苗期外源脱落酸和茉莉酸缓减小麦花后干旱胁迫的效应及其生理机制
2018-03-28谢静静戴廷波
谢静静,王 笑,蔡 剑,周 琴,戴廷波,姜 东
(南京农业大学农学院/农业部作物生理生态与生产管理重点实验室/江苏省现代作物生产协同创新中心,江苏南京 210095)
花后干旱是限制小麦产量稳定的最主要逆境因子之一[1-2]。干旱影响小麦叶片叶绿素荧光特性,降低叶片光合能力与抗氧化酶活性,导致活性氧积累,造成氧化胁迫,加重膜脂损伤,抑制了小麦的正常生长发育,从而降低小麦的产量和品质[2-3]。脱落酸(abscisic,ABA)作为“胁迫激素”在植物耐旱性形成中发挥着重要的作用[4]。ABA能够提高干旱逆境下叶片的叶绿素含量和净光合速率,激活抗氧化系统相关酶活性,清除活性氧,进而减轻干旱对作物的伤害[5]。茉莉酸类物质(jasmonates,JAs)在一些逆境响应中具有与脱落酸相似的作用,主要代表物质是茉莉酸(jasmonic acid,JA)和茉莉酸甲脂(methyle Jasmonate,MeJA)。研究发现,外源JA可显著提高干旱胁迫下芸薹属作物的鲜重、叶绿素含量和叶片含水量,增强植株的氧化应激[6]。MeJA在干旱胁迫诱导的气孔运动中发挥重要作用[7]。不同时期喷施ABA和JA对作物生长的调控机制存在显著差异。苗期施用ABA可增强小麦叶片抗氧化能力,降低干旱胁迫导致的活性氧积累及膜脂过氧化伤害,从而增强植株耐旱性[8]。小麦花后施用ABA通过诱导叶片内源生长素和赤霉素的积累,增强叶片光合能力,延缓旗叶衰老,最终提高籽粒产量[9]。花后喷施MeJA可显著提高干旱胁迫下大豆植株叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)等抗氧化酶的活性,增加干旱胁迫下产量和收获指数[10]。苗期施用JA可通过诱导信号物质一氧化氮(NO)合成,调控抗坏血酸-谷胱甘肽循环,增强植株抗氧化能力,有助于提高小麦幼苗耐旱性[11]。然而关于小麦生长发育早期喷施外源ABA和JA是否在提高植株对花后干旱胁迫的耐性方面存在长期效应还尚未见报道。本研究在小麦生育前期叶面喷施ABA、JA及其对应的抑制剂,并在花后施加干旱胁迫处理,以期明确外源ABA和JA对小麦花后干旱胁迫的减缓作用及其生理机制,为小麦抗旱栽培提供技术与理论支撑。
1 材料与方法
1.1 试验设计
供试品种为本区域大面积推广的小麦品种扬麦16。采用盆栽试验,土壤自然风干后过筛与沙子2∶1混合,与肥料拌匀后称取7.5 kg装入塑料桶中,等量水沉实后播种。每桶施用尿素1.5 g和磷酸二氢钾1 g,拔节期追肥,氮肥基追比为3∶2。每桶选取饱满均匀一致的种子播种21粒,三叶一心期间苗,每桶留苗7株。
分别在四叶一心期和六叶一心期进行外源ABA和JA的叶面喷施,共设6个处理:(1)正常水分处理(CK,土壤含水量相当于田间持水量70%~80%);(2)花后干旱胁迫(D,土壤含水量相当于田间持水量30%~40%);(3)0.01 mmol·L-1ABA+花后干旱胁迫(ABA+D);(4)0.1 mmol·L-1JA+花后干旱胁迫(JA+D);(5)1 μmol·L-1氟啶酮(FLU,ABA合成抑制剂)+花后干旱胁迫(FLU+D);(6)0.2 mmol·L-1去甲二氢愈创木酸(NDGA,JA合成抑制剂)+花后干旱胁迫(NDGA+D),喷施溶液含有0.1%的乙醇(能够溶解ABA的最少含量)和0.002%的吐温20%,每天清晨和傍晚进行喷施,连续喷施两天,CK和D处理均对应喷等量的乙醇和吐温溶液。前期正常水分处理,在小麦花后3 d开始控水,到花后7 d土壤含水量维持在田间持水量的30%~40%,土壤水势在-1.8 MPa左右,干旱胁迫处理5 d后复水,植株继续生长至成熟。试验为随机区组设计,重复3次。干旱胁迫5 d后取植株旗叶进行相关指标测定,每个处理取24片旗叶,鲜样立即放入液氮中速冻,后转入-80 ℃冰箱中保存。成熟期每个处理取3盆进行产量及其构成因素的测定。
