新疆博格达山岩面生地衣群落结构特征
2018-03-22艾尼瓦尔吐米尔热衣木马木提阿不都拉阿巴斯
艾尼瓦尔·吐米尔,热衣木·马木提,阿不都拉·阿巴斯
新疆大学生命科学与技术学院, 乌鲁木齐 830046
地衣群落研究的目标在于深入地揭示地衣群落的结构、物种多样性、生态、动态、分类及其在不同生态系统中的分布等基本规律[1- 2]。地衣群落结构特征是地衣群落的基本属性也是认识群落的组成、变化和发展趋势的基础。一般地衣群落结构反映了群落对环境的适应,演替和动态机制,进而有助于地衣群落的分类和有效保护。物种多样性是生物多样性在物种水平上的表现形式,生态学方面物种分布的均匀程度,一般是从群落组织水平上进行研究,也称为群落多样性[4- 5]。群落物种多样的研究是遗传多样性和生态系统多样性研究的基础,是一个群落结构和功能复杂性的量度,是反映群落特征的重要指标之一[2,6]。因此,进行群落物种多样性研究不仅能更好地反映群落在组成、结构、功能和动态等方面的异质性,也可反映不同自然地理条件与群落的相互关系及其发展变化[5- 6]。分析地衣群落的结构和物种多样性对揭示地衣群落的更新、稳定性与演替规律具有极为重要的意义[2]。
岩面生地衣是陆地生态系统生物多样性的主要成分之一,定居在岩石上,腐蚀岩石形成土壤,改变岩石的微环境,为其他植物的定居,种群和群落的形成提供栖息地,从而促进植物群落的演替[1,7]。国外地衣生态学研究历史较长,在岩面生地衣与岩石的关系、岩面生地衣群落结构、物种分布等方面进行了不少的定量研究[8- 11]。在国内近年来随着植物生态学定量研究的发展,也有学者尝试采用多元分析方法对地面生、树生和岩面生地衣群落进行定量研究[12- 14]。其中我们长期以来在新疆乌鲁木齐南部山区、阿尔泰山两河源国家级自然保护区和托木尔峰国家级自然保护区等地区应用双向指示种分析(TWINSPAN)、除趋势对应分析(DCA)、典范对应分析(CCA),除趋势典范对应分析(DCCA)等对岩面生地衣群落的分类,群落物种分布格局、地衣群落和生态环境之间的生态关系进行了定量研究[15- 20]。
天池博格达峰国家级自然保护区在极短的水平距离内具有天山北坡典型的山地垂直自然带谱,是全球温带干旱区山地垂直带的典型代表,是中国生物多样性保护优先区域[21- 22]。目前有关天池博格达峰国家级自然保护区岩面生地衣群落结构方面尚未见定量研究报道。因此,定量研究该地区岩生地衣群落结构特征以及地衣物种分布与环境因子的关系,对于探明该地区地衣群落结构和生态分布特征提供一定的科学依据,同时认识干旱区地衣植物的分布格局具有一定的参考价值。
1 方法
1.1 研究区概况
