近30年渤海鱼类种群早期补充群体群聚特性和结构更替*
2018-03-29卞晓东万瑞景金显仕单秀娟关丽莎
卞晓东 万瑞景 金显仕① 单秀娟 关丽莎
(1. 农业部海洋渔业可持续发展重点实验室 山东省渔业资源与生态环境重点实验室中国水产科学研究院黄海水产研究所 青岛 266071;2. 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
渤海属半封闭型的浅海,包括辽东湾、渤海湾和 莱州湾3个海湾,中部存在浅滩地形,其南、北、西三面环陆,仅在东部通过渤海海峡与黄海相连,平均水深 18 m。注入渤海的河流有黄河、海河、滦河和辽河等,其中黄河的年径流量几乎占到了1/2(马伟伟等, 2016)。渤海曾是数十种重要经济渔业生物的繁衍生息地,生物资源量丰富,是我国的“战略鱼仓”;“三湾一河口”是集产卵场、育幼场和渔场于一体的关键栖息地(金显仕等, 2015)。渤海渔业资源种群早期发生量和补充量直接影响到渤海乃至黄海渔业资源动态及其可持续产出(金显仕等, 1998)。
渔业种群早期生活史阶段是其生命中最脆弱、对栖息环境变化敏感性最强的阶段,小规模环境变化也可能会对其资源补充过程产生剧烈影响(Houde,1987)。多重外来干扰作用下,渤海鱼类早期补充过程各个关键环节已随其栖息地(产卵场)生境要素发生不可逆变化或变迁。如大规模环湾围填海工程和临港工业区的建设,造成湿地、滩涂和海湾等鱼虾蟹贝的栖息地大量消失,甚至改变区域性海域的水动力条件和自净能力,进而影响补充群体的分散输运动力学基础;陆源污染物入海总量呈波动式上升趋势,多数近海水域受到不同程度的营养盐、重金属、持续性有机污染物和新型污染物等污染,以“三湾一河口”的污染尤为严重;黄河、辽河、海河等大河径流的减少等,更是严重改变了河口生态环境;另外,大规模海水养殖既直接占用了渔业生物的栖息地和饵料基础,引起滩涂、湿地、海草床等生境的改变,并加剧近海水域富营养化,引发赤潮等生态灾害(金显仕等, 2015)。特别是海洋生物资源的高强度捕捞,破坏了生物资源的自我更新机制,导致生物群落内多重生态位交替失调和渔业资源衰退(朱鑫华等, 2002; 唐启升等, 2013)。
从20世纪50年代开始,黄海水产研究所开展了渤海鱼卵、仔稚鱼的监测调查工作。当时调查对象主要为小黄鱼(Larimichthys polyactis)、带鱼(Trichiurus lepturus)等经济种类;1964~1981年在开展渤海中国对虾(Fenneropenaeus chinensis)产卵场调查时也坚持收集过鱼卵、仔稚鱼资料(姜言伟等, 1988)。此后,尤以 1982~1983年的周年逐月调查(姜言伟等, 1988)和 1992~1993年的周年季度月调查较为全面(万瑞景等, 1998)。进入21世纪,特别是2013~2017年,黄海水产研究所执行的多项科研项目,又在渤海进行了多轮周年季度月调查。30余年渤海各类调查积累了大量翔实的鱼卵仔稚鱼资料。本研究拟基于历史调查资料,结合近年来渤海产卵场调查结果,以1982~1983年周年调查资料为本底,探讨环境-捕捞胁迫下渤海鱼类种群早期补充群体时序分布规律,为保护和修复渔业种群早期生活史关键栖息地提供科学依据。
1 材料与方法
1.1 数据来源
黄海水产研究所1982年4月~1983年5月进行的周年逐月渤海渔业资源增殖基础调查(姜言伟等,1988);1992年8月、10月和1993年2月、5月下旬至 6月上旬 4个航次渔业资源监测调查(万瑞景等,1998);2013~2014 年、2014~2015 年、2015~2016 年以及 2016~2017年进行的多轮周年季度月渔业资源监测调查中,鱼卵仔稚鱼调查均为主要内容之一,其与资源底拖网调查和渔场环境调查同步进行。本研究采用不同调查时期渤海鱼卵仔稚鱼采样站位及时间见表1和图1。
鱼卵、仔稚鱼样品采集用口径80 cm、长270 cm、38 GG筛绢制成大型浮游生物网,采用水平拖网,每站表层拖网10 min,拖曳速度为2 n mile/h。标本用5%海水福尔马林溶液固定。实验室内从大型浮游生物网采集样品中挑出鱼卵和仔稚鱼。样品全样本分析,体视显微镜下经形态学鉴定并反复核实,确认至每一物种单元,并按种类及其发育阶段分别记录个体数。发育阶段包括卵(分裂期、原肠期、胚胎期、孵化期、死卵)和仔稚幼鱼(前期仔鱼、后期仔鱼、稚鱼和幼鱼)。
1.2 鱼卵仔稚鱼生态密度和种类数
本研究所用生态密度为个体数生态密度(Ecological density of number)。不同调查时期鱼卵生态密度取黄渤海区主要产卵季节5~8月数据来计算;仔稚鱼生态密度则取周年调查各月数据来计算。在数据整理过程中,由于水平拖网速度难以严格控制,为便于同质比较和数据质量控制,样品定量分析时以每站每网的实际鱼卵仔稚鱼(粒、尾)作为指标进行比较(姜言伟等, 1988;万瑞景等, 1998)。因不同时期各月调查站位和数量不同,以1982~1983年调查结果作为本底(效应值为1),采用R中的GLM模型(Generalized linear model, 广义线性模型),考虑捕捞努力量的空间分布(渔区)和数据尺度特征(季节),分别对鱼卵或仔稚鱼生态密度进行标准化,以构建资源丰度指数(Abundance index)指示鱼卵、仔稚鱼生态密度(官文江, 2015)。鱼卵或仔稚鱼种类数则为考察期出现鱼卵或仔稚鱼物种数。
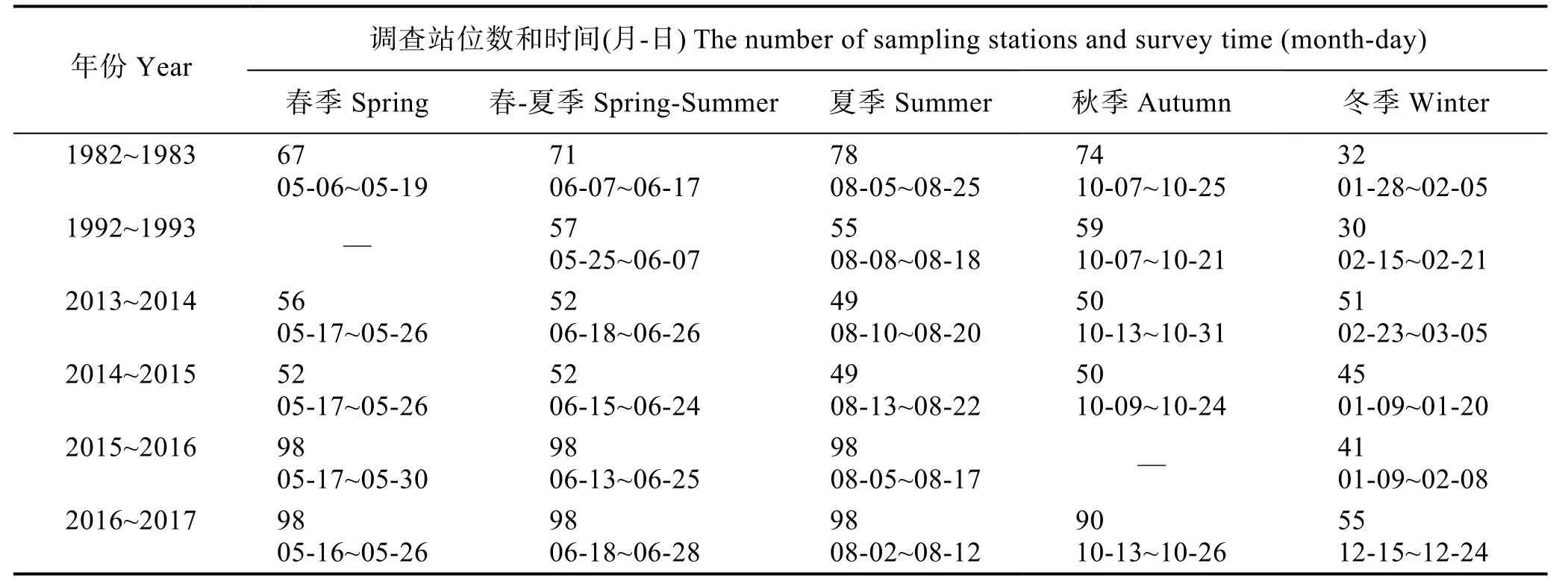
表1 不同调查时期渤海鱼卵仔稚鱼采样时间、区域和站位数Tab.1 Survey time, area and number of sampling stations in the Bohai Sea in different survey periods
1.3 不同调查时期各季节鱼卵仔稚鱼优势种类和重要种类
运用 Pinkas相对重要性指数[Index of relative importance, IRI,(1)](Pinkaset al, 1971)对每航次调查过程中出现鱼卵和仔稚鱼种类组成分别进行分析,以确定鱼卵或仔稚鱼优势种类(Dominant species)、重要种类(Important species)和主要种类(Main species)的成分;

式(1)中N%为每航次调查过程中某种鱼卵或仔稚鱼个体数量占鱼卵或仔稚鱼总量比例;W%为每航次调查过程中某种鱼卵或仔稚鱼生物量比例。由于鱼卵和仔稚幼鱼个体都很小,因此不考虑生物量,只考虑个体数量这一因素,IRI的计算公式可以简化为式(2)(卞晓东等, 2010)。取IRI值大于1000种类定义为鱼卵或仔稚鱼优势种;鱼卵重要种类 IRI值为500~1000,仔稚鱼重要种类IRI值为350~1000。
1.4 不同调查时期各季节鱼卵仔稚鱼物种多样性水平及其种类更替