1.2 测定项目与方法
1.2.1 旗叶水分含量和水势的测定
用烘干法测定叶片相对含水量(LRWC)。叶片相对含水量=(叶片鲜重-叶片干重)/(叶片饱和重-叶片干重)×100%。采用HR33T露点微伏计(Wescor Inc., Logan, UT, USA)测定叶片凌晨叶水势,每处理测定三个重复。
1.2.2 旗叶叶绿素含量与光合和荧光参数的测定
采用SPAD-502叶绿素仪(Minolta Co, Japan)测定叶绿素含量。采用LI-6400(LI-Cor Inc, USA)便携式光合作用测定系统测定旗叶净光合速率,时间为9:00-11:00,采用开放式气路,CO2浓度为380 μmol·L-1,红蓝光源,光合有效辐射为1 200 μm·m-2·s-1。叶绿素荧光参数使用叶绿素荧光成像系统CF Imager(Technologica, UK)进行测定,先将幼苗暗适应30 min,进行暗适应测定,然后再进行光适应测定。
1.2.3 旗叶渗透物质与激素含量的测定
可溶性糖含量测定采用蒽酮硫酸比色法,游离氨基酸含量测定采用茚三酮比色法,脯氨酸含量测定采用磺基水杨酸法,以上均参考慈敦伟等[12]的方法。酶联免疫法(ELISA)测定ABA和JA含量,试剂盒分别来自中国农业大学和北京冬歌生物科技有限公司。
1.2.4 旗叶活性氧含量及抗氧化酶活性的测定
过氧化氢含量采用H2O2测定试剂盒,试剂盒来自南京建成生物工程研究所;超氧阴离子自由基(O2-·)和抗氧化酶活性等的测定参照郑春芳等[13]的方法,O2-·产生速率测定采用羟胺法;SOD活性测定采用氮蓝四唑(NBT)光还原法;过氧化氢酶(CAT)测定采用过氧化氢法;抗坏血酸过氧化物酶(APX)活性测定采用抗坏血酸法;谷胱甘肽还原酶(GR)活性测定采用紫外比色法;丙二醛(MDA)含量测定采用TCA-TBA法。
1.3 数据分析
数据采用SPSS 20.0软件进行分析和显著性差异检验,使用Microsoft Excel 2010作图。
2 结果与分析
2.1 外源ABA和JA对花后干旱胁迫下小麦籽粒产量的影响
与对照(CK)相比,花后干旱胁迫(D)显著降低小麦穗粒数、千粒重和籽粒产量(表1),减产幅度为24.6%,对穗数影响不显著。与D处理相比,喷施ABA和JA处理显著提高了小麦的穗粒数和千粒重,虽然增加了产量,但影响不显著。喷施ABA和JA抑制剂处理的产量及其构成因素与D处理差异不明显。这表明外源ABA和JA能在一程度上有效缓解花后干旱胁迫对小麦产量形成的影响。
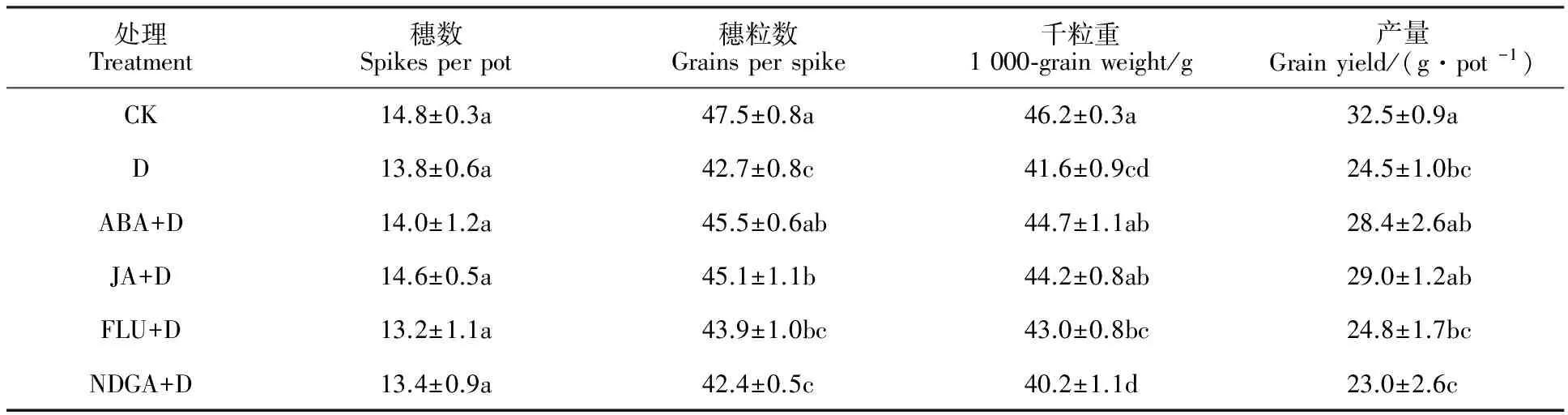
表1 外源ABA和JA对花后干旱胁迫下小麦产量及其构成因素的影响Table 1 Effects of exogenous ABA and JA on grain yield and yield components in wheat under post-anthesis drought stress