天池博格达峰国家级自然保护区位于中国新疆维吾尔自治区阜康市境内,东天山博格达峰北麓,准噶尔盆地古尔班通古特沙漠的南缘,地处88°00—88°20E,43°45—43°59N,总面积38069hm2。其中:核心区面积18146hm2,缓冲区7162hm2,实验区12761hm2。是新疆维吾尔自治区第一个、中国第七个被纳入联合国“人与生物圈”网络的保护区。博格达峰北邻准噶尔盆地,南隔达坂城谷地与吐鲁番盆地遥遥相望,处于荒漠包围之中。由于山体高大,在3500m以上高山地区北冰洋来的湿气流被受阻,故气候湿润多雨,年降水量600—700mm,年均温-6℃,雪线以上低于-9℃。保护区的核心区处在天山的逆温层中,气温温和稳定,年均温2℃;年降水量497.5mm。保护区种子植物有200多种。植被垂直分布明显,自上而下可分为:高山垫状植被带、高山草甸带、亚高山草甸带、山地森林带、山地草原带、河谷落叶阔叶林。森林以原始雪岭云杉林为主,广布于中山带阴坡与河谷[21- 22]。
1.2 野外调查
调查样点的设立:在博格达峰自然保护区选择有代表性的地段根据不同的海拔、地形和植被类型设置了20m×20m的20个样点。调查样点内岩石的海拔高度、坡向、坡度、光照强度、岩石大小、岩石pH、人为干扰等[8- 9,16,18]。
岩面生地衣群落的调查:在每个样点中选择表面较扁平,坡角0°到90°之间,面积≥2500cm2(50cm×50 cm)的岩石,设立20cm×20cm的小样方1至5个,每个小样方由256个1.25cm×1.25cm的小方格组成。记录小样方内出现的每个地衣物种及其所占网格数,同时采集地衣标本。地衣种类盖度以地衣物种出现的网格数占网格总数的百分数表示[9,16,18]。光照强度(岩石表面接受的太阳直射辐照量)根据岩石的坡向、坡度和纬度,采用Gates(1956)和Campbell(1998)的公式计算[9,23- 24]。
S(岩石斜面)=S(岩石水平面)×cos (i)
S(岩石水平面)=S(p)×sin(ø)
cos(i)= cos(β)cos(z)+sin(z)sin(β)cos(α-η)
sin(ø)=sin(λ) sin(δ)+cos(λ) cos(δ) cos(15t)
式中,S:太阳直射辐照量;η:岩石的坡向;λ:纬度;δ:太阳赤纬;t:太阳中午时间;z:太阳天顶角;α:太阳方位角;β:岩石坡度。
光照强度的划分标准为:S<80 (最弱)、 80 ≤S<120 (弱)、120 ≤S<160 (中等)、160 ≤S<200 (强)、S≥ 200 (最强)。根据各样点周围的旅游设施、放牧、污染源的距离等目测人为干扰的强度。各样点的环境概况和环境变量的分级见表1。对所采集的地衣标本在实验室采用形态解剖观察和显色反应方法进行物种鉴定[25- 27]。
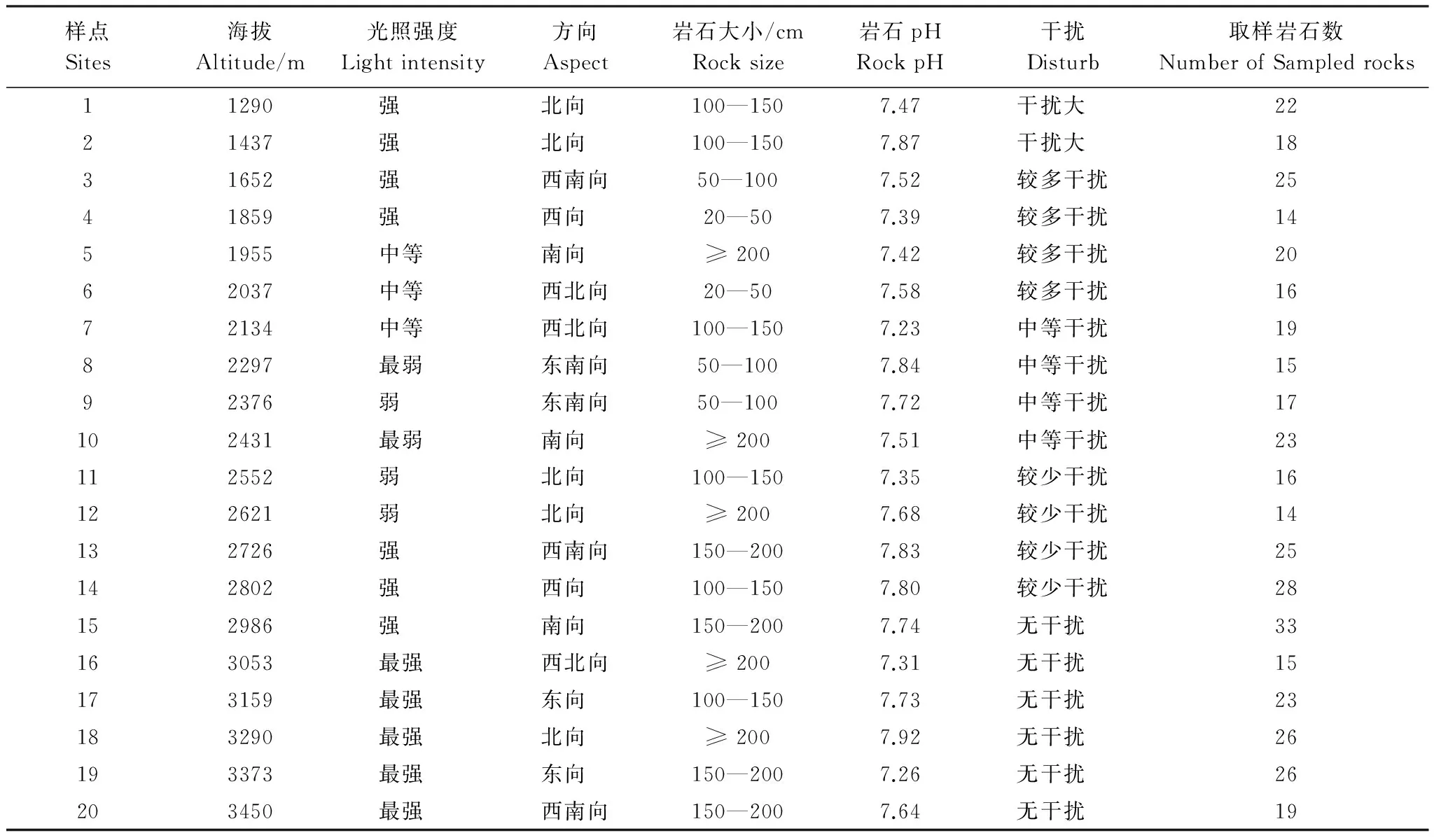
表1 博格达山20个样点的环境变量及其等级
1.3 数据分析
本文以样点为对象,以岩面生地衣的盖度为指标,37个岩面生地衣种 (表2)和20个样点构成37×20的矩阵(附录Ⅰ)。

表2 博格达山岩面生地衣物种组成
