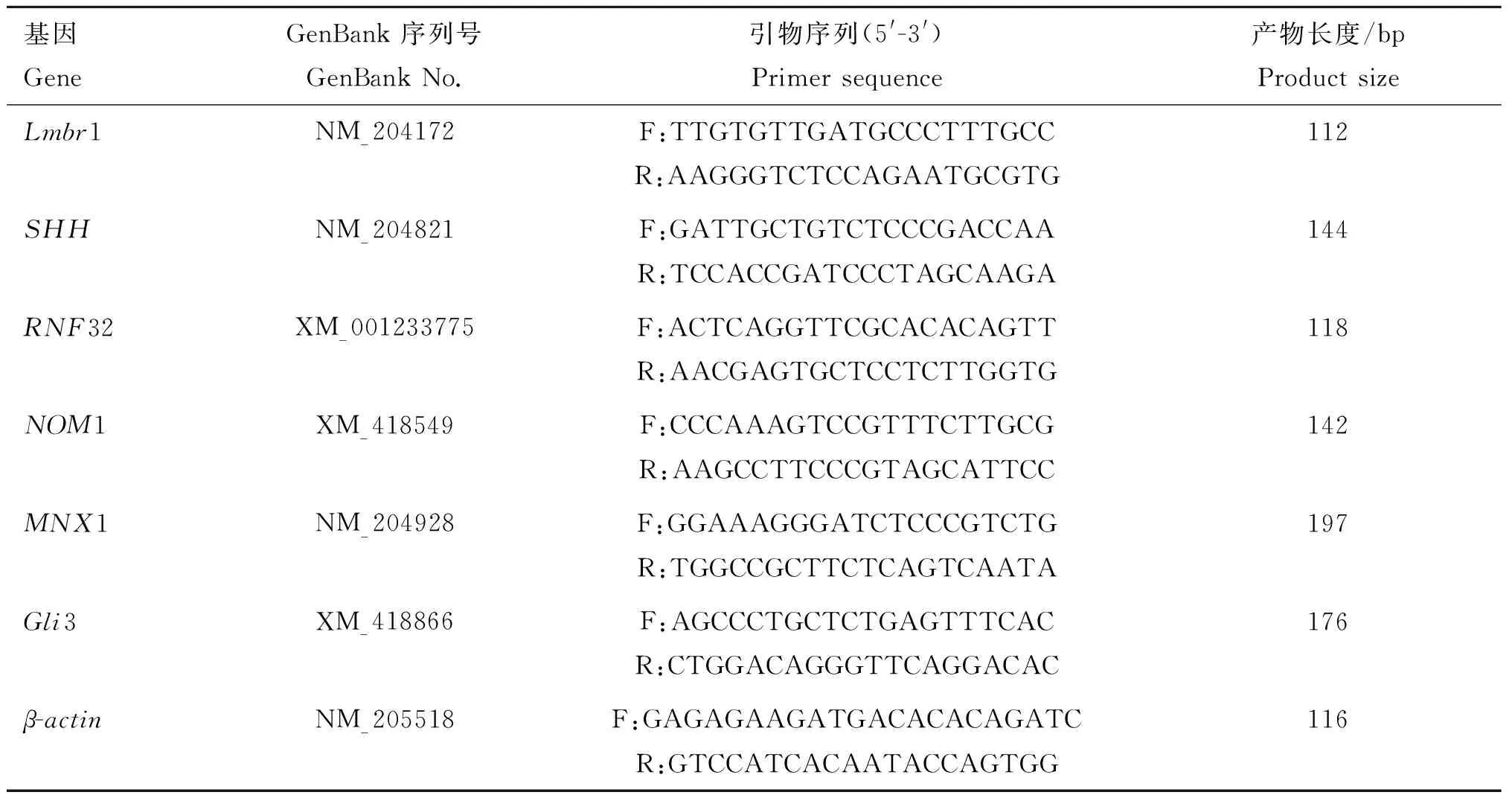
基因GeneGenBank序列号GenBankNo.引物序列(5'-3')Primersequence产物长度/bpProductsizeLmbr1NM_204172F:TTGTGTTGATGCCCTTTGCCR:AAGGGTCTCCAGAATGCGTG112SHHNM_204821F:GATTGCTGTCTCCCGACCAAR:TCCACCGATCCCTAGCAAGA144RNF32XM_001233775F:ACTCAGGTTCGCACACAGTTR:AACGAGTGCTCCTCTTGGTG118NOM1XM_418549F:CCCAAAGTCCGTTTCTTGCGR:AAGCCTTCCCGTAGCATTCC142MNX1NM_204928F:GGAAAGGGATCTCCCGTCTGR:TGGCCGCTTCTCAGTCAATA197Gli3XM_418866F:AGCCCTGCTCTGAGTTTCACR:CTGGACAGGGTTCAGGACAC176β-actinNM_205518F:GAGAGAAGATGACACACAGATCR:GTCCATCACAATACCAGTGG116
1.2.7荧光定量参考TaKaRa公司的RNAiso Plus试剂说明书提取组织总RNA,用NanoDrop2000紫外分光光度计检测组织总RNA的浓度和纯度。利用反转录试剂盒将RNA反转成cDNA,主要分为去除基因组DNA反应(5× gDNA Eraser Buffer 2 μL,gDNA Eraser 1 μL,总RNA 1 μg,加无RNase ddH2O至10 μL)和反转录反应(5×PrimeScript Buffer 2 μL,PrimeScript RT Enzyme Mix I 0.5 μL,RT Primer Mix 2 μL,上一步的反应液10 μL,加无RNase ddH2O至20 μL)。
以cDNA为模板,进行PCR扩增,反应体系:2×TaqMaster Mix 5.0 μL,上下游引物各0.5 μL,cDNA样品为1.0 μL,加ddH2O至总体积为10 μL。PCR程序:95 ℃预变性5 min;94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,35个循环。
1.2.8数据的统计与分析运用SPSS20.0软件进行数据处理,对实际趾型比例与理论比例进行卡方检验,P>0.05表示差异不显著,即实际趾型比例符合理论比例,P<0.05为差异显著,即实际趾型比例不符合理论比例。定量试验数据采用2-ΔΔCt法计算各目的基因的表达量,ΔCt=目的基因平均Ct值-内参基因平均Ct值。采用Duncan法进行单因素方差分析。运用Graphpad Prism5.0软件进行作图,数据最终以“平均值±标准差”显示。
2 结 果
2.1 不同趾型贵妃鸡杂交结果分析
3个杂交组合后代个体的趾型分布和实际比例、理论比例及卡方检验结果如表2所示。四五趾×四五趾组合后代个体多趾与正常趾的实际比例为3.67∶1,符合3∶1的理论比值(P>0.05),另外两个组合都不符合理论比值(P<0.05)。
2.2 五五趾型贵妃鸡测交结果分析
9只五五趾型贵妃公鸡与纯合四四趾型贵妃母鸡测交后代(F1代)的数量、趾型分布以及多趾与正常趾的比例见表3。