同一列数值后的不同小写字母表示处理间在0.05水平上差异显著。
The lowercase letters in same column indicate significant differences among the treatments at 0.05 level.
2.2 外源ABA和JA对花后干旱胁迫下小麦旗叶激素含量的影响
花后干旱胁迫下小麦旗叶内源ABA含量显著提高,内源JA含量也略有上升,但与对照差异不显著(图1)。与D处理相比,叶面喷施JA显著提高了小麦旗叶内源ABA和JA的含量;喷施ABA后内源ABA含量显著升高,内源JA含量虽有增加,但变化不显著。喷施抑制剂FLU和NDGA对内源ABA和JA影响不显著。这表明外源ABA和JA可促进内源ABA和JA的积累,激活耐旱相关生理响应。
2.3 外源ABA和JA对花后干旱胁迫下小麦旗叶水分含量和水势的影响
花后干旱胁迫显著降低小麦旗叶相对含水量和叶片水势(图2)。干旱胁迫下,叶面喷施ABA显著提高旗叶相对含水量和水势,与D处理相比的增幅分别为18.9%和9.9%;喷施JA处理的叶片相对含水量略有升高,显著增加叶片水势;FLU+D和NDGA+D处理的叶片含水量和水势与D处理没有显著差异。这说明外源ABA和JA有利于保持干旱胁迫下小麦叶片水分状态,提高耐旱性。

图1 外源ABA和JA对花后干旱胁迫下小麦旗叶内源ABA和JA含量的影响

图2 外源ABA和JA对花后干旱胁迫下小麦旗叶相对含水量和水势的影响
2.4 外源ABA和JA对花后干旱胁迫下小麦旗叶净光合速率、SPAD值和荧光参数的影响
花后干旱胁迫显著降低小麦旗叶净光合速率(Pn)、SPAD值、最大光化学效率(Fv/Fm)和实际光化学效率(ФPSⅡ)(图3)。与D处理相比,喷施ABA和JA后旗叶Pn、SPAD值、Fv/Fm和ФPSⅡ明显增加。FLU+D和NDGA+D处理的旗叶Pn、SPAD值、Fv/Fm和ФPSⅡ与D处理差异不明显。可见,外源ABA和JA有效缓解了干旱胁迫对叶绿素的破坏,降低了干旱胁迫对光系统Ⅱ的损伤,提高了旗叶的光合能力。
2.5 外源ABA和JA对花后干旱胁迫下小麦旗叶抗氧化系统的影响
花后干旱胁迫显著提高了小麦旗叶O2-·、H2O2含量、MDA含量和相对电导率(图4),明显增加旗叶SOD、CAT、APX和GR活性(图5)。干旱胁迫下,喷施ABA和JA后旗叶O2-·、H2O2和MDA含量及相对电导率显著降低,4种抗氧化酶活性均不同程度提高。FLU+D与ABA+D处理的旗叶O2-·、H2O2和MDA含量及相对电导率均不同程度增加,对4种抗氧化酶活性影响较小。这表明外源ABA和JA能抑制干旱胁迫下小麦旗叶活性氧积累,提高抗氧化能力,降低膜脂过氧化程度,减轻干旱胁迫对小麦的不利影响。

图3 外源ABA和JA对花后干旱胁迫下小麦旗叶净光合速率(Pn)、SPAD值和叶绿素荧光参数的影响

图4 外源ABA和JA对花后干旱胁迫下小麦旗叶活性氧积累、相对电导率和MDA含量的影响
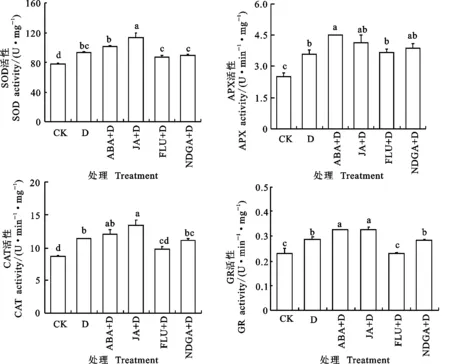
图5 外源ABA和JA对花后干旱胁迫下小麦旗叶抗氧化酶活性的影响
2.6 外源ABA和JA对花后干旱胁迫下小麦旗叶渗透物质含量的影响
花后干旱胁迫显著提高了小麦旗叶的可溶性糖、游离氨基酸和脯氨酸含量(图6)。与D处理相比,ABA+D处理的旗叶可溶性糖、游离氨基酸和脯氨酸的含量分别增加了22.3%、38.3%和56.6%,JA+D处理分别增加了15.6%、42.8%和62.8%,喷施抑制剂FLU处理对旗叶可溶性糖和游离氨基酸含量影响不大,抑制剂NDGA处理显著降低游离氨基酸和脯氨酸的含量。说明外源ABA和JA进一步促进了干旱胁迫下旗叶渗透调节物质的积累,有利于植株渗透调节能力。