采用 α多样性测度方法—香农-威纳指数[Shannon-Wiener index,H′(3)]和辛普森多样性指数[Simpson's diversity index,DS(4)]研究渤海不同调查时期各季节鱼卵和仔稚鱼物种多样性;采用β多样性测度方法—Jaccard群落种类组成相似性指数[Coefficient of community, CC (5)]比较各年间鱼类早期补充群体(鱼卵仔稚鱼种数合并计数)种类相似程度(Whittaker, 1972)。
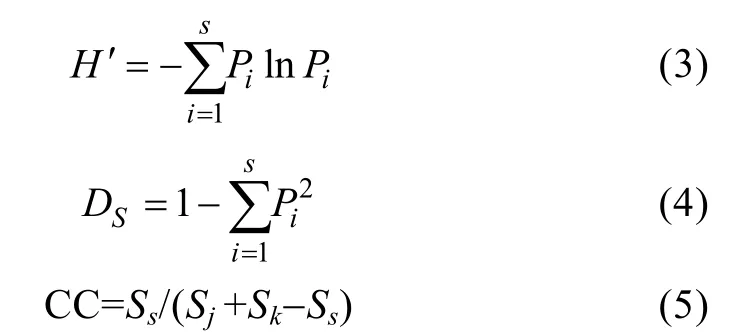
式(3)和式(4)中S为不同调查时期各季节鱼类早期补充群落中出现的鱼卵或仔稚鱼种类数,Pi为群落中第i种鱼卵或仔稚鱼所占鱼卵或仔稚鱼总量的个体比例。式(5)中Ss为 2个比较年份间鱼类早期补充群体共有种类数,Sj和Sk为2个比较年份各自拥有种数。当 CC为 0~0.25时,群落之间极不相似;当 CC为0.25~0.50时,群落之间中等不相似;当 CC为0.50~0.75时;群落之间中等相似;当CC为0.75~1.00时,群落之间极相似。
1.5 产卵亲体适温类型和栖所类型分析
根据鱼类区系分类的有关文献索引(田明诚等,1993)将产卵亲体的适温类型划分为暖温种(Warm temperate species, WT),暖水种(Warm water species,WW)和冷温种(Cold temperate species, CT)。根据刘静等(2011)将渤海产卵亲体栖所类型划分为大陆架浅水底层鱼类(Continental shelf demersal fish, CD),大陆架岩礁性鱼类(Continental shelf reef-associated fish, CRA),大陆架浅水中上层鱼类(Continental shelf pelagic-neritic fish, CPN),大陆架浅水中底层鱼类(Continental shelf benthopelagic fish, CBD),大陆架大洋洄游性中上层鱼类(Oceanic pelagic fish, OEP)和大洋深水底层鱼类(Oceanic bathydemersal fish, OMP)。

图1 不同调查时期渤海鱼卵、仔稚鱼采样区域和站位Fig.1 Survey area and sampling stations in the Bohai Sea in different survey periods
2 结果
2.1 渤海各调查时期不同季节鱼卵、仔稚鱼种类数和生态密度
近30年来,渤海各调查季节鱼卵种类数(冬季除外)和资源生态密度与 1982~1983年相比均呈现不同程度下降,在春和春夏季均呈现先降后升格局(图2a,图2 b)。渤海主要产卵季节(5月、6月和8月)鱼卵种数由1982~1983年的38种,逐次下降到2015~2016年的 18种(不足 1982~1983年的 1/2),在 2016~2017年调查中鱼卵种类数小幅回调至 26种(图3a)。1982~1983年鱼卵资源丰度指数为1,1992~1993年跃升为3.09,进入2010年后,鱼卵资源丰度指数急剧下跌,近年来一直维持在0.05~0.09水平(图3a),当前鱼卵资源丰度不足1982~1983年的1/10。渤海各调查时期不同季节出现仔稚鱼种类数和资源生态密度在春季、春夏季、秋季均呈现先降后升格局;而在冬季则均呈现明显的增加趋势(图2c, 图2d)。渤海仔稚鱼种数则由1982~1983年的 52种(其中虾虎鱼类 3种),下降到2014~2015年的25种(种类数不足1982~1983年的1/2),随后上升至2016~2017年的 40种(其中虾虎鱼类9种;图3b)。仔稚鱼资源丰度指数1982~1983年为 1,在 1992~1993年指数上升为 1.087,此后仔稚鱼资源丰度指数急剧下跌,近年来则维持在低位(0.679~ 0.761;图3b),当前仔稚鱼资源丰度不足1982~1983 年的 3/4。
2.2 渤海各调查时期不同季节鱼卵仔稚鱼优势和重要种类

图2 渤海各调查时期不同季节鱼卵、仔稚鱼种类数和生态密度概况Fig.2 Number of species and ecological density to fish eggs and larvae profiles along the survey season across different survey times
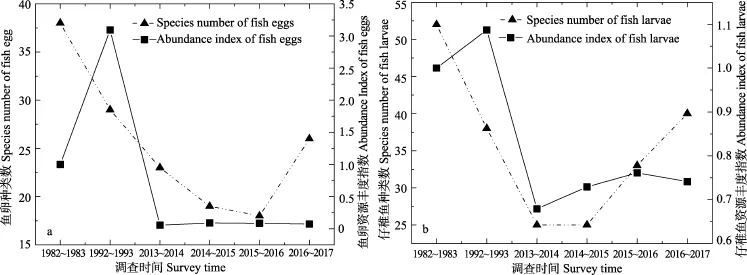
图3 渤海各调查时期不同季节鱼卵(a)仔稚鱼(b)资源丰度指数和种类数Fig.3 Abundance Index and species number of fish eggs (a) and larvae (b)
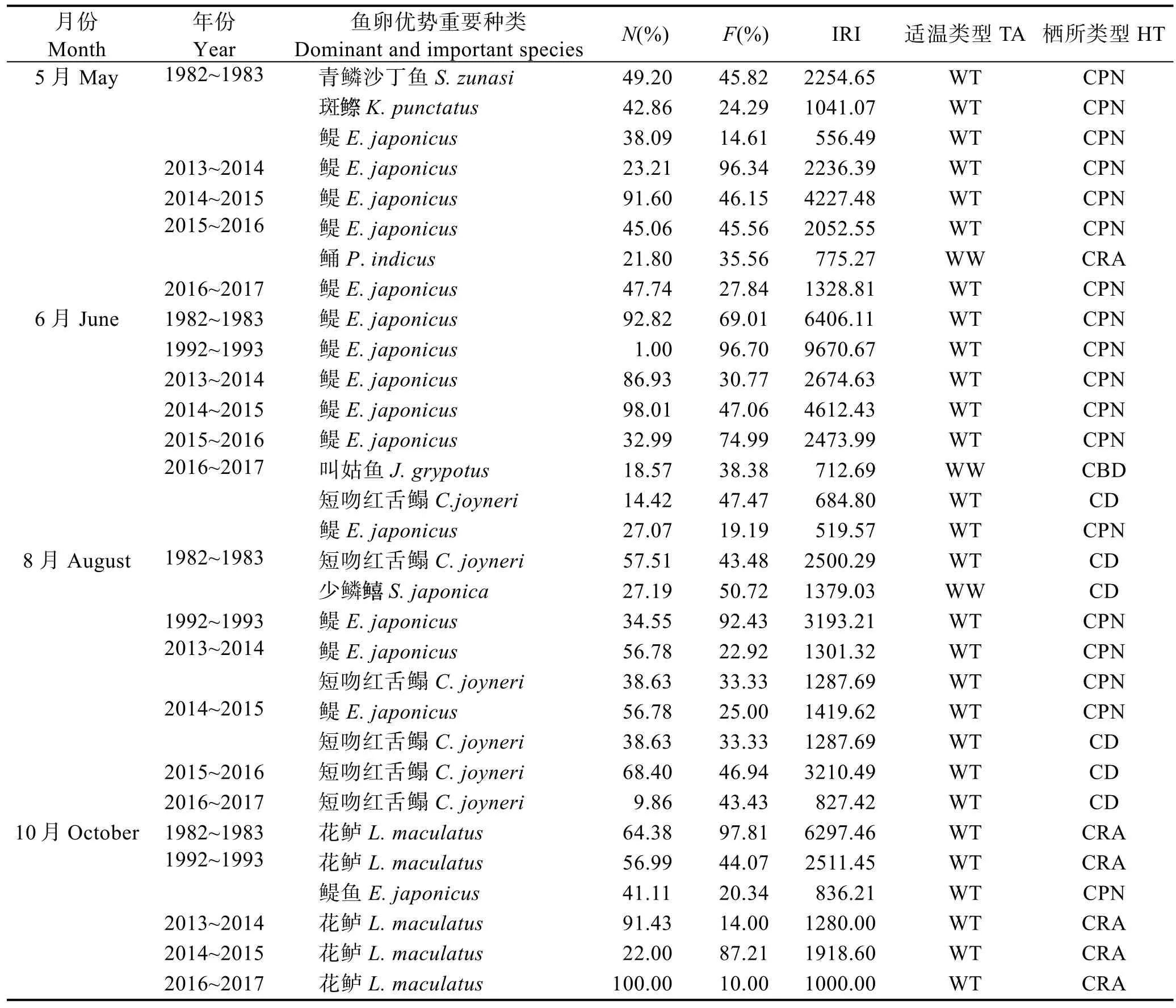
表2 渤海各调查时期鱼卵优势(IRI>1000)和重要(IRI>500)种类及其亲体适温和栖所类型Tab.2 The dominant (IRI>1000) and important (IRI>500) species of fish eggs with temperature adaptation (TA)and habitat types (HT) to their adult fish in the Bohai Sea in recent 30 years
各调查季节鱼卵优势种变化不明显,在增温季节(5月、6月)优势种类为暖温性CPN鱼类,1982~1983年为青鳞沙丁鱼(Sardinella zunasi)、斑(Konosirus punctatus)和鳀鱼(Engraulis japonicas) 3 种,1992~1993年是鳀鱼大暴发年,鳀鱼优势度显著提高,成为唯一优势种类;进入2010年,仍然以鳀鱼为优势种类(2013~2015年),2016~2017年CD和CBD鱼类如鲬(Platycephalus indicus)、叫姑鱼(Johnius grypotus)和短吻红舌鳎(Cynoglossus joyneri)优势度均显著提高与鳀鱼共同成为优势种类(表2)。在夏季 8月,表层水温最高,水平温差小,优势种类为暖温性和暖水性的 CD 鱼类,1982~1983年为短吻红舌鳎和少鳞(Sillago japonica),1992~1993年,鳀鱼是唯一优势种类;进入2010年,过渡为以鳀鱼和短吻红舌鳎(2013~2015年)为主,近年来(2015~2017年)短吻红舌鳎成为单一优势种类(表2)。在降温季节(10月),优势种类为 CRA鱼类,如 1982~1983年为花鲈(Lateolabrax maculatus),随后在 1992~1993年,鳀鱼优势度上升与花鲈同为优势种,近年来又恢复至以花鲈为单一优势种类(表2)。