从表3中可以看出,不同父本后代的趾型分布以及多趾与正常趾的比例存在一定的差异。每只测交公鸡后代都出现五五趾、四五趾和四四趾3种表型,1号和2号公鸡后代出现的四四趾个体最多,都为9只,多趾与正常趾的比例分别为0.89∶1和1.22∶1;9号公鸡后代出现的四四趾个体最少,为1只,多趾与正常趾的比例为16.00∶1。计算得出,每只公鸡多趾性状对正常四趾的外显率分别为47.06%、55.00%、55.56%、65.22%、70.58%、73.91%、77.78%、87.50%、94.11%。
表2不同杂交组合趾型实际比例与理论比例
Table2Thepercentageofdifferenttoetypesforeachgroup

组合Group后代数量Amount3种趾型比Threetoetypesratio多趾:四趾Polydactyly:Four-fourtoe理论比值TheoreticalratioP值P-value四五趾×四五趾Four-fivetoe×Four-fivetoe14074∶36∶303.67∶13∶10.329四五趾×五五趾Four-fivetoe×Five-fivetoe14892∶23∶333.48∶11∶10.000五五趾×五五趾Five-fivetoe×Five-fivetoe329248∶50∶319.16∶11∶00.000
3种趾型依次是五五趾、四五趾、四四趾。P>0.05表示差异不显著,P<0.05表示差异显著
The three toe types are five-five toe, four-five toe, four-four toe, respectively.P>0.05 indicates no significant difference,P<0.05 indicates significant difference
表3不同测交组合后代的趾型分布
Table3Thetoetypesdistributionindifferentcrossbreedinggroups

公鸡编号Number(♂)F1代数量/只AmountofF1五五趾/只Five-fivetoe四五趾/只Four-fivetoe四四趾/只Four-fourtoe多趾∶四趾Polydactyly∶Four-fourtoe1177190.89∶12207491.22∶13183781.25∶142313281.88∶15176652.40∶16239862.83∶172716563.50∶18169527.00∶1917142116.00∶1
2.3 贵妃鸡肢芽组织胚胎发育等5~10天Lmbr1、SHH、RNF32、NOM1、MNX1、Gli3基因的表达情况
Lmbr1、SHH、RNF32、NOM1、MNX1、Gli3等基因在贵妃鸡胚胎发育的等5~10天肢芽组织的相对表达量的趋势如图2所示。可以看出,SHH基因在胚胎发育的等6~10天的表达量一直很低,其中第6天的表达量明显降低。Lmbr1、RNF32、NOM1、MNX1、Gli3基因在胚胎发育的第7和第9天出现表达峰值,其中第7天表达量明显升高,第9天Lmbr1、RNF32、Gli3基因的表达量也明显升高。候选基因在胚胎发育时期的表达量,第8天低于第7和第9天。