3 讨 论
3.1 外源ABA和JA对花后干旱下小麦旗叶内源激素及籽粒产量的影响
植物激素对植物体的生命活动起着重要的调节作用,干旱引起的ABA积累可诱导气孔关闭,提高细胞的导水能力,有利于维持细胞膨压,进而促进植物的生长发育[14];JA的积累能够诱导一系列植物响应干旱的形态和生理反应,从而提高植物对干旱胁迫的耐性[15]。研究发现,外源ABA可以诱导干旱胁迫下玉米内源ABA的合成,提高叶片净光合速率,维持叶片的水分含量[16];MeJA处理后,干旱胁迫下花生幼苗内源ABA含量升高,耐旱性增强[17]。本试验中,喷施ABA和JA的小麦旗叶在干旱胁迫下具有更高的ABA和JA含量,表明外源ABA和JA促进了花后干旱胁迫下小麦内源逆境激素ABA和JA的积累,进而调控干旱响应的相关生理过程。小麦遭遇花后干旱胁迫,籽粒产量会受到不同程度的影响,主要归因于小麦千粒重的变化[18-19]。本研究结果表明,花后干旱胁迫对穗数的影响不大,但显著降低了穗粒数和千粒重。干旱胁迫下,外源ABA和JA显著增加了小麦的穗粒数和千粒重,提高了最终的籽粒产量,而施用抑制剂处理表现出相反的趋势,表明外源ABA和JA增强了小麦应对花后干旱胁迫的耐性。

图6 外源ABA和JA对花后干旱胁迫下小麦旗叶可溶性糖、游离氨基酸和脯氨酸含量的影响
3.2 外源ABA和JA对花后干旱下小麦旗叶光合能力和抗氧化系统的影响
干旱胁迫会引起叶绿素的降解,抑制叶片净光合速率[20],从而影响作物的干物质积累及籽粒产量形成[21-22]。有研究表明,外源ABA可显著提高干旱胁迫下草莓叶片的净光合速率,有效缓解干旱胁迫对草莓光合的抑制作用[23];外源JA能够明显增加盐胁迫下荞麦叶片的净光合速率,提高植株的耐盐性[24]。本研究中,花后干旱胁迫下经外源ABA和JA处理的小麦旗叶SPAD值更高,光合能力更强,表现出更高的耐旱性。叶绿素荧光参数能够有效反映植物对干旱胁迫的耐性[25]。本研究中,喷施ABA和JA显著增加了干旱胁迫下小麦旗叶的最大光化学效率和实际光化学效率,这与前人[26-27]的研究结果一致,表明外源ABA和JA有效降低了干旱对小麦叶片光系统Ⅱ的损伤,有利于保证光合电子传递和光合磷酸化的正常进行,提高了水分亏缺条件下植物对光能的吸收和利用。活性氧(ROS)在细胞内的积累会导致膜脂过氧化,损害细胞膜系统,从而对植物造成伤害。在非严重胁迫情况下,抗氧化系统相关酶能够有效清除体内产生的ROS,但当遭遇重度干旱胁迫时,抗氧化酶无法及时清除体内过量的活性氧,从而引起氧化胁迫损伤[28]。Souza等[29]研究发现,外源ABA可以显著增加干旱胁迫下玉米叶片的SOD、CAT、APX、POD等抗氧化酶的活性,同时降低MDA的含量。Qiu等[30]指出,外源JA处理显著增加了花生幼苗的SOD和CAT活性。本研究发现,花后干旱胁迫诱导SOD、CAT、APX和GR活性升高,但旗叶仍具有较高的ROS含量,表明干旱诱导的抗氧化酶并不能完全清除过量的活性氧。外源ABA和JA处理的小麦旗叶O2-·和H2O2含量显著降低,SOD、CAT、APX、GR等抗氧化酶的活性较D处理相比进一步升高,有效清除了干旱胁迫产生的ROS,细胞膜脂过氧化程度降低,细胞内电解质外渗减少,从而缓解了干旱胁迫对细胞的损伤,提高了植株的耐旱性。