各调查季节仔稚鱼优势种变化幅度超过鱼卵,在增温季节(5月、6月)优势种以暖温性CPN鱼类为主,1982~1983年和 1992~1993年为鳀鱼、斑和青鳞沙丁鱼,近年来暖温性CPN鱼类(Liza haematocheila)、CD 鱼类矛尾虾虎鱼(Chaeturichthys stigmatias)和冷温性CD鱼类许氏平鲉(Sebastes schlegeli)优势度迅速上升,在某些年份成为优势种类(表3)。在8月,优势种类由暖温性 OEP鱼类尖嘴扁颌针鱼(Strongylura anastomella)变为1992~1993年的鳀鱼和斑,近年来暖水性CPN鱼类白氏银汉鱼(Allanetta bleekeri)和暖温性CPN鱼类沙氏下鱼(Hyporhamphus sajori)优势度显著提高,逐渐变成优势种类(表3)。降温季节(10月),在强厄尔尼诺年1982~1983年(Wuet al, 2016)优势种为暖水性 CD鱼类少鳞,1992~1993年后优势种变为鳀鱼和 CRA鱼类花鲈(表3)。冬季(1~3月)渤海表层水温最低,在强厄尔尼诺年 1982~1983年冬季(暖冬; Wuet al, 2016)优势种类为暖温性CRA鱼类花鲈;其余年份冷温性 CD鱼类大泷六线鱼(Hexagrammos otakii)和方氏云鳚(Enedrias fangi)成为优势种类(表3)。

表3 渤海各调查时期仔稚鱼优势(IRI>1000)和重要(IRI>350)种类及其亲体适温和栖所类型Tab.3 The dominant (IRI>1000) and important (IRI>350) species of fish larvae with temperature adaptation (TA)and habitat types (HT) to their adult fish in the Bohai Sea in recent 30 years
2.3 渤海鱼卵仔稚鱼物种多样性水平和种类更替
多轮周年调查结果显示,鱼卵物种多样性水平(H′和DS)季节变化特征为在升温季节春夏季较高,在降温季节秋冬季较低,冬季(1~2月)最低;H′和DS呈“双峰分布”,分别在春季(5月)和夏季(8月)达到峰值,春夏季(6月)物种多样性水平略低(图4a, 图4c)。各调查时期,春季鱼卵H′范围为 0.20~1.95,峰值为1982~1983年的 1.95,1992~1993年急剧下降至 0.22,并在2013~2014年至谷值0.20;自2014~2015年始呈现上升趋势,在2016~2017年升至1.4左右。春夏季鱼卵H′范围为 0.13~1.84,其中 1982~1983 年为 0.42,1992~1993年降至 0.22,并在 2014~2015年至谷值0.12;自 2015~2016年始呈现快速上升趋势,2016~2017 年至峰值 1.84。夏季H′范围为 0.38~1.22,其中1982~1983年为1.21,1992~1993年急剧下降至谷值 0.38外;此后逐渐上升,于 2016~2017年达峰值 1.23。秋季H′范围为 0~0.78,其中 1982~1983年为 0.12,峰值在 1992~1993年的 0.78,谷值则在2016~2017年的0。冬季调查除2016~2017年外没有采集到鱼卵(图4a)。

表4 渤海各调查时期鱼类种群早期补充群体种类组成年间β相似性指数矩阵Tab.4 Matrix of the β similarity index (CC) of ichthyoplankton communities in the Bohai Sea between different survey periods
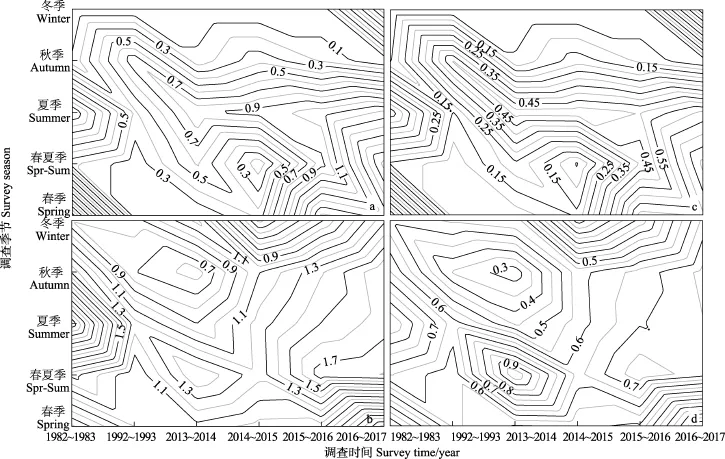
图4 渤海各调查时期不同季节鱼卵和仔稚鱼香农-威纳和辛普森多样性指数Fig.4 Shannon-Wiener and Simpson's diversity index to fish eggs and larvae profiles along the survey season across different survey times
仔稚鱼H′和DS在升温季节(5~8月)逐次升高,降温季节(10月至翌年2月)逐次降低。仔稚鱼H′和DS呈“单峰分布”,夏季达到全年峰值,谷值在春季(图4b,图4d)。各调查时期,春季H′范围为0.15~1.06,其变化趋势与鱼卵相反,其中1982~1983年为次谷值0.57,峰值为1992~1993年的1.00,此后在2013~2016年间均维持在0.95以上,而2016~2017年为H′值仅为0.15。春夏季H′范围为0.55~1,其中1982~1983年为0.65,谷值为 1992~1993年的 0.56,近年来(2013~2017年)由于虾虎鱼科(Gobiidae)鱼类被鉴定种类大幅增加(至2016~2017年被鉴定种类为9种),H′出现较大幅度上升,并在2016~2017年升至峰值1.81。夏季仔稚鱼H′范围均较高,在0.89~2.59之间,其中峰值出现在1982~1983年的2.59,1992~1993年降至1.24,并在2013~2014年跌至谷值 0.89,近年来(2014~2017年)出现增长趋势,2016~2017年H′上升至 1.65左右。秋季仔稚鱼H′范围均较高,在 0.54~1.69之间,其中1982~1983 年值为 1.48,1992~1993 年降至 0.70,并在2013~2014年回落至谷值0.54,近年来又呈现回调趋势,并在 2016~2017年升至峰值 1.69。冬季H′范围为 0.15~1.06,年间变异比较大,1982~1983年H′为 0.80,峰值出现在 2016~2017年的 1.27,谷值为2014~2015 年的 0.16(图4b)。
渤海各调查时期鱼类种群早期补充群体种类更替现象明显。1982~1983年和1992~1993年鱼类早期补充群体CC值位于0.50~0.75区间,群落中等相似;而与2013~2017年各年间鱼类早期补充群体CC值均位于 0.25~0.50区间,群落间中等不相似(表4)。1992~1993年 CC 值与 1982~1983 年、2013~2014 年和 2014~2015各年间鱼类早期补充群体 CC值位于0.50~0.75区间,群落中等相似;而与 2015~2016年和 2016~2017年CC值均位于 0.25~0.50区间,群落间中等不相似。近年来种类更替率呈现明显加快趋势,即便是在最近调查年份2013~2014年,其鱼类早期补充群体CC值与2015~2016年为0.47,位于0.25~0.50区间,群落间中等不相似(表4)。渤海鱼类种群早期补充群体种类组成年间β相似性指数聚类分析结果显示群落相似性程度依次为2015~2016年,2016~2017年,2013~2014年,2014~2015年,1982~1983年和 1992~1993年。