图2 Lmbr1、SHH、RNF32、NOM1、MNX1、Gli3在贵妃鸡胚胎发育时期相对表达量变化Fig.2 The relative expression trends of Lmbr1, SHH, RNF32, NOM1, MNX1, Gli3 in the limb bud tissue during embryonic development
3 讨 论
3.1 贵妃鸡多趾性状的遗传
本试验结果显示,3个杂交组合的多趾与四四趾的比率和理论值都存在差异,其中四五趾×四五趾组合的后代趾型比例与理论值接近,而四五趾×五五趾和五五趾×五五趾组合的后代趾型比例均与理论值不符。另外,五五趾×五五趾组合多趾性状的外显率为90.58%,此结果与汤琳琳[23]研究中五趾北京油鸡纯繁后代五趾性状比例为89.19%的结果基本一致。另有研究报道,纯合多趾亲本与正常四趾个体杂交后代个体多趾的外显率达96%左右,杂合多趾公鸡与正常四趾母鸡杂交后代个体多趾的外显率仅为79%,这表明杂合子多趾性状表达失败的概率要大于纯合子[12]。本试验中,每个杂交组合后代均出现了四四趾个体,在 D.C.Warren[11]的研究中,多趾杂合与多趾杂合的正反交试验中亦出现了少量的非多趾个体。 J.B.Hutchinson[12]认为,多趾双亲后代出现正常四趾的原因是由于基因修饰造成的,另有P.D.Sturkie[24]的研究表明,低温会抑制多趾基因的表达。本试验结果可以证实前人对于鸡多趾性状的研究,即鸡多趾性状和正常四趾一样呈常染色体显性遗传,但多趾性状却有着不同的外显率。从而推测,贵妃鸡的多趾性状也是属于显性遗传且外显率不同,该多趾性状可能不是由一对基因控制。
在测交试验中,尽管用于测交的公鸡表型都是五五趾,但后代却出现了不同趾型的个体。1~7号公鸡测交后代出现了较多比例的四四趾个体,推测1~7号公鸡可能为多趾杂合子,此种表型个体的后代还会出现四趾的个体,不符合实际生产对多趾个体的选育要求,在生产中应该及时淘汰。8~9号多趾公鸡的测交后代仅存在1~2个个体为四四趾,可能是试验操作中的各种误差或者人为错误造成的,而且8、9号公鸡测交后代五趾性状的外显率分别达到87.50%、94.11%,可用于近一步扩繁五五趾个体,但仍需进行测交分析。总之,通过测交试验,亦没能获得完全为五五趾纯合型的公鸡,可能是由于贵妃鸡的多趾性状并不是由一对等位基因决定的。
3.2 多趾候选基因在贵妃鸡胚胎发育时期的表达
已有的多趾性状的分子标记(Lmbr1 C4645A)不能够应用于贵妃鸡多趾性状的选择,根据前人对丝羽乌骨鸡多趾性状的研究,本试验对贵妃鸡多趾性状的候选基因做了进一步的研究,结果可知,Lmbr1、RNF32、NOM1、MNX1、Gli3分别在第7和第9天出现表达峰值,推测贵妃鸡的多趾性状可能受到Lmbr1、RNF32等基因的共同调控,多趾形成的关键时期可能是在胚胎发育的第7~9天。
杜晓惠[25]研究了Lmbr1基因在白来航与丝羽乌骨鸡胚胎第3~6天的表达,发现丝羽乌骨鸡Lmbr1基因的表达量均略高于白来航鸡,但都未达到显著水平,仅在第3~5天差异较大。在本试验结果中,Lmbr1基因在贵妃鸡胚胎发育的肢芽组织中第7和第9天有较高的表达量,这个可能是由于品种的特异性造成的。SHH基因的调控机制主要有两个:第一,Lmbr1基因第五内含子极化活性区调控序列(Zone of polarizing activity regulatory sequence,ZRS),可以作为增强子远程调控距其1 Mb左右的SHH基因在胚胎肢芽后缘表达,研究表明,SHH基因调控序列ZRS的406AG和105CG杂合突变分别为两个中国汉族三指节拇指轴前多指家系的致病突变。