3.3 外源ABA和JA对花后干旱下小麦旗叶渗透调节物质的影响
干旱胁迫下,植株通过调控渗透调节物质的合成和积累降低植物细胞渗透势,从而维持细胞膨压,提高作物的耐旱性。可溶性糖、脯氨酸等是响应干旱胁迫较为重要的渗透调节物质[31],在稳定生物膜结构、保持细胞水分、提高作物抗旱性上发挥重要的作用。阮英慧等[5]研究发现,外源ABA能够提高干旱胁迫下大豆叶片可溶性糖和游离氨基酸含量,较高浓度的ABA可以诱导脯氨酸的积累,提高大豆叶片的耐旱性。Wu等[32]认为,MeJA预处理的花椰菜在干旱胁迫下具有较高的可溶性糖和脯氨酸含量,有利于缓解了干旱胁迫的不利影响。本试验中,外源ABA和JA促进了干旱胁迫下旗叶可溶性糖、游离氨基酸和脯氨酸等渗透物质的积累,提高了细胞质溶质浓度,缓解了干旱引起的细胞水分散失,减少了干旱胁迫的伤害。
综上所述,外源ABA和JA诱导干旱胁迫下小麦旗叶内源激素ABA和JA的含量升高,激活抗氧化酶系统相关酶活性,有效提高了活性氧的清除能力,减少膜脂过氧化程度,缓解叶绿素的降解,避免干旱引起的氧化胁迫对光合系统的破坏。此外,外源ABA和JA提高了小麦旗叶渗透调节能力,降低干旱导致的水分散失。外源ABA和JA通过以上调控途径增强了花后干旱胁迫下小麦的光合能力,促进了干物质的积累,最终提高了小麦的籽粒产量,有利于小麦耐旱性的增强。
[1] JI-BANG H U,WANG C Y,GUO T C,etal.Effects of high temperature and drought stress on grain filling characteristics in wheat during grain filling period [J].JournalofHenanAgriculturalUniversity,2008,42(6):597.
[2] FAROOQ M,HUSSAIN M,SIDDIQUE K H M.Drought stress in wheat during flowering and grain-filling periods [J].CriticalReviewsinPlantSciences,2014,33(4):343.
[3] SAIRAM R K,DESHMUKH P S,SHUKLA D S.Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat [J].JournalofAgronomyandCropScience,2010,178(3):171.
[4] NAKASHIMA K,YAMAGUCHI-SHINOZAKI K.ABA signaling in stress-response and seed development [J].PlantCellReports,2013,32(7):959.
[5] 阮英慧,董守坤,刘丽君,等.干旱胁迫下外源脱落酸对大豆花期生理特性的影响 [J].大豆科学,2012,31(3):385.
RUAN Y H,DONG S K,LIU L J,etal.Effects of exogenous abscisic acid on physiological characteristics in soybean flowering under drought stress [J].SoybeanScience,2012,31(3):385.
