野榆钱菠菜三种异型种子幼苗的生长特性
2018-03-26马赫魏岩穆晨
马赫,魏岩,穆晨
(1.新疆维吾尔自治区草原总站,新疆 乌鲁木齐 830049;2.新疆农业大学草业与环境科学学院,新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
荒漠是一种严酷多变的自然环境,降水少,蒸发量大,昼夜温差大,在时间和空间上都具有高度异质性[1],这种极端的自然环境严重抑制了植物的生长和生存,但在长期的自然选择与进化中,以荒漠、半荒漠植物为主的具有旱生、盐生结构特征的植被适应了这种极端环境[2],逐渐发展成此环境的建群种和共建种[3]。这些荒漠、半荒漠植物为确保在荒漠环境中生存和发展,在长期的进化历程中演化出了与这种严酷生境相适应的特殊适应机制,种子异型性现象便是荒漠植物适应严酷环境的一种特殊机制[4-5]。
种子异型性(seed heteromorphism)是指在同一母体植株上产生具有不同形状或行为的种子的现象[5-7]。种子异型性现象被认为是植物为适应恶劣异质环境,在时间和空间上为避免密集负效应及减弱同胞子代间的竞争等而进化出的一种“两头下注”的适应策略[5,8-9],“两头下注”策略是荒漠植物为适应长期的逆境胁迫使种子产生适度变异从而提高其对异质环境的适应能力以及种群的繁殖成功率[6]的一种生存策略。具种子异型性的植物主要集中在菊科(Asteraceae)、藜科(Chenopodiaceae)、禾本科(Poaceae)、十字花科(Brassicaceae)、豆科(Leguminosae)等几个有限的科中[7]。异型种子的差异主要体现在颜色、大小以及存有的附属物上[10],这些差异也体现在异型种子的萌芽期[10]、生存和生殖时间[11]、种子的散布期[12]、种子库动态期[10]上。
藜科植物在新疆荒漠地区形成多种群落,是重要的建群种和共建种,在荒漠植被景观、荒漠草地利用上都有着很重要的作用[13],野榆钱菠菜(Atriplexaucheri)是一种典型的具种子异型性的泌盐植物,主要分布于中国新疆的荒漠、半荒漠、盐碱地上[3],在改良荒漠盐碱地方面有着巨大的开发潜力[14-15]。野榆钱菠菜具有3种异型种子:无苞片包被的黑色种子(单粒重1.2 mg),小型苞片包被的黑色种子(1.7 mg),大型苞片包被的褐色种子(3.6 mg)[16]。目前,国内外对植物种子异型性现象的研究主要集中在形态观察、种子萌发行为、植物耐盐性、散布方式、子代种子耐盐性上[17-20],关于异型种子在萌发物候及萌生幼苗是否存在差异性方面尚缺少系统研究。本研究以种子异型性植物野榆钱菠菜为研究对象,探究异型种子在萌发物候上的差异以及异型种子萌生幼苗的生长特性,旨在揭示种子异型性植物如何利用“两头下注”策略适应荒漠环境,为阐明种子异型性植物的生态适应机制奠定了理论基础,为开发这种优良荒漠牧草、改善新疆荒漠环境提供科学根据。
1 材料与方法
1.1 研究材料