应用双向指示种分析方法(TWINSPAN)和除趋势对应分析法(DCA)进行岩面生地衣群落数值分类[28-29]。TWINSPAN分析的参数设置为,假种切割水平采用5级,合计水平值分别为0、2、5、10、20;分类最小种组采用5个;每次分类的指示种最大数量为10个;最终结果矩阵显示的最大物种为699;分类的最大水平数为6次;各假种水平的加权值分别为1、1、2、2、3。TWINSPAN分类图左边数字为地衣种编号和拉丁名缩写,最上方为样点编号,最下方是样点分组结果,最右边是地衣分类结果,中间数字为分级水平,“—”表示地衣种在该样方不存在,“1”表示一次分类结果,“0”表示另一组[30- 31]。DCA排序时对数据进行对数转换,未处理稀有种,选择Kaiser′s rule确定排序轴。采用典范对应分析(CCA)对岩面生地衣种类的分布格局进行分析。CCA分析时,由除趋势对应分析(DCA)得出地衣种的单峰响应值(梯度长度SD)。当SD>2时,即可进行CCA分析。对标准化后的地衣种类盖度数据进行卡方处理;环境因子采用最大值法进行标准化处理[8,16,18,29]。地衣群落物种多样性指数和均匀度指数能够客观地反映群落内物种组成以及物种在各群落内部的分布特征[2],而群落相似性指数是比较不同群落相似性关系的重要参数[2,32]。本文我们根据岩面生地衣的种类和盖度,计算岩面生地衣群落的Shannon-Wiener多样性指数、Simpson′s 多样性指数、Margalef丰富度指数、Patrick丰富度指数和Pielou均匀度指数,比较各群落的多样性。计算Sörensen′s相似性指数和Jaccard′s相似性指数,比较各群落间的相似性特点。
Shannon-Wiener 多样性指数(H′)计算公式[32-34]:
式中,pi为第i物种相对盖度的百分比,pi=ni/N,其中ni为第i物种的相对盖度;N为s个种的相对盖度之和。
Simpson′s 多样性指数计算公式(D):
式中,S为物种数量,pi为第i物种相对盖度的百分比。
Margalef 丰富度指数(DMa):
式中,S为群落中物种相对盖度的之和,N为群落中观察到的物种总相对盖度。
Patrick丰富度指数:Dp=S,其中Dp为丰富度指数;S为群落内的物种数[32-34]。
式中,J为均匀度指数,H′为实测多样性值,Hmax为理论上群落多样性的最大值Hmax=log2S,其中S为总物种数[32-34]。
式中,S12为两个群落共有的物种数;S1(S2) 为群落1 (群落2)中的物种数。
式中,S12为两个群落都出现的物种数;S1(S2) 为群落1 (群落2)中的物种数。
应用WinTWINS 2.3统计软件进行双向指示种分析,利用MVSP 3.13d for Windows软件包进行除趋势对应和典范对应分析。

图1 TWINSPAN分类结果矩阵图 Fig.1 The matrix of species and sampling site classification by TWINSPAN 缩写参数,参见表2
2 结果
2.1 岩面生地衣群落数量分类
以岩面生地衣盖度为指标,对研究区域的20个样点进行TWINSPAN与DCA分析,分别得到图1与图2。从TWINSPAN图上,20个样点依据岩面生地衣盖度指标,可被分成5个岩面生地衣群丛。