[6] ALAM M M,NAHAR K,HASANUZZAMAN M,etal.Exogenous jasmonic acid modulates the physiology,antioxidant defense and glyoxalase systems in imparting drought stress tolerance in differentBrassicaspecies [J].PlantBiotechnologyReports,2014,8(3):279.
[7] MUNEMASA S,MORI I C,MURATA Y.Methyl jasmonate signaling and signal crosstalk between methyl jasmonate and abscisic acid in guard cells [J].PlantSignalingandBehavior,2011,6(7):939.
[8] WEI L,WANG L,YANG Y,etal.Abscisic acid enhances tolerance of wheat seedlings to drought and regulates transcript levels of genes encoding ascorbate-glutathione biosynthesis [J].FrontiersinPlantScience,2015,6:458.
[9] 杨东清,王振林,尹燕枰,等.外源ABA和6-BA对不同持绿型小麦旗叶衰老的影响及其生理机制 [J].作物学报,2013,39(6):1096.
YANG D Q,WANG Z L,YIN Y P,etal.Effects of exogenous ABA and 6-BA on flag leaf senescence in different types of stay-green wheat and relevant physiological mechanisms [J].ActaAgronomicaSinica,2013,39(6):1096.
[10] ANJUM S A,WANG L,FAROOQ M,etal.Methyl jasmonate-induced alteration in lipid peroxidation,antioxidative defence system and yield in soybean under drought [J].JournalofAgronomyandCropScience,2011,197(4):296.
[11] SHAN C,ZHOU Y,LIU M.Nitric oxide participates in the regulation of the ascorbate-glutathione cycle by exogenous jasmonic acid in the leaves of wheat seedlings under drought stress [J].Protoplasma,2015,252(5):1397.
[12] 慈敦伟,姜 东,戴廷波,等.镉胁迫对小麦幼苗膜脂过氧化作用及镉吸收转运的影响 [J].麦类作物学报,2009,29(3):509.
CI D W,JIANG D,DAI T B,etal.Effect of Cd stress on lipid peroxidation and Cd uptake and translocation in wheat seedling [J].JournalofTriticeaeCrops,2009,29(3):509.
[13] 郑春芳,姜 东,戴廷波,等.外源一氧化氮供体硝普钠浸种对盐胁迫下小麦幼苗碳氮代谢及抗氧化系统的影响 [J].生态学报,2010,30(5):1175.
ZHENG C F,JIANG D,DAI T B,etal.Effects nitroprusside,anitric oxide donor,on carbon and nitrogen metabolism and the activity of the antioxidation system in wheat seedings under salt stress [J].ActaEcologicaSinica,2010,30(5):1175.
[14] SAVCHENKO T,KOLLA V A,WANG C Q,etal.Functional convergence of oxylipin and abscisic acid pathways controls stomatal closure in response to drought [J].PlantPhysiology,2014,164(3):115.
[15] OLLAS C D,DODD I C.Physiological impacts of ABA-JA interactions under water-limitation [J].PlantMolecularBiology,2016,91(6):641.
[16] SOUZA T C D,MAGALHAES P C,CASTRO E M D,etal.The influence of ABA on water relation,photosynthesis parameters,and chlorophyll fluorescence under drought conditions in two maize hybrids with contrasting drought resistance [J].ActaPhysiologiaePlantarum,2013,35(2):515.
[17] 潘瑞炽,古焕庆.茉莉酸甲酯对花生幼苗生长和抗旱性的影响 [J].植物生理学报,1995,31(3):215.
PAN R C,GU H Q.Effect of methyl jasmonate on the growth and drought resistance in peanut seedlings [J].PlantPhysiologyJournal,1995,31(3):215.
[18] WANG X,VIGNJEVIC M,LIU F,etal.Drought priming at vegetative growth stages improves tolerance to drought and heat stresses occurring during grain filling in spring wheat [J].PlantGrowthRegulation,2015,75(3):677.
[19] 史国安,郭香凤,刘素云,等.灌浆期干旱对小麦旗叶衰老和籽粒产量的影响 [J].西北农业学报,1999,8(2):26.
SHI G A,GUO X F,LIU S Y,etal.Effect of water stress on flag leaves senescence and grain yield of winter wheat in grain filling stage [J].ActaAgriculturaeBoreali-OccidentalisSinica,1999,8(2):26.