2014年10月底在新疆准噶尔盆地南缘(43°45′44″-43°47′25″ N, 87°33′16″-87°34′56″ E)乌鲁木齐荒漠的自然种群随机选取50株植株,在实验室自然晾干,将3种异型种子分离,选取成熟饱满的种子放置在纸袋中作好标记,在室温条件下于阴暗处储藏。
1.2 实验方法
1.2.1实验设计与材料的种植 实验地位于新疆农业大学校园内(43°48′46″ N, 87°33′54″ E),共3个小区,每个小区随机种植一种异型种子,设置10个重复,每个重复长30 cm 、宽30 cm,每个重复内均匀播撒400粒种子,此密度接近藜科植物在野外条件下的密度[20]。于2014年11月中旬下雪前播种,播种完毕后,用厚约1 cm的细土均匀覆盖。
1.2.2异型种子萌发物候、出苗率、子叶存留时间的观察 自2015年3月1日起每天关注实验地内的种子萌发出土情况,观察记录3种异型种子的出苗率、子叶出土时间以及天气情况。
野榆钱菠菜为双子叶植物,幼苗属子叶出土幼苗,实验中其萌发物候及出苗率的测定以子叶出土为准。子叶出土后,记录每个重复内的出苗率,若出苗多则采用四分法计数,求平均值;每隔10 d测量子叶的长度、宽度;种子萌发出土后,每种异型种子随机选取10株幼苗做好标记,记录幼苗的真叶出现时间及子叶脱落时间。
1.2.3幼苗形态、生长特性的观察与记录 每隔10 d取样一次,每次随机挑选5株幼苗,用取样器将幼苗完整取出,取样时保证根等幼苗构件的完整性,带回实验室测量,用剪刀将子叶、胚轴、根剪下,测量根的长度,在万分之一电子秤上测量根、子叶、胚轴的鲜重,然后将幼苗的构件按地上部分及地下部分装入纸袋做好标记,于阴暗处自然晾干,在万分之一天平上测量其干重。至2015年5月11日,3种异型种子幼苗的子叶均已脱落,此时停止取样。
1.3 统计分析
生物量及根冠比均以干重为准,地上生物量为幼苗地上部分构件的总质量(干重),地下生物量为幼苗地下部分构件的总质量(干重),根冠比为地下生物量与地上生物量的比值(干重);用SPSS 20.0对出苗率、子叶长宽、幼苗根长、根冠比采用单因素方差分析法进行差异显著性检验;用Sigmaplot 12.5对所得数据进行制图。
2 结果与分析
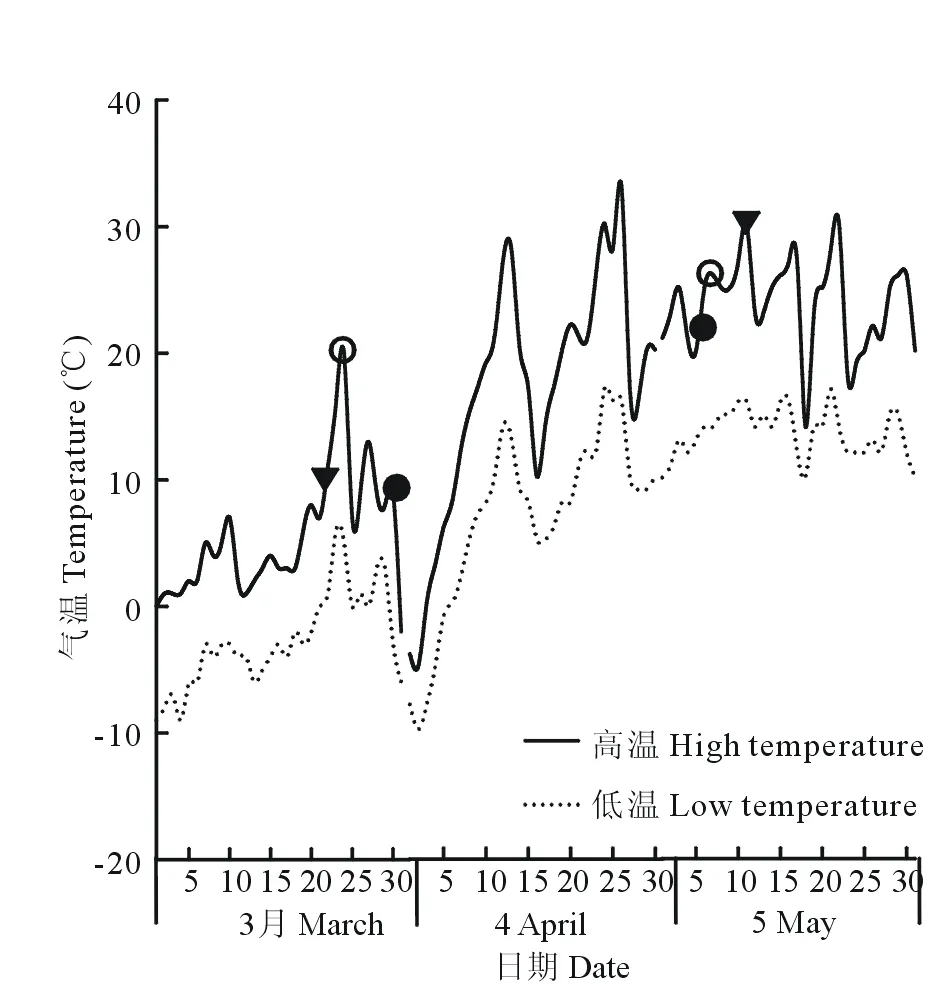
图1 实验区3、4、5月气温Fig.1 Experimentation area temperature in March, April and May 图中符号(●)、(○)、(▼)分别表示黑色小种子、黑色大种子、褐色种子的出苗时间及子叶脱落时间。The symbols (●), (○), (▼) indicate the emergence time and cotyledon abscission time of the small black seeds, big black seeds and brown seeds.