2.4 渤海各调查时期产卵亲体栖所类型和适温类型种类数
不同调查时期渤海鱼类早期补充群体对应亲体各栖所类型均以CD种数最高,其次为CPN、CBD、CRA和OEP,而OMP种类年间波动较大(图5)。自1982~1983年起至2014~2015年,CD和CBD鱼类种类数由41种急剧下降至19种,占总种类数百分比也由1982~1983年的最高67.21%,下降至2014~2015年的54.28%;2016~2017年种数又上升至34种,所占比率恢复至 64.15%(图5)。CPN 种类数由 1982~1983年16种,逐渐下降至2014~2015年的11种,占总种类数百分比与之相反呈现上升趋势,由1982~1983年的26.23%上升至 2014~2015年的 31.43%;2016~2017年CPN种类数小幅上升至12种,占总种类数百分比则下降至22.64%。自1982~1983年起CRA种类数基本保持稳定(2015~2016年除外),占总种类数百分比率呈现上升趋势,近年来出现小幅回调。OEP(尖嘴扁颌针鱼)和 OMP种类(黄Lophius litulon)年间波动较大。不同调查时期渤海鱼类早期补充群体对应亲体各适温类型均以暖温性种数最高,暖水性种类次之,冷温性种类最低,没有出现冷水性鱼类(图6)。不同调查时期各适温类型鱼类年间占总种类数百分比率波动不大。暖温性鱼类占总种类数百分比在各调查时期均超过50%,其中1992~1993年暖温性鱼类占总种类数百分比为62.22%最高,2016~2017年暖温性鱼类占总种类数百分比最低为50.94%。暖水性鱼类占总种类数百分比在 2015~2016年最低为23.68%。冷温性鱼类占总种类数百分比则呈上升趋势,由1982~1983年的11.48%,上升至2016~2017年的 20.75%(图6)。
3 讨论
自20世纪90年代以来我国乃至全球范围内,渔业资源普遍衰退,资源结构发生了很大的变化,这在渤海尤为突出,表现为渔获小型化和低质化,种类交替明显,重要渔业资源已不能形成渔汛,对渤黄海渔业的支持功能日益衰退(Tanget al, 2003; Shanet al,2016; 李忠义等, 2017)。渔业种群的变动主要由补充量变化驱动,过度捕捞和早期补充不足是渔业资源衰退的两个重要原因。30余年鱼卵仔鱼调查一直沿用相同调查网具和方法,样品鉴定分析则采用参考资料和分类标准一致的传统形态学分类方法。作为渔业资源的早期补充资源,与1982~1983年调查相比当前各季节(冬季除外)鱼卵和仔稚鱼种类数和资源丰度指数均已呈现较大程度下降。当前鱼卵种类数仅为20世纪80年代的1/2左右,资源丰度不足彼时的1/10;仔稚鱼种类数和资源丰度仅为20世纪80年代的3/4左右。各调查时期相同季节鱼卵优势种变化不明显,春季和春夏季优势种类由小型 CPN鱼类如青鳞沙丁鱼、斑和鳀鱼变为当前的鳀鱼(近年来小型 CD和CBD鱼类如叫姑鱼和短吻红舌鳎优势度在春夏季显著提高,与鳀鱼共同成为优势种类);夏季由暖温性和暖水性的小型CD鱼类如短吻红舌鳎和少鳞变为当前短吻红舌鳎;秋季优势种无明显变化。仔稚鱼优势种变化幅度超过鱼卵,春季和春夏季由小型 CPN鱼类斑、青鳞沙丁鱼和鳀鱼变为 CPN鱼类鱼;夏季由OEP鱼类尖嘴扁颌针鱼变为小型CPN鱼类沙氏下鱼;秋季由小型CD鱼类少鳞变为小型CPN鱼类鳀鱼;冬季由CRA鱼类花鲈变为CD鱼类大泷六线鱼和方氏云鳚。底层重要经济种类早期补充群体优势度急剧下降。鱼类早期补充群体种类更替现象明显,近年来种类更替率呈现明显加快趋势。各调查时期相同季节鱼卵和仔稚鱼物种多样性水平、各适温类型产卵亲体种数、各主要栖所类型产卵亲体种类数均呈现先降后升变动趋势。全年综合陆架浅水中上层鱼类种数所占比例升高,中底层和底层鱼类所占比例有所下降。

图5 渤海鱼类种群早期补充群体对应亲体各栖所类型年间种数比较Fig.5 Comparison of the number of species to different habitat type in parent of the recruitment stock between different survey periods

图6 渤海鱼类种群早期补充群体对应亲体各适温类型年间种类数比较Fig.6 Comparison of the number of species to different temperature adaptation type in parent of the recruitment stock between different survey periods
3.1 环境变化对渤海鱼类种群早期补充群体群聚特性和结构更替影响的认识
近海环境变化对渔业生物种群补充过程的影响及其资源效应是海洋渔业科学前沿领域的基础科学问题。挪威著名鱼类学家Hjort(1914)提出了渔业资源评估最早的3个学说之一“波动论”,指出鱼类早期生活阶段仔幼鱼的成活率取决于鱼类初次摄食时开口饵料的丰度和栖息地海流、水温、盐度等非生物因素,环境条件是形成鱼类世代变动的主要原因。Cushing (1975、1990)将 Hjort(1914)理论进行了扩展,提出了“匹配与不匹配”假说(“match-mismatch” hypothesis),指出鱼类仔稚鱼及其饵料生物在时间和空间上的一致性与否是决定补充强度的关键因素。鱼类在繁殖期间对环境要求较为严格,需选择最适于其生殖活动并对卵子和仔鱼存活最为有利环境栖息,并在长期演化过程中形成不同的生活史策略来提高其早期补充幼体与饵料生物的时空匹配性从而达到快速生长,以提高幼体成活率(Pecket al, 2012)。渤黄海鱼类区系主要品种具低盐河口近岸海区产卵特性(朱鑫华等, 1994)。渤黄海河口和近岸海区海况变化复杂,主要存在低盐和高盐性质2种水系(Chenet al, 2009),这2种水系锋带(即盐度水平梯度大的混合区)在适温条件下往往成为多数鱼群最密集产卵中心亦即鱼卵、仔稚鱼密集区,且鱼群集散程度在很大程度上取决于盐度水平梯度的大小,亦即水系锋带是否显著;因适宜环境在锋带区范围很小,因而造成鱼群(亦或鱼卵仔鱼)高度集中并随着锋区而移动(邱道立等, 1965; Weiet al, 1992; 万瑞景等, 2008、2014)。