M.Davey等[16,26-28]针对荷兰人群做了ZRS增强子对SHH远程调控的新见解。第二,Gli3转录因子在调控SHH基因表达方面起着重要的作用。在脊椎动物中Gli家族包括3个成员,其中Gli3对肢发育的影响最大,其是一个编码锌指蛋白的基因。它的突变影响了自身正常表达,导致足轴前多趾、手轴后多趾、手足并指(趾)等,SHH功能的丧失导致严重的肢断裂,SHH在前肢的异常表达引起指(趾)的重复[7]。汤琳琳[23]检测了SHH在胚胎发育第6~10天的表达,发现除第6天外,五趾北京油鸡的表达量均高于石岐杂鸡。此结果表明,SHH在肢芽的表达导致多趾性状的形成,在正常四趾中检测不到SHH,而其身体有SHH的表达,说明缺乏SHH会引起趾头的减少。在本研究中,SHH除了第5天胚龄外,6~10胚龄都是较低表达水平,推测SHH基因可能多在贵妃鸡胚胎早期参与趾的形成,但仍需对SHH基因在贵妃鸡胚胎期的表达情况做进一步研究。R.Schweitzer等[29]利用原位杂交技术对鸡各部位的Gli3基因表达进行了检测,发现Gli3在肢芽早期发育阶段表达,在发育后期表达量下降;Gli3在第4.5天的肢芽间充质高表达。与本试验结果有相同之处。本试验发现,Gli3在贵妃鸡胚胎发育的第9天表达量最高,仍需对胚胎发育5 d之前该基因的表达做进一步的研究。最近研究表明[26],在Gli3基因第5外显子C.480dupC(p.Alal61fs)发生的杂合移码突变会导致多指(趾)并指(趾)的发生。
H.C.Heus等[30]通过基因定位筛选出与人类多趾相关的候选基因还有NOM1、RNF32。NOM1是与MIF4G结构域相关的核仁蛋白,RNF32是泛素连接酶,其催化蛋白质的降解。本试验中,这两个候选基因的相对表达量均为中等水平,是否控制贵妃鸡多趾性状的表达需要进一步的研究。MNX1基因又称为HLXB9(Homeo box 9),汤琳琳[23]研究MNX1在胚胎期的表达研究发现,在胚胎发育的第6~10天各个时期其表达在五趾北京油鸡都显著高于石岐杂鸡,推测MNX1基因可能是调控北京油鸡五趾性状的主效基因。除以上研究外,初芹等[31]研究了北京油鸡多趾性状对体重的影响,发现不同趾型对北京油鸡的体重无显著影响,而腿肌品质与趾型的相关性要大于胸肌。
综上所述,本试验在前人研究的基础上,借助于具有多趾性状这一外貌特征的贵妃鸡为模式动物,对趾的发育机制进行了研究。今后,我们可以通过扩大对不同动物的研究比较,为人的多指(趾)畸形的形成原因及临床治疗提供合理参考。
4 结 论
贵妃鸡多趾性状为常染色体显性遗传,但多趾性状外显率不同,双亲都为多趾的个体后代出现四四趾个体可能是由于基因的遗传修饰造成的,也可能是多基因调控的结果。在贵妃鸡胚胎发育的第5~10天,肢芽组织Lmbr1、RNF32、NOM1、MNX1、Gli3基因分别在第7和第9天出现表达峰值,推测贵妃鸡的多趾性状可能受到Lmbr1、RNF32等基因的共同调控,多趾性状形成的关键时期可能是胚胎发育的第7~9天。
参考文献(References):
[1]BIESECKER L G. Polydactyly: how many disorders and how many genes?[J].AmJMedGenet, 2002, 112(3): 279-283.