2.1 异型种子后代幼苗的萌发物候、出苗率、真叶出现时间、子叶存留时间
由图1可知,在相同温度、水分、土壤条件下,野榆钱菠菜3种异型种子在3月底均已破土出苗,褐色种子最先破土出苗,但子叶脱落时间最晚,说明褐色种子幼苗的子叶存留时间最长。黑色大种子出苗时间与幼苗子叶脱落时间介于褐色种子与黑色小种子之间,其子叶存留时间较褐色种子短,较黑色小种子长,黑色小种子破土出苗时间最晚,但子叶脱落时间最早,其子叶存留时间最短。从图1气温变化情况可知,野榆钱菠菜3种异型种子幼苗能够在-6~30 ℃条件下生存,说明其幼苗有较强的耐寒和耐高温性。
经观察(表1),褐色种子幼苗其子叶3月22日初见破土,23、24日大量破土,出苗数量大,出苗期3 d,出苗率74.30%,4月19日幼苗真叶出现,5月11日子叶脱落,存留时间50 d,子叶单独为幼苗提供养分的时间为28 d。
黑色大种子幼苗其子叶3月24日破土,出苗数量少,出苗期较长,4月13日以后未见破土,出苗期21 d,黑色大种子幼苗未见大量出苗迹象,出苗率6.58%,4月15日幼苗真叶出现,5月7日子叶脱落,存留44 d,子叶单独为幼苗提供养分的时间为22 d。
黑色小种子幼苗其子叶3月29日破土,零星出苗,出苗数量极少,出苗期17 d,出苗率1.55%,4月13日真叶出现,子叶5月6日脱落,存留38 d,子叶单独为幼苗提供养分的时间为16 d。

表1 野榆钱菠菜3种异型种子的出苗率及子叶的存留时间Table 1 The emergence ratio and the cotyledon retention time of three types of dimorphic A. aucheri seeds
注:同列不同字母表示不同类型幼苗之间差异显著(Duncan post-hoc 多重比较,P<0.05)。
Notes: Different lowercase letters in the same column indicate significant differences in different types of seedlings(Duncan post-hoc test;P<0.05).
2.2 异型种子幼苗的子叶形态及生长动态

图2 黑色小种子、黑色大种子、褐色种子幼苗的子叶长度和宽度 Fig.2 The cotyledon length and width of seedlings grown from small black seeds, big black seeds and brown seeds 图中不同小写字母表示同型幼苗不同时间的差异;不同大写字母表示不同型幼苗在同一时间的差异(Duncan post-hoc 多重比较,P<0.05)。下同。Different lowercase letters indicate significant differences in all stages; Different capital letters indicate significant differences among three types of seeds at same stage (Duncan post-hoc test; P<0.05). The same below.