在捕捞强度基本保持不变的前提下,全球变化下ENSO(El Niño/La Niña-Southern Oscillation)等气候事件会通过影响主要水系的水动力条件(如温盐场、流场)和营养基础等造成东西太平洋两侧海域主要经济鱼类资源量剧烈变动(金显仕等, 2015)。这种影响在渤海表现也比较明显,如自1960~1997年,渤海表层盐度升高了2.82 PSU,气温升高了0.92℃,表层水温上升了 0.41℃(方国宏等, 2002);且受气候因素时空变异和人类活动影响,海区内鱼类赖以产卵繁殖的黄河入海径流量自1980~2002年,一直呈现负增长,并在 1997~2002年间维持在历史底位;2002年后入海径流量呈现一定程度回升,但是自1978年以来入海径流量显著线性降低趋势仍未改变(Fanet al, 2008;Renet al, 2015)。从生物物候学角度,作为初级生产者和鱼类早期补充群体饵料的硅藻类春季水华发生时间及其季节性周期多数保持固定时间节点,其主要由白昼时长和光照强度决定而并非由水温调节(Eilertsenet al, 1995; Mcquoidet al, 2010)。鱼类早期补充群体(季节性浮游生物)其发生和生命周期时间节点则主要受温度调节的生理反应控制(Edwardset al,2004),随着近年来海水温度的不断上升,其生理周期(胚胎发育、仔鱼孵化和开口摄食等)将提前,在变暖趋势不变情景下其将继续持续下去(Edwardset al, 2004)。这可能造成鱼类早期补充群体与饵料生物的错配,从而导致早期补充能力下降进而出现渔业种群的衰退,即上行控制(Bottom-up control)。此外,自20世纪50年代始,特别是近20年来渤海区不断出现的暖冬现象(Wuet al, 2017)使得近年来冬季产卵繁殖鱼类仔稚鱼种类和资源丰度指数呈现上升趋势(因冬季产卵鱼类主要为产沉性卵或卵胎生,采用传统鱼卵仔稚鱼调查取样方法无法获取,从而使得冬季仔稚鱼优势种变化幅度超过鱼卵)。尽管多数鱼类早期浮游幼体对全球气候变化有响应,但不同鱼种响应程度不同,为准确评估近海环境变化对渔业种群补充过程的影响,应当进一步了解气候变化驱动的非生物(海水温度)和生物因素(饵料生物的匹配与不匹配)对特定鱼种早期生活史的影响,并厘清其在不同类群以及相同类群不同栖息地间的影响程度(Pecket al, 2012)。
3.2 捕捞对渤海鱼类种群早期补充群体群聚特性和结构更替影响的认识
捕捞主要通过对目标鱼种的开发利用以及栖息地底质扰动,导致渔业生物资源量、群落结构和群体生物学特征改变,其对各鱼种影响程度则主要取决于相应种类生活史特征、营养关系及其栖息地环境受损程度(Bianchiet al,2000)。自2000年以来补充型过度捕捞导致的生殖群体资源量降低(Zhaoet al, 2003),小型化、低龄化和性成熟提前(Tanget al, 2007)及其亲体效应(后代成活率降低; Wanet al, 2012)等成为解释渔业种群早期补充不足和整个渔业种群衰退的主要因素。在高强度捕捞压力下(渤海内捕捞强度自1950年以来增长了近40倍;Shanet al, 2013)渤海鱼类资源的早期补充能力较 20世纪 80年代已急剧下降,伴随着鱼类早期补充群落结构的变化,年间和季节间种类数密度也随之降低,优势种、优势度以及个体生态密度降低,特别是底层重要经济种类早期补充群体优势度呈现急剧下降趋势,如带鱼、真鲷(Pagrosomus major)、褐牙鲆(Paralichthys olivaceus)、半滑舌鳎(Cynoglossus semilaevis)、棘头梅童鱼(Collichthys lucidus)、小黄鱼等,有些种类则因产卵群体过度捕捞或栖息地丧失在近年调查中绝迹如鳓鱼(Ilisha elongata)等。同在渔业资源衰退背景下,仔稚鱼优势种变化幅度超过鱼卵,究其原因可能是捕捞(捕捞结构和捕捞强度的时空差异)等人类活动影响下具有特殊繁殖生物学特性的鱼类,如具卵膜丝卵鱼类(虾虎鱼科矛尾虾虎鱼、科Hemiramphidae沙氏下鱼、银汉鱼科Atherinidae白氏银汉鱼)和卵胎生鱼类(鲉科Scorpaenide的许氏平鲉)等仔稚鱼自2010年以来在特定调查季节成为优势种类。而此类鱼类鱼卵由于其特殊繁殖生物学特性,采用传统调查取样方法无法获取,从而不能反映在鱼卵种类的优势度组成上。而物种多样性指数的改变则主要是由物种均匀度和丰富度改变,在较高捕捞压力下,群落物种多样性指数反而有时会呈现上升现象(Bianchiet al,2000)。这在渤海春、夏季鱼卵仔稚鱼物种多样性水平变化趋势中表现也较明显,其峰值均出现在高捕捞压力资源严重衰退背景下的2016~2017年。而在某些情境下多样性指数改变主要是由于调查方法的改进和物种鉴定方法提升而引起(Bianchiet al, 2000),如自2010年以来因海区内鉴定的虾虎鱼类仔稚鱼种类数增加也使得仔稚鱼物种多样性水平计算结果的准确性有所提高。
全球范围内中上层小型鱼类是海洋生态系统的主要组成部分,支撑了重要的渔业(Maynou, 2014),这在渤海也无例外。鳀鱼、斑、青鳞沙丁鱼、沙氏下鱼和白氏银汉鱼等中上层小型鱼类鱼卵或仔稚鱼也是各调查年份鱼类早期补充群体主体成分。以鳀鱼为例,鳀鱼主要摄食浮游动物,如中华哲水蚤(Calanus sinicus)、太平洋磷虾(Euphausia pacifica)等,又是40多种高营养层次重要经济鱼类,如蓝点马鲛(Scomberomorus niphonius)、小黄鱼、带鱼等主要饵料(Weiet al, 1992),在食物网中是将浮游动物转化为高营养级鱼类重要中间环节,对海洋生态系统的能量流动和转换有着重大的作用,是海洋生态系统中的关键物种(Tanget al,2005)。在黄渤海区自20世纪90年代以来开始兴起的鳀鱼大规模开发利用使得该种群遭受的捕捞压力急剧增加,其资源出现了严重的衰退现象,其繁殖生物学特性对种群长期承受巨大的捕捞压力产生适应性响应,表现为卵子卵径明显变小,自然死亡卵子所占的比例呈现较大幅度的上升等(Wanet al, 2012)。同时整个渤海生态系统中由于对顶级捕食者的高强度持续过度捕捞,食物网变得愈来愈简单,低营养级的物种成为控制渔业生态系统能量流动的主要种类,优势种群更替和饵料生物更替而产生的下行控制作用(Top-down control)影响整个生态系统生物资源的可持续产出(Tanget al, 2003)。随着下行控制影响时间的推移,以次级生产力(浮游动物)作为主控因子的丰腰控制(Wasp-waste control)持续影响生态系统的可持续产出,并最终导致各营养层级产出年际变化呈现出不稳定和无序(Tanget al, 2003),这可能是不同调查时期生态习性和食性各异的鱼类早期补充群体的种类更替明显并且近年来呈现加快趋势的原因。
30余年调查结果显示,鱼卵和仔稚鱼种类数和资源丰度指数、物种多样性水平等均在2013~2015年间跌至谷值。经分析除捕捞和气候变化因素外,发生于2011年6月蓬莱19-3油田的两起严重溢油事故(周利,2014; 李忠义等, 2017)对海域海洋生态环境的持续影响或许是主要原因之一。渔业种群早期补充群体的时序分布规律既决定于生物本身生理发育阶段间行为适应因素,又受制于水域温盐度场势季节消长趋势等环境要素,这种生物的行为特征与生态环境变化的适应性调节,直接影响到渔业种群早期补充群体空间分布的可塑性动态格局;任何一种控制机制如“上行控制”、“下行控制”或“丰腰控制”均不足以直接或清楚解释其长期变化规律,其时序变化特征则是对人类活动和环境变化的综合反映。