[2]BIESECKER L G. Polydactyly: how many disorders and how many genes? 2010 update[J].DevDyn, 2011, 240(5): 931-942.
[3]TEMTAMY S, MCKUSICK V. The genetics of hand malformations[M]. New York: Alan R Liss Inc, 1978: 91-149.
[4]钟文耀, 田文. 桡侧多指畸形的影像学分型[J]. 中国骨与关节杂志, 2017, 6(4): 301-304.
ZHONG W Y, TIAN W. A radiological classification system of radial polydactyly[J].ChineseJournalofBoneandJoint, 2017, 6(4): 301-304. (in Chinese)
[5]黄艳群. 鸡多趾候选基因Lmbr1的克隆和功能研究[D]. 成都: 四川农业大学, 2003.
HUANG Y Q. Cloning and functional analysis of chickenLmbr1 gene——a candidate for polydactyly phenotype[D]. Chengdu: Sichuan Agricultural University, 2003. (in Chinese)
[6]ZHANG Z B, NIE C S, JIA Y X, et al. Parallel evolution of polydactyly traits in Chinese and European chickens[J].PLoSOne, 2016, 11(2): e0149010.
[7]赵献芝, 白小青, 左福元, 等. 多指(趾)基因研究进展[J]. 上海畜牧兽医通讯, 2007(1): 5-7.
ZHAO X Z, BAI X Q, ZUO F Y, et al. Research progress of multi-finger (Toe) genes[J].ShanghaiJournalofAnimalHusbandryandVeterinaryMedicine, 2007(1): 5-7. (in Chinese)
[8]BOND C J. Further observations on polydactyly and heterodactyly in fowls[J].JGenet, 1926, 16(2): 253-256.
[9]PUNNETT R C, PEASE M S. Genetic studies in poultry. VII. Notes on polydactyly[J].JGenet, 1929, 21(3): 341-366.
[10]PITEL F, BERGÉ R, COQUERELLE G, et al. Mapping the Naked Neck (NA) and Polydactyly (PO) mutants of the chicken with microsatellite molecular markers[J].GenetSelEvol, 2000, 32(1): 73-86.
[11]WARREN D C. Inheritance of polydactylism in the fowl[J].Genetics, 1944, 29(3): 217-231.
[12]HUTCHINSON J B. A possible explanation of the apparently irregular inheritance of polydactyly in poultry[J].AmNat, 1931, 65(699): 376-379.
[13]杜晓惠. 鸡多趾候选基因的SNPs及表达差异研究[D]. 成都: 四川农业大学, 2005.
DU X H. Study on the SNPs and expression difference of chicken polydactyly candidate genes[D]. Chengdu: Sichuan Agricultural University, 2005. (in Chinese)
[14]杜志强. 通过基因组扫描定位鸡的重要性状基因座[D]. 北京: 中国农业大学, 2003.
DU Z Q. Mapping quantitative trait loci (QTLs) via genome scanning in Chicken[D]. Beijing: China Agricultural University, 2003. (in Chinese)
[15]黄艳群, 陈文, 邓学梅, 等. 鸡Lmbr1基因内含子多态与趾型的关联[J]. 中国农业科学, 2007, 40(6): 1254-1259.