3种异型种子幼苗子叶出土时均为两片舒展的长披针形绿色叶片,幼苗的子叶出土时大小不同,褐色种子幼苗子叶出土时长(1.14±0.12) cm,宽(0.32±0.04) cm,黑色大种子幼苗子叶出土时长(0.56±0.03) cm,宽(0.22±0.01) cm,黑色小种子幼苗子叶出土时长(0.35±0.02) cm,宽(0.15±0.02) cm(图2)。
图2反映了3种异型种子幼苗子叶的大小及生长速度,褐色种子幼苗子叶长度4月13日达到最大值,宽度4月23日达到最大值,黑色大种子幼苗和黑色小种子幼苗子叶长度和宽度均在4月23日达到最大值。
2.3 三种异型种子幼苗的生长动态和根冠比
由图3可知,褐色种子幼苗的根在早期明显长于两种黑色种子,4月3日测量时,褐色种子幼苗的根长为(2.516±0.42)cm,黑色大种子幼苗的根长为(1.128±0.21) cm,黑色小种子幼苗的根长为(0.774±0.22) cm,褐色种子幼苗的根长是黑色大种子的2.23倍,是黑色小种子幼苗根长的3.25倍。5月13日最后一次测量时,褐色大种子幼苗的根长为(7.892±0.86) cm,黑色大种子幼苗的根长是(7.188±0.65) cm,黑色小种子幼苗的根长为(6.008±0.92) cm,此时,相应的比例缩小为1.10和1.31倍。
由图4可知,3种异型种子幼苗的地上、地下生物量在4月3日至4月23日增长速率较为平缓,5月3日后增长速率显著增大,这可能与真叶的出现有关。对比3种异型种子的地上和地下生物量可知,在实验期内,3种异型种子幼苗地上生物量的差距在持续增大,地下生物量的差距在逐渐减小,这说明褐色种子幼苗主要将营养物质分配给地上部分,两种黑色种子幼苗将更多的营养物质分配给地下部分。

图3 黑色小种子、黑色大种子、褐色种子幼苗的根长动态Fig.3 The root length dynamic of seedlings grown from small black seeds, big black seeds and brown seeds
由图5可知,3种异型种子幼苗的根冠比动态有差异,褐色种子与黑色大种子幼苗的根冠比在试验期间呈下降趋势,黑色小种子幼苗根冠比的变化趋势为先增长后下降,其根冠比4月23日最高,为0.46;黑色大种子幼苗4月3日根冠比最高,为0.24;褐色种子幼苗4月3日最高,为0.27。至5月13日,黑色小种子幼苗根冠比为0.132,黑色大种子幼苗为0.092,褐色种子幼苗为0.088。

图4 黑色小种子、黑色大种子、褐色种子幼苗的地上(a)、地下(b)生物量动态Fig.4 The shoot (a) and root (b) biomass dynamic of seedlings grown from small black seeds, big black seeds and brown seeds

图5 黑色小种子、黑色大种子、褐色种子幼苗根冠比动态Fig.5 The root-top ratio dynamic of seedlings grown from small black seeds, big black seeds and brown seeds
3 讨论
3.1 异型种子的萌发物候及出苗率有差异
种子异型性是植物在适应时空高度异质性环境时进化出的一种器官的多态现象[7,21-23],具有重要的进化生态意义[5-6,24-26]。具种子异型性的植物其异型种子往往表现出不同的形态结构特征和生态行为特征[27-28]。
异型种子不同的形态结构特征通常使其具有不同的休眠类型和休眠程度[6,24,29-31],已有研究结果表明,有苞片包被的褐色种子没有休眠特性,无苞片包被的黑色种子具有休眠特性,本研究中,在相同温度、水分、土壤条件下,野榆钱菠菜3种异型种子萌发物候不同步,褐色种子较黑色大种子提前萌发2 d,较黑色小种子提前萌发7 d,且褐色种子的出苗率显著高于两种黑色种子的出苗率,出苗率高达74.3%,显著高于黑色大种子的6.58%和黑色小种子的1.55%。前人研究表明,种子异型性植物产生的异型种子通常只有一种种子形成长久的土壤种子库,其他种子形成短暂的土壤种子库[7],而根据本实验数据结果,野榆钱菠菜的黑色大种子和黑色小种子均形成长久的土壤种子库,褐色种子形成短暂的土壤种子库,此结果与另一种子异型性植物Atriplexsagittata相同[21],持久的土壤种子库能够持续更新地上种群,当地上种群因遭遇极端环境导致种群覆灭时保障种群的延续[17]。