致谢:中国水产科学研究院黄海水产研究所资源室的前辈和同事协助取样和数据收集,谨致谢忱。
Bianchi G, Gislason H, Graham K,et al. Impact of fishing on size composition and diversity of demersal fish communities.ICES Journal of Marine Science, 2000, 57(3): 558–571
Bian XD, Zhang XM, Gao TX,et al. Category composition and distributional patterns of ichthyoplankton in the Yellow River estuary during spring and summer 2007. Journal of Fishery Sciences of China, 2010, 17(4): 815–827 [卞晓东,张秀梅, 高天翔, 等. 2007年春、夏季黄河口海域鱼卵、仔稚鱼种类组成与数量分布. 中国水产科学, 2010, 17(4):815–827]
Chen CTA. Chemical and physical fronts in the Bohai, Yellow and East China Seas. Journal of Marine Systems, 2009,78(3): 394−410
Cushing DH. Marine ecology and fisheries. Cambridge, UK:Cambridge University Press, 1975
Cushing DH. Plankton production and year-class strength in fish populations: An update of the match/mismatch hypothesis.Advances in Marine Biology, 1990, 26(6): 249–293
Edwards M, Richardson AJ. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature, 2004,430(7002): 881–884
Eilertsen HC, Sandberg S, Tellefsen H. Photoperiodic control of diatom spore growth: A theory to explain the onset of phytoplankton blooms. Marine Ecology Progress Series,1995, 116: 303–307
Fang GH, Wang K, Gou FY,et al. Long-term changes and interrelations of annual variations of the hydrographical and meteorological parameters of the Bohai Sea during recent 30 years. Oceanologia et Limnologia Sinica, 2002, 33(5):515–525 [方国洪, 王凯, 郭丰毅, 等. 近30年渤海水文和气象状况的长期变化及其相互关系. 海洋与湖沼, 2002,33(5): 515–525]
Fan H, Huang H. Response of coastal marine eco-environment to river fluxes into the sea: A case study of the Huanghe(Yellow) River mouth and adjacent waters. Marine Environmental Research, 2008, 65(5): 378–387
Jiang YW, Wan RJ, Chen RS. Investigation of eggs and larvae of osteichthyes in the Bohai Sea. Marine Fisheries Reseach,1988(9): 121–149 [姜言伟, 万瑞景, 陈瑞盛. 渤海硬骨鱼类鱼卵、仔稚鱼调查研究. 海洋水产研究, 1988(9): 121–149]
Jin XS, Tang QS. The structure, distribution and variation of the fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 1998, 5(3): 18–24 [金显仕, 唐启升. 渤海渔业资源结构、数量分布及其变化. 中国水产科学,1998, 5(3): 18–24]
Jin XS, Dou SZ, Shan XJ,et al. Hot spots of frontiers in the research of sustainable yield of Chinese inshore fishery.Progress in Fishery Sciences, 2015, 36(1): 124–131[金显仕, 窦硕增, 单秀娟, 等. 我国近海渔业资源可持续产出基础研究的热点问题. 渔业科学进展, 2015, 36(1):124–131]
Guan WJ. Application ofRprogram in marine fisheries. Beijing:China Ocean Press, 2015, 224–229 [官文江.R语言在海洋渔业中的应用. 北京: 海洋出版社, 2015, 224–229]
Hjort J. Fluctuations in the great fisheries of northern Europe,viewed in the light of biological research. Rapports et Procès-Verbaux des Réunions du Conseil Permanent International Pour L’Exploration de la Mer, 1914, 20: 1–228 Houde, ED. Fish early life dynamics and recruitment variability.American Fisheries Society Symposium, 1987, 2: 17–29