群丛1:包括样点2、11、12和13,除了样点2分布在海拔1400m外,其他样点分布在海拔2500—2750m之间。主要地衣种类有斑纹网衣(Lecideatessellata)、杜瑞氏黄梅(Xanthoparmeliadurietzii) 、粉芽盾衣(Peltulaeuploca)、盾脐鳞衣(Rhizoplacapeltata) 、包氏饼干衣(Rinodinabohlinii)、粉盘平茶渍(Aspiciliaalphoplaca)、白边平茶渍(Aspiciliasublaqueata)、伴藓大孢蜈蚣衣(Physconiamuscigena)等25种。地衣总平均覆盖度为30.145%,盖度最大的是斑纹网衣,为4.275%;其次为粉芽盾衣,盖度为3.803%。命名为斑纹网衣+粉芽盾衣+杜瑞氏黄梅群丛。
群丛2:包括样点1、3、4和样点7,分布在海拔1290—2150m之间。主要地衣有袋衣(Hypogymniaphysodes)、白边平茶渍(Aspiciliasublaqueata)、墙茶渍(Lecanoramuralis)、砖孢胶衣(Collemasubconveniens)、丽石黄衣(Xanthoriaelegans)、粉盘平茶渍(Aspiciliaalphoplaca)、杜瑞氏黄梅(Xanthoparmeliadurietzii)等17个种。地衣总平均盖度为15.885%,盖度最大的是袋衣,为2.533%;其次白边平茶渍,为1.933%。命名为袋衣+白边平茶渍+砖孢胶衣群丛。
群丛3:包括样点5、6、8、9、10、14。分布在海拔1950—2800m之间的针阔叶林混合林带和针叶林带。主要地衣有聚茶渍(Lecanoraaccumulata)、丽石黄衣(Xanthoriaelegans)、袋衣(Hypogymniaphysodes)、亚洲平茶渍(Aspiciliaasiatica) 、蓝灰蜈蚣衣(Physciacaesia)、包氏饼干衣(Rinodinabohlinii)、中华石果衣(Endocarponsinense)等30个种。地衣总平均盖度为37.87%,盖度最大的是聚茶渍,为3.877%,其次丽石黄衣,为2.597%。命名为聚茶渍+丽石黄衣+亚洲平茶渍群丛。
群丛4:包括样点15、18、19。分布在海拔2900—3400m之间。主要地衣有中华石果衣(Endocarponsinense)、伴藓大孢蜈蚣衣(Physconiamuscigena)、垫脐鳞衣(Rhizoplacamelanophthalma)、杜瑞氏黄梅(Xanthoparmeliadurietzii)、粉芽盾衣(Peltulaeuploca) 、绿黑地图衣(Rhizocarponviridiatrum)、短绒皮果衣(Dermatocarponvellereum)、包氏饼干衣(Rinodinabohlinii)等24个种。地衣总平均盖度为30.458%,其中中华石果衣的盖度最大为5.26%,其次为伴藓大孢蜈蚣衣为,3.806%。命名为中华石果衣+伴藓大孢蜈蚣衣+垫脐鳞衣群丛。