异型种子具有不同的萌发策略,本研究中褐色种子在10~20 ℃左右即能大量萌发出苗(图1),黑色大种子与黑色小种子出苗期较长且出苗率低,这说明褐色种子的萌发属于“机会主义”的萌发策略,黑色大种子和黑色小种子的萌发属于“谨慎”的萌发策略,这种“机会主义”策略与“谨慎”策略在同一植株上共存的现象称之为“两头下注”策略, “两头下注”策略是荒漠植物在长期适应荒漠异质条件的过程中进化出的特殊生态机制,虽然褐色种子的“机会主义策略”出苗率高且集中出苗能快速建植种群,但在环境条件不可预测或不利的时候,这种冒险的、快速的萌发出苗对种群是有害的[5,7-8],而采取“谨慎”萌发策略的形成持久土壤种子库的两种黑色种子会在条件适宜时萌发出苗,保障种群的延续,这是种子异型性植物能够长期适应荒漠异质环境的重要生态适应机制[32-34]。
3.2 异型种子幼苗生长特性有差异,但逐渐缩小
异型种子产生的幼苗有差异,这种差异可能随植株的生长逐渐消失[5,35-37],本研究结果表明,褐色种子幼苗在初期较两种黑色种子幼苗大,其子叶、根、地上地下生物量均显著大于两种黑色种子幼苗(图2和图3),并且褐色种子幼苗的子叶单独为幼苗提供养分的时间长达28 d,而黑色小种子和黑色大种子幼苗的子叶单独为其幼苗提供养分的时间分别为16和22 d,这表明两种黑色种子通过缩短子叶存留时间尽早长出真叶的策略产生相对多的养分,为其生长创造更好的条件,这种策略能够最大限度地缩小其幼苗与褐色种子幼苗间的差异,这说明异型种子后代幼苗中较弱的幼苗(黑色种子幼苗)能够通过改变其供养方式使幼苗较快生长,增加抵御异质环境的能力,在相同的环境条件下,异型种子幼苗的这种不同供养策略可能是由遗传决定的,这是种子异型性植物在荒漠环境中进化出的又一重要适应机制。
3.3 异型种子幼苗对养分的分配策略有差异
植物根的生长状态直接影响着植物地上部分的生长及其生物量构成[38],对生长在荒漠生境中的植物来说,水分是最为重要的生长因子,较大、较强壮的根系意味着生存的机率更大,本研究数据结果表明,褐色种子幼苗的根长在4月3日测量时分别是黑色大种子和黑色小种子幼苗的2.23和3.25倍,在5月13日最后一次测量中,相应的比例缩小为1.10和1.31倍,这表明野榆钱菠菜3种异型种子幼苗根的差异在逐渐缩小,植物地下部分的生长状况直接影响着地上部分的生长状况[39],对植物的生长有着不可替代的作用,与植物对环境的适应能力、对种间竞争的抵抗能力密切相关[40],在相同环境条件下,两种黑色种子幼苗的根相对于褐色种子幼苗生长更快,这表明种子异型性植物产生的小种子幼苗能够通过根的快速生长增加其在荒漠异质条件中成功生存的机率。
根冠比是植物地下部分与地上部分的比值,是衡量植株光合作用产物的重要指标,也是估算植物地上生物量与地下生物量关系的重要参数[41-42],能够直观反映出植物体光合作用产物的分配比例[43]。本研究中,3种异型种子幼苗的根冠比动态有差异,褐色种子和黑色大种子幼苗的根冠比呈逐渐降低的状态,黑色小种子幼苗的根冠比先升高后降低,这表明野榆钱菠菜3种异型种子幼苗的生物量分配策略不同,黑色小种子幼苗在幼苗早期尽可能地将养分输送到地下部分,使其拥有较高的根冠比,根冠比的增长能够使植物增大对养分和水分的吸收量,使植物的抗旱能力增强,对植物适应不良环境具有重要的作用[43],同时,黑色小种子幼苗在3种异型种子幼苗中最早长出真叶,这样其幼苗的地上地下部分相辅相成,使幼苗能够更快地生长。褐色种子和黑色大种子,其种子相对较大,营养物质储存充分,萌发后幼苗、根相对较大,或许其根足以支撑地上部分的增长,所以褐色种子和黑色大种子幼苗在地上地下生物量分配方面更加侧重于地上,利用健壮的幼苗以抵抗荒漠环境可能的生存风险,保证种群的顺利生存[44],这种不同的生物量分配策略使种子异型性植物的种群在面对荒漠异质环境不同的环境状况时都有相应的应对方式,是种子异型性植物适应荒漠异质条件的重要生态机制。
4 结论
本研究以种子异型性植物野榆钱菠菜为研究对象,采用室外仿自然条件培育的方法在相同环境条件下对其3种异型种子的萌发物候、出苗率及异型种子的幼苗生长特性进行了研究,主要得出以下结论:1) 种子异型性植物产生的异型种子其萌发物候、幼苗出苗率及子叶存留时间有差异,大种子萌发早、出苗率高,子叶存留时间长;小种子萌发晚、出苗率低,子叶存留时间短。2)种子异型性植物“两头下注”策略产生的异型种子对种群的贡献不同,非休眠种子(褐色种子)以“机会主义”的萌发策略保证种群在条件适宜时快速建植,休眠种子(黑色种子)以“谨慎”的萌发策略确保种群在遭遇极端环境时保障种群的延续。3)种子异型性植物产生的异型种子其幼苗具有不同的生物量分配策略,非休眠种子(褐色种子)幼苗主要将养分输送到地上部分;休眠种子(黑色种子)幼苗通过缩短子叶存留时间、快速长出真叶以提高光合速率的方式将养分更多的分配到地下部分。异型种子幼苗这种不同的生物量分配策略使种子异型性植物在面对荒漠异质环境的不同状况时都有着不同应对方式,能最大限度地保障种群的成功生存和延续,是种子异型性植物能够长期适应荒漠异质环境的重要生态适应机制。
References:
[1] Zhang J G, Wang X P, Li X R,etal. Advances and prospect of researches on desert lift history strategies. Journal of Desert Research, 2005, 25(3): 306-314.