Liu J, Ning P. Species composition and faunal characteristics of fishes in the Yellow Sea. Biodiversity Science, 2011, 19 (6):764–769 [刘静, 宁平. 黄海鱼类组成、区系特征及历史变迁. 生物多样性, 2011, 19 (6):764–769]
Li ZY, Wu Q, Shan XJ,et al. Interannual variations in fish community structure in the Bohai Sea. Journal of Fishery Sciences of China, 2017, 24(2): 403–413 [李忠义, 吴强,单秀娟, 等. 渤海鱼类群落结构的年际变化. 中国水产科学, 2017, 24(2): 403–413]
Ma WW, Wan XQ, Wan K. Interannual variation and formation of wind-driven currents of Bohai Sea in winter. Oceanologia et Limnologia Sinica, 2016, 47(2): 295–302 [马伟伟,万修全, 万凯. 渤海冬季风生环流的年际变化特征及机制分析. 海洋与湖沼, 2016,47(2): 295–302]
Maynou F, Sabatés A, Salat J. Clues from the recent past to assess recruitment of Mediterranean small pelagic fishes under sea warming scenarios. Climatic Change, 2014,126(1–2): 175–188
Mcquoid MR, Hobson LA. Diatom resting stages. Journal of Phycology, 2010, 32(6): 889–902
Peck MA, Huebert KB, Llopiz JK. Intrinsic and extrinsic factors driving match-mismatch dynamics during the early life history of marine fishes. Advances in Ecological Research,2012(47): 177–302
Pinkas L, Oliphant MS, Iverson I. Food habits of albacore,bluefin tuna and bonito in California waters. California Department of Fish and Game, Fish Bulletin, 1971(152):1–105
Qiu DL, Liu XS, Wang ZX. Migration of the small yellow croakerLarimichthys polyactisin the Bohai Sea and Yellow Sea in relation to the hydrographic conditions. InAnthology sequel of papers on marine fishery resources(Editorial Committee of the China Scientific Council on the Marine Fisheries Resources 1962, eds). Beijing: Agriculture Press,1965, 43–35 [邱道立, 刘效舜, 王遵孝. 渤、黄海小黄鱼的洄游与水文环境的关系. 海洋渔业资源论文选集续集(1962年海洋渔业资源学术会议论文编审委员会). 北京:农业出版社, 1965, 43−55]
Ren H, Li G, Cui L,et al. Multi-scale variability of water discharge and sediment load into the Bohai Sea from 1950 to 2011. Journal of Geographical Sciences, 2015, 25(1): 85–100
Shan XJ, Jin XS, Dai FQ,et al. Population dynamics of fish species in a marine ecosystem: A case study in the Bohai Sea,China. Marine and Coastal Fisheries, 2016, 8(1): 100–117
Shan XJ, Sun P, Jin XS,et al. Long-term changes in fish assemblage structure in the Yellow River Estuary ecosystem,China. Marine and Coastal Fisheries, 2013, 5(1): 65–78
Tang QS, Jin XS, Wang J,et al. Decadal-scale variations of ecosystem productivity and control mechanisms in the Bohai Sea. Fisheries Oceanography, 2003, 12(4–5): 223–233
Tang QS, Guo XW, Sun Y,et al. Ecological conversion efficiency and its influencers in twelve species of fish in the Yellow Sea ecosystem. Journal of Marine Systems, 2007,67(3–4): 282−291
Tang QS, Su JL, Sun S,et al. A study of marine ecosystem dynamics in the coastal ocean of China. Advanced in Earth Sciences, 2005, 20(12): 1288−1299 [唐启升, 苏纪兰, 孙松, 等. 中国近海生态系统动力学研究进展. 地球科学进展, 2005, 20(12): 1288−1299]