群丛5:包括样点16、17和20,分布在海拔3000—3500m之间。主要地衣种类有石胶衣(Collemaflaccidum)、短绒皮果衣(Dermatocarponvellereum)、绿黑地图衣(Rhizocarponviridiatrum)、伴藓大孢蜈蚣衣(Physconiamuscigena) 、砖孢胶衣(Collemasubconveniens)、翘白角衣(Siphulapteruloides)、东方黄梅(Xanthoparmeliaorientalis)等18个种。地衣总平均盖度为19.331%,盖度最大的是石胶衣,为2.332%;其次为短绒皮果衣,为1.891%。命名为石胶衣+短绒皮果衣+绿黑地图衣群丛。
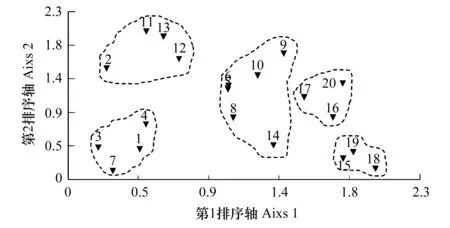
图2 20个样点的除趋势对应排序图Fig.2 DCA ordination diagram of 20 sampling sites1—20表示样点序号
TWINSPAN以间断形式对20个样点进行了划分,DCA排序以连续形式表明20个样点间岩面生地衣种类组成上的关系。DCA排序结果表明,第一排序轴的特征值为12.441,累计贡献率为43.289,第二排序轴的特征值为4.035,累计贡献率为57.331;而第三排序轴的特征根为2.450,累计贡献率为65.856,说明20个样点在时空分布上得到了很好的排序效果,岩面生地衣群落类型在排序图上的分类与TWINSPAN分类结果吻合。
2.2 岩面生地衣群落物种多样性
对5个岩面生地衣群落进行了Shannon-Wiener多样性指数、Simpson′s 多样性指数、Margalef丰富度指数和Patrick丰富度指数的分析(表3)。从表3可以看出,群丛3的多样性最大为4.357,其次为群丛1,为4.025;群丛2的多样性最低为3.196。物种在群丛1和群丛3的分布比较均匀,均匀度分别为0.866和0.887。
Jaccard′s相似性指数和Sörensen′s相似性指数的分析表明(表4),群落1和群落4的相似性最大分别为0.815和0.898,其次为群落1和群落5,为0.433和0.605;群落2和群落3的相似性最低为0.321和0.486。
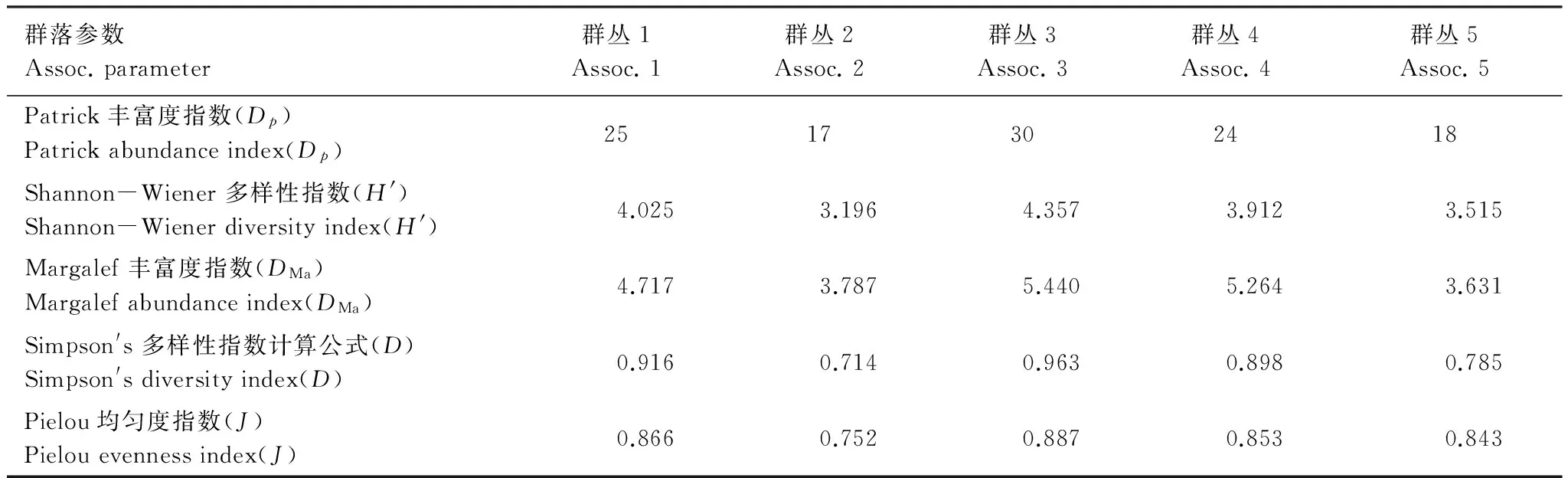
表3 岩面生地衣群落多样性和均匀度指数
群丛1—群丛5代表5个不同的岩面生地衣群落

表4 岩面生地衣群落相似性指数
2.3 分布格局与环境因子的关系
为了确定影响岩面生地衣物种分布的自然和人为因素,对37种岩面生地衣与环境参数数据进行了CCA分析。CCA排序结果见图3,排序图中用圆形、正三角形、菱形和倒三角形来表示各象限的岩面生地衣种类。从CCA排序图可见,第一象限的种类的分布与海拔、岩石pH、岩石大小等有关。其中垫脐鳞衣Rh.melanophthalma(10)、小茶渍L.hageni(13)、朝鲜黄梅X.coreana(18)、伴藓大孢蜈蚣衣Ph.muscigena(23)等种类分布在高海拔、光照较强和pH值较高的裸露的岩石表面。地图衣Rh.geographicum(27)、淡肤根石耳U.verginis(28)、灰黄枝衣T.lacunosus(31)和中华石果衣E.sinense(36)等种类分布在中等海拔地区,较大的岩石表面,但对岩石pH的要求不高。亚凹网衣L.subconcava(15)和包氏饼干衣R.bohlinii(24)的分布与岩石坡向的关系较大。