张景光, 王新平, 李新荣, 等. 荒漠植物生活史对策研究进展与展望. 中国沙漠, 2005, 25(3): 306-314.
[2] Kong L S, Guo K, Wang Q B. The characteristics of element content of dominant species in south Junggar desert, Xinjiang. Acta Ecologica Sinica, 2002, 22(8): 1202-1210.
孔令韶, 郭柯, 王其兵. 新疆南准噶尔荒漠优势植物的化学成分含量特点. 生态学报, 2002, 22(8): 1202-1210.
[3] Mao Z M. Flora Xinjiangensis. Vol.2(1). Urumqi: Xinjiang Science & Technology & Hygiene Publishing House, 1994: 84-106.
毛祖美.新疆植物志(第二卷第一册). 乌鲁木齐: 新疆科技卫生出版社, 1994: 84-106.
[4] Gutterman Y. Seed germination in desert plants. Berlin Heidelberg: Springer, 1993.
[5] Venable D L. The evolutionary ecology of seed heteromorphism. The American Naturalist, 1985, 126(5): 577-595.
[6] Imbert E. Ecological consequences and ontogeny of seed heteromorphism. Perspectives in Plant Ecology Evolution & Systematics, 2002, 5(1): 13-36.
[7] Wang L, Dong M, Huang Z Y. Review of research on seed heteromorphism and its ecological significance. Chinese Journal of Plant Ecology, 2010, (5): 578-590.
王雷, 董鸣, 黄振英. 种子异型性及其生态意义的研究进展. 植物生态学报, 2010, (5): 578-590.
[8] Lloyd D G. Variation strategies of plants in heterogeneous environments. Biological Journal of the Linnean Society, 1984, 21(4): 357-385.
[9] Meyers L A, Bull J J. Fighting change with change, adaptive variation in an uncertain world. Trends in Ecology and Evolution, 2002, 17(12): 551-557.
[10] Mandak B, Pysek P. The effects of light quality, nitrate concentration and presence of bracteoles on germination of different fruit types in the heterocarpusAtriplexsagittata. Journal of Ecology, 2001, 89(2): 149-158.
[11] Cheplick G P, Quinn J A.Amphicarpumpurshiiand the “pessimistic strategy” in amphicarpic annuals with subterranean fruit. Oecologia, 1982, 52(3): 327-332.
[12] Koller D, Roth N. Studies on the ecological and physiological significance of amphicarpy inGymanrrhenamicrantha(Compositae). American Journal of Botany, 1964, 51(1): 26-35.
[13] Xu P. Xinjiang grassland resources and its utilization. Urumqi: Xinjiang Science & Technology & Hygiene Publishing House, 1993.