Tang QS, Fang JG, Zhang JH,et al. Impacts of multiple stressors on coastal ocean ecosystems and Integrated Multi-trophic aquaculture. Progress in Fishery Sciences, 2013, 34(1): 1–11[唐启升, 方建光, 张继红, 等. 多重压力胁迫下近海生态系统与多营养层次综合养殖. 渔业科学进展, 2013, 34(1):1–11]
Tian MC, Sun BL, Yang JM. Analysis of the fish fauna of the Bohai Sea. Studia Marina Sinica, 1993(1): 157–167 [田明诚,孙宝龄, 杨纪明. 渤海鱼类区系分析. 海洋科学集刊,1993(1): 157–167]
Wan RJ, Jiang YW. The distribution and variation of eggs and larvae of osteichthyes in the Bohai Sea. Journal of Fishery Sciences of China, 1998, 5(1): 43–50 [万瑞景, 姜言伟. 渤海硬骨鱼类鱼卵和仔稚鱼分布及其动态变化. 中国水产科学, 1998, 5(1): 43–50]
Wan RJ, Bian XD. Size variability and natural mortality dynamics of anchovyEngraulis japonicuseggs under high fishing pressure. Marine Ecology Progress Series, 2012(465):243–251
Wan RJ, Wei H, Sun S,et al. Spawning ecology of the anchovyEngraulis japonicusin the spawning ground of the Southern Shandong PeninsulaⅠ. Abundance and distribution characters of anchovy eggs and larvae. Acta Zoologica Sinica, 2008,54(5): 785–797 [万瑞景, 魏皓, 孙珊, 等. 山东半岛南部产卵场鳀鱼的产卵生态Ⅰ. 鳀鱼鱼卵和仔稚幼鱼的数量与分布特征. 动物学报, 2008, 54(5): 785–797]
Wan RJ, Ni XB, Bian XD,et al. Species composition and abundance distribution pattern of ichthyoplankton and their relationship with environmental factors in the East China Sea ecosystem. Journal of Fisheries of China, 2014, 38 (9):1375–1398 [万瑞景, 曾定勇, 卞晓东, 等. 东海生态系统中鱼卵、仔稚鱼种类组成、数量分布及其与环境因素的关系. 水产学报, 2014, 38(9): 1375−1398]
Wei S, Jiang WM. Study on food web of fishes in the Yellow Sea.Oceanologia et Limnologia Sinica, 1992, 23(2): 182–192[韦晟, 姜卫民. 黄海鱼类食物网的研究. 海洋与湖沼,1992, 23(2): 182–192]
Wei H, Su J, Wan RJ,et al. Tidal front and the convergence of anchovy (Engraulis japonicus) eggs in the Yellow Sea.Fisheries Oceanography, 2003, 12(4–5): 434–442
Wu R, Li C, Lin J. Enhanced winter warming in the Eastern China coastal waters and its relationship with ENSO.Atmospheric Science Letters, 2017, 18(1): 11–18
Whittaker RH. Evolution and measurement of species diversity.Taxon,1972, 21(2-3): 213–251
Zhao X, Hamre J, Li F,et al. Recruitment, sustainable yield and
possible ecological consequences of the sharp decline of the anchovy (Engraulis japonicus) stock in the Yellow Sea in the 1990s. Fisheries Oceanography, 2003, 12(4–5): 495–501
Zhou L. Ecological responses of marine phytoplankton to the oil spills. Master′s Thesis of Graduate University of Chinese Academy of Sciences, 2014 [周利. 海洋浮游植物对溢油的生态响应研究. 中科院研究生院硕士研究生学位论文,2014]
Zhu XH, Tang QS. Structuring dominant components within fish community in Bohai Sea system. Studia Marina Sinica,2002(44): 159–168 [朱鑫华, 唐启升. 渤海鱼类群落优势种结构及其种间更替. 海洋科学集刊, 2002(44): 159–168]
Zhu XH, Wu HZ, Xu FS,et al. The heterogeneity of spatiotemporal patterns of nekton community structure in the coastal waters of Yellow Sea and Bohai Sea. Acta Zoologica Sinica, 1994, 40(3): 241−252 [朱鑫华, 吴鹤洲,徐凤山, 等. 黄、渤海沿岸水域游泳动物群落结构时空格局异质性研究. 动物学报, 1994, 40(3): 241−252]