图3 岩面生地衣分布与环境因子关系的CCA排序图Fig.3 CCA revealing the relationships of the saxicolous lichens species with environmental factors
第二象限中糙聚盘衣G.scabra(1)、黑绿胶衣C.fuscovirens(3)、斑纹网衣L.tessellata(16)、蓝灰蜈蚣衣Ph.caesia(22)、石橙衣C.saxicola(30)、粉芽石黄衣X.sorediata(33)、盾脐鳞衣Rh.peltata(14)、丽石黄衣X.elegans(32)、聚茶渍L.accumulata(9)等种类广泛的分布在不同海拔的岩石表面;而粉芽盾衣P.euploca(29)、白边平茶渍A.sublaqueata(7)和粉盘平茶渍A.alphoplaca(5)较多的分布在光照较弱的低和中海拔地区的岩石。第三象限的亚洲平茶渍A.asiatica(6)和青海茶渍L.kukunorensis(11)分布在人为干扰较大的区域,小疣巨孢衣M.verrucosa(8)、墙茶渍L.muralis(12)、袋衣H.physodes(17)、翘白角衣S.pteruloides(37)分布在干扰较少的中海拔地区;杜瑞氏黄梅X.durietzii(19)、东方黄梅X.orientalis(20)和皮果衣D.miniatum(34)分布在人为干扰较少的地带中。第四象限中地衣种类的分布受到光照的影响,其中石胶衣C.flaccidum(2)矮饼干衣R.demissa(25)、绿黑地图衣Rh.viridiatrum(26)和短绒皮果衣D.vellereum(35)等种类分布在光照强的高海拔和裸露,植被郁闭度较低的地带。砖孢胶衣C.subconveniens(4)和菊叶黄X.somloensis(21)分布在中等海拔,干扰较少的地带。
在CCA排序图中与第一排序轴的相关系最大的是海拔高度(R=0.810);其次是光照强度(R=0.729);再次是岩石pH(R=0.716)。与第一排序轴相关性最小的是坡向(R=0.021),其次为岩石大小(R=0.322);人为干扰与第一排序轴的相关性为负值(R= -0.793)。与第二排序轴相关性最大的是坡向(R=0.473),其次为岩石大小(R=0.353),与岩石pH和海拔的相关性较小,分别为0.259和0.185;第二排序轴与光照强度和人为干扰的相关性为负值分别为-0.446和-0.336。从相关性分析可知,博格达山岩面生地衣群落种类的分布主要受到海拔高度、光照强度、岩石pH等因素的影响和限制;而岩石大小、坡向和人为干扰等因素的影响不大,从而形成了不同海拔岩面生地衣群落的地带性分布格局。
3 讨论