许鹏. 新疆草地资源及其利用. 乌鲁木齐: 新疆科技卫生出版社, 1993.
[14] De Villiers A J, Van Rooyen M W, Theron G K,etal. Removal of sodium and chloride from a saline soil byMesembryanthemumbarklyi. Journal of Arid Environments, 1995, 29(3): 325-330.
[15] Zhao Z Y, Li Z S, Zhang F H,etal. Impacts of halophytes planting on salt balance in agricultural development region of Karamay city. Bulletin of Soil and Water Conservation, 2013, 33(4): 211-215.
赵振勇, 李中邵, 张福海, 等. 盐生植物种植对克拉玛依农业开发区盐分平衡的影响. 水土保持通报, 2013, 33(4): 211-215.
[16] Wei Y, Yan C, Yin L K. Seed polymorphism and ecotype ofAtriplexaucheri. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(3): 485-487.
魏岩, 严成, 尹林克. 野榆钱菠菜(Atriplexaucheri)的种子多型性及生态型. 西北植物学报, 2003, 23(3): 485-487.
[17] Wang L, Huang Z, Baskin C C,etal. Germination of dimorphic seeds of the desert annual halophyteSuaedaaralocaspica(Chenopodiaceae), a C4plant without kranz anatomy. Annals of Botany, 2008, 102(5): 757-769.
[18] Song J, Fan H, Zhao Y Y,etal. Effect of salinity on germination, seedling emergence, seedling growth and ion accumulation of a euhalophyteSuaedasalsain an intertidal zone and on saline inland. Aquatic Botany, 2008, 88(4): 331-337.
[19] Wei Y, Dong M, Huang Z Y. Seed polymorphism, dormancy and germination ofSalsolaaffinis(Chenopodiaceae), a dominant desert annual inhabiting the Junggar Basin of Xinjiang, China. Australian Journal of Botany, 2007, 55(4): 1046-1053.
[20] Mandak B, Pysek P. How does seed heteromorphism influence the life history stages ofAtriplexsagittata(Chenopodiaceae). Flora-Morphology, Distribution, Functional Ecology of Plants, 2005, 200(6): 516-526.
[21] Mandak B, Pysek P. Fruit dispersal and seed banks inAtriplexsagittata: the role of heterocarpy. Journal of Ecology, 2001, 89(2): 159-165.
[22] Brändel M. Dormancy and germination of heteromorphic achenes ofBidensfrondosa. Flora, 2004, 199(3): 228-233.
[23] Brändel M. Ecology of achene dimorphism inLeontodonsaxatilis. Annals of Botany, 2007, 100(6): 1189-1197.
[24] Mandák B. Seed heteromorphism and the life cycle of plants: a literature review. Preslia, 1997, 69: 129-159.
[25] Mohamed-Yasseen Y, Costanza S. The role of seed coats in seed viability. The Botanical Review, 1994, 60(4): 426-439.
[26] Vleeshouwers L M, Bouwmeester H J. A simulation model for seasonal changes in dormancy and germination of weed seeds. Seed Science Research, 2001, 11(1): 77-92.
[27] Khan M A, Ungar I A. Seed polymorphism and germination responses to salinity stress inAtriplextriangularisWilld. International Journal of Plant Sciences, 1984, 145(4): 487-494.
[28] Philipupillai J, Ungar I A. The effect of seed dimorphism on the germination and survival ofSalicorniaeuropaeaL. populations. American Journal of Botany, 1984, 71(4): 542-549.
[29] Li W Q, Liu X J, Mao R Z,etal. Advances in plant seed dimorphism (or polymorphism) research. Acta Ecologica Sinica, 2006, 26(4): 1234-1242.
李伟强, 刘小京, 毛任钊, 等. 植物种子二形性(多形性)研究进展. 生态学报, 2006, 26(4): 1234-1242.
[30] Liu P W, Wei Y. Seed dimorphism and germination behavior ofAtriplexmicrantha, an annual inhabiting Junggar desert. Acta Ecologica Sinica, 2007, 27(10): 4233-4239.
刘鹏伟, 魏岩. 准噶尔荒漠异苞滨藜(Atriplexmicrantha)的种子二型性及其萌发行为. 生态学报, 2007, 27(10): 4233-4239.