[20] 吴 顺,张雪芹,蔡 燕.干旱胁迫对黄瓜幼苗叶绿素含量和光合特性的影响 [J].中国农学通报,2014,30(1):133.
WU S,ZHANG X Q,CAI Y.Effects of drought stress chlorophyll contents and photosynthetic characteristics of cucumber seedlings [J].ChineseAgriculturalScienceBulletin,2014,30(1):133.
[21] SHANKER A K,MAHESWARI M,YADAV S K,etal.Drought stress responses in crops [J].FunctionalandIntegrativeGenomics,2014,14(1):11.
[22] 谷艳芳,丁圣彦,高志英,等.干旱胁迫下冬小麦光合产物分配格局及其与产量的关系[J].生态学报,2010,30(5):1167.
GU Y F,DING S Y,GAO Z Y,etal.The pattern of photosynthate partitioning in drought-stressed winter wheat and its relationship with yeild [J].ActaEcologicaSinica,2010,30(5):1167.
[23] 王书宏,杜永吉.外源激素对干旱胁迫下草莓光合特性的影响 [J].中国农学通报,2008,24(12):367.
WANG S H,DU Y J.Effects of exogenous hormones on photosynthetic parametersof strawberry under drought stress [J].ChineseAgriculturalScienceBulletin,2008,24(12):367.
[24] 杨洪兵,孙 萍.外源水杨酸和茉莉酸对荞麦幼苗耐盐生理特性的效应 [J].植物生理学报,2012,48(8):767.
YANG H B,SUN P.Effects of exogenous salicylic acid and jasmonic acid on physiological traits of salt tolerance in buckwheat(FagopyrumesculentumMoench) seedlings [J].PlantPhysiologyJournal,2012,48(8):767.
[25] 张仁和,马国胜,柴 海,等.干旱胁迫对玉米苗期叶绿素荧光参数的影响 [J].干旱地区农业研究,2010,28(6):170.
ZHANG R H,MA G S,CHAI H,etal.Effect of drought stress on chlorophyll fluorescence of maize leaves at seedling [J].AgriculturalResearchintheAridAreas,2010,28(6):170.
[26] KAUR M,GUPTA A K,ZHAWAR V K.Antioxidant response and Lea genes expression under exogenous ABA and water deficit stress in wheat cultivars contrasting in drought tolerance [J].JournalofPlantBiochemistryandBiotechnology,2012,23(1):18.
[27] 董桃杏,蔡昆争,曾任森.茉莉酸甲酯(MeJA)对干旱胁迫下水稻幼苗光合作用特性的影响 [J].生态环境学报,2009,18(5):1872.
DONG T X,CAI K Z,ZENG R S.Effects of methyl jasmonate(MeJA) on photosynthetic traits of rice seedlings under drought stress [J].EcologyandEnvironmentalSciences,2009,18(5):1872.
[28] 叶 君,邓西平,王仕稳,等.干旱胁迫下褪黑素对小麦幼苗生长、光合和抗氧化特性的影响 [J].麦类作物学报,2015,35(9):1275.
YE J,DENG X P,WANG S W,etal.Effects of melatonin on growth,photosynthetic characteristics and antioxidant system in seedling of wheat under drought stress [J].JournalofTriticeaeCrops,2015,35(9):1275.
[29] SOUZA T C D,MAGALHAES P C,CASTRO E M D,etal.ABA application to maize hybrids contrasting for drought tolerance:changes in water parameters and in antioxidant enzyme activity [J].PlantGrowthRegulation,2014,73(3):205.
[30] QIU Z,GUO J,ZHU A,etal.Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress [J].EcotoxicologyandEnvironmentalSafety,2014,104(1):202.
[31] 杜伟莉,高 杰,胡富亮,等.玉米叶片光合作用和渗透调节对干旱胁迫的响应 [J].作物学报,2013,39(3):530.
DU W L,GAO J,HU F L,etal.Responses of drought stress on photosynthetic trait and osmotic adjustment in two maize cultivars [J].ActaAgronomicaSinica,2013,39(3):530.
[32] WU H,WU X,LI Z,etal.Physiological evaluation of drought stress tolerance and recovery in cauliflower(Brassicaoleracea L.) seedlings treated with methyl jasmonate and coronatine [J].JournalofPlantGrowthRegulation,2012,31(1):113.