HUANG Y Q, CHEN W, DENG X M, et al. The association of single nucleotide polymorphisms in ChickenLmbr1 intron with Chicken polydactyl[J].ScientiaAgriculturaSinica, 2007, 40(6): 1254-1259. (in Chinese)
[16]DAVEY M, PATON B, CLELLAND A, et al. 13-P057 allelic imbalance of Shh expression and polydactyly are linked to a SNP in the ZRS region of Lmbr1 in Silkie chickens[J].MechDev, 2009, 126(S1): S212.
[17]邓学梅. 用于鸡基因定位的资源群体的建立和黑色素等质量性状的遗传分析[D]. 北京: 中国农业大学, 2001.
DENG X M. The establishment of resource groups for chicken gene mapping and the genetic analysis of quality traits such as melanin[D]. Beijing: China Agricultural University, 2001. (in Chinese)
[18]YANG K, CHEN Y, GU C, et al. Polydactyly mutation is linked with chromosome 2p region in Beijing fatty chicken[J].AnimGenet, 2013, 44(1): 118-119.
[19]SUN Y F, LIU R R, ZHAO G P, et al. Genome-wide linkage analysis and association study identifies loci for polydactyly in chickens[J].G3 (Bethesda), 2014, 4(6): 1167-1172.
[20]HUANG Y Q, DENG X M, DU Z Q, et al. Single nucleotide polymorphisms in the chickenLmbr1 gene are associated with chicken polydactyly[J].Gene, 2006, 374: 10-18.
[21]曲鲁江, 张泽宾, 陈余, 等. 与鸡多趾性状相关的SNP分子标记及其应用: 中国, CN104694538A[P]. 2015-06-10.
QU L J, ZHANG Z B, CHEN Y, et al. SNP molecular marker related to chicken polydactyly character and application thereof: CN, CN104694538A[P]. 2015-06-10. (in Chinese)
[22]HE C, CHEN Y C, YANG K X, et al. Genetic pattern and gene localization of polydactyly in Beijing fatty chicken[J].PLoSOne, 2017, 12(5): e0176113.
[23]汤琳琳. 五趾北京油鸡趾骨发育相关候选基因表达分析[D]. 上海: 上海交通大学, 2011.
TANG L L. Expression analysis of polydactyly candidate gene of Beijing fatty chicken[D]. Shanghai: Shanghai Jiao Tong University, 2011. (in Chinese)
[24]STURKIE P D. Suppression of polydactyly in the domestic fowl by low temperature[J].JExperZoology, 1943, 93(3): 325-346.
[25]杜晓惠. 鸡多趾候选基因的SNPs及表达差异研究[D]. 成都: 四川农业大学, 2005.
DU X H. Study on the SNPs and expression difference of chicken polydactyly candidate genes[D]. Chengdu: Sichuan Agricultural University, 2005. (in Chinese)
[26]康冉冉, 黄色新, 李杰, 等. GLI3基因新突变导致多指(趾)并指(趾)畸形一家系[J]. 中华医学遗传学杂志, 2017, 34(4): 490-493.
KANG R R, HUANG S X, LI J, et al. A novel mutation of GLI3 gene underlying synpolydactyly in a family[J].ChineseJournalofMedicalGenetics, 2017, 34(4): 490-493. (in Chinese)
[27]BAAS M, POTUIJT J W P, HOVIUS S E R, et al. Intrafamilial variability of the triphalangeal thumb phenotype in a Dutch population: evidence for phenotypic progression over generations?[J].AmJMedGenetA, 2017, 173(11): 2898-2905.
[29]SCHWEITZER R, VOGAN K J, TABIN C J. Similar expression and regulation ofGli2 andGli3 in the chick limb bud[J].MechDev, 2000, 98(1-2): 171-174.
[30]HEUS H C, HING A, VAN BAREN M J, et al. A physical and transcriptional map of the preaxial polydactyly locus on chromosome 7q36[J].Genomics, 1999, 57(3): 342-351.
[31]初芹, 张剑, 张尧, 等. 多趾性状对北京油鸡体重的影响研究[J]. 中国畜牧杂志, 2012, 48(7): 13-15.
CHU Q, ZHANG J, ZHANG Y, et al. Study on the Effect of polydactyly trait on body weight of Beijing fatty chicken[J].ChineseJournalofAnimalScience, 2012, 48(7): 13-15. (in Chinese)