[31] Yao S X, You T Y, Xu D S,etal. Seed heteromorphism and germination mechanism ofChenopodiumalbumin Xinjiang arid region. Acta Ecologica Sinica, 2010, 30(11): 2909-2918.
姚世响, 油天钰, 徐栋生, 等. 新疆干旱区植物藜的种子异型性及其萌发机理. 生态学报, 2010, 30(11): 2909-2918.
[32] Friedman J, Stein Z. The influence of seed-dispersal mechanisms on the dispersion ofAnastaticahierochuntica(Cruciferae) in the Negev Desert, Israel. Journal of Ecology, 1980, 68(1): 43-50.
[33] Ungar I A. Population ecology of halophyte seeds. The Botanical Review, 1987, 53(3): 301-334.
[34] Wei M H, Huang J H. Seed polymorphism and germination behavior ofSalicorniaeuropaeainhabiting in the area of Ayding Lake, Xinjiang, China. Pratacultural Science, 2015, 32(12): 2064-2070.
魏梦浩, 黄俊华. 艾丁湖盐角草种子异型性及萌发特性. 草业科学, 2015, 32(12): 2064-2070.
[35] Khan M A, Ungar I A. The effect of salinity and temperature on the germination of polymorphic seeds and growth ofAtriplextriangularisWilld. American Journal of Botany, 1984, 71: 481-489.
[36] Imbert E. Seed heteromorphism inCrepissancta(Asteraceae): Performance of two morphs in different environments. Oikos, 1997, 79(2): 325-332.
[37] Li X R, Yao S Y, Chen S S,etal. Physiological responses to salt stress of plants derived from heteromorphic seeds ofChenopodiumalbum. Acta Ecologica Sinica, 2015, 35(24): 8139-8147.
李晓荣, 姚世响, 陈莎莎, 等. 藜异型性种子后代植株盐响应生理机制. 生态学报, 2015, 35(24): 8139-8147.
[38] And W J D, Zhang J. Root signals and the regulation of growth and development of plants in drying soil. Annual Review of Plant Biology, 2003, 42(1): 55-76.
[39] Yang X G, Fu H, Zhang H R,etal. Effects of water stress on leaf water potential and biomass at seedling stage ofZygophyllumxanthoxylum. Acta Prataculturae Sinica, 2006, 15(2): 37-41.
杨鑫光, 傅华, 张洪荣, 等. 水分胁迫对霸王苗期叶水势和生物量的影响. 草业学报, 2006, 15(2): 37-41.
[40] Sun X, Yu Z. A study on root system ofNitrariatangutorum. Journal of Desert Research, 1992, 12(4): 50-54.
孙祥, 于卓. 白刺根系的研究. 中国沙漠, 1992, 12(4): 50-54.
[41] Peng Y Q, Zhou F, Wan H J,etal. The response of seedling and photosynthetic characteristics ofSuaedasalsaandAtriplextriangularisto different salinity. Acta Prataculturae Sinica, 2012, 21(6): 64-74.
彭益全, 周峰, 万红建, 等. 碱蓬和三角叶滨藜幼苗生长光合特性对不同盐度的响应. 草业学报, 2012, 21(6): 64-74.
[42] Huang D Q, Yu L, Zhang Y S,etal. Study on root-shoot ratios of natural grasslands and their relationships with climatic factors in the northern slopes of the Qilian Mountains. Arid Zong Research, 2011, 28(6): 1025-1030.
黄德青, 于兰, 张耀生, 等. 祁连山北坡天然草地根冠比与气候因子的关系. 干旱区研究, 2011, 28(6): 1025-1030.
[43] Huang C B, Zeng F J, Lei J Q,etal. Effects of irrigation on plant growth and nitrogen use characteristics ofCalligonumcaputmedusaeSchrenk seedlings. Acta Ecologica Sinica, 2014, 34(3): 572-580.
黄彩变, 曾凡江, 雷加强, 等. 灌溉对沙拐枣幼苗生长及氮素利用的影响. 生态学报, 2014, 34(3): 572-580.
[44] Fenner M. Relationships between seed weight, ash content and seedling growth in twenty-four species of compositae. New Phytologist, 1983, 95(4): 697-706.