岩面生地衣作为先锋生物在植物群落原生演替早期首先定居在裸露而条件极差的岩石环境,经常受到剧烈变化的极端温度、水分和养分含量无规则波动和变化的影响。国外已有的研究表明,岩面生地衣群落的结构和物种组成随着海拔、坡度、光照、养分丰富度、岩石理化性质、雪的盖度和岩石表面微地形而发生变化[10,35-37]。Matthes等在加拿大安大略省Niagara Escarpment生物圈保护区采用DCA、CCA和PCCA等群落多元排序方法,研究了12个环境变量与石生地衣群落物种空间分布格局的关系[10]。结果发现,研究地区的石生地衣群落的物种组成受到岩石坡度、坡向、光照、湿度等因素小尺度变化的影响,同时发现由于受到人为干扰的影响叶状和枝状地衣种类组成从北到南呈现出梯度性减少的格局。Bjelland在挪威西部Vingen地区研究植被盖度、光照、栖息地微环境变量(岩石大小、岩石表面粗糙度、岩石表面凹度)和海洋因素对岩面生地衣物种组成的影响时发现,岩面生地衣群落的种类组成很少受到栖息地微环境变量的影响,但植被盖度对地衣群落的影响大于光照、海洋因素等的影响[36]。艾尼瓦尔·吐米尔等研究乌鲁木齐南部山区森林生态系统岩面生地衣群落时发现,海拔1100—2200m之间的阔叶针叶混交林带湿润石生地衣群落占优势;在海拔2300—2800m的植被高度比较低,太阳辐射强烈,湿度较低地区形成了干燥石生地衣群落。而海拔2800m以上的高山岩石上,耐寒、耐旱的地衣种类较多,形成了高山石生地衣群落[16]。艾尼瓦尔·吐米尔等报道阿尔泰山两河源自然保护区和托木尔峰国家级自然保护区的岩面生地衣群落的物种组成和分布具有海拔地带性分布特征[18,20]。
本研究结果表明,在博格达山分布在不同海拔的岩面生地衣群落多样性和相似性间存在差异。分布在低中海拔地区的群丛2中岩面生地衣种类的组成和分布除了受到自然因素的影响外,还受到环境污染等人为因素的影响,因此物种多样性较低。群丛1、群丛3和群丛4基本分布在中高海拔地区,因为该地带的岩石表面湿度较好,光照强度适中、岩石腐蚀程度高,所以物种多样性最高。群丛5分布在高山垫状植被带,该地带光照强烈、降水量丰富、温度低,风速高,地衣种类主要由耐旱、耐寒的壳状地衣组成,地衣种类的分布受到海拔、光照和岩石pH的影响较大。Allen 等在欧洲的研究表明,岩面生地衣群落的种类组成和丰富度随着岩石的坡向而发生变化[38]。MacDonald等在加拿大Nova Scotian沿海地区对岩面生地衣群落研究时发现,岩石朝着海洋的一面上岩面生地衣的丰富度和盖度低于相对面[39]。本研究中我们发现在博格达山地区岩石的坡度可以改变岩石表面的曝光度和接受的太阳直射福照度,从而导致岩石表面温度的变化,这些变化影响岩面生地衣种类组成和分布。人为干扰主要是通过改变微环境的结构和质量,导致环境污染而影响地衣代谢活动,进而导致地衣体的损害[40-41]。天池博格达山国家级自然保护区是新疆主要的旅游景点之一,科学合理的发展和管理旅游业,减少保护区内的各种人为干扰是有利于提高该地区的环境质量和生态环境的保护。同时将有效的提高该保护区的岩面生地衣物种多样性方面具有重要意义。因此,我们建议在该保护区今后应该加强对地衣群落物种组成及分布的定期研究,建立长期评价该地区环境质量变化和生物多样性动态的以地衣作为指示生物的生物学评价机制是非常必要的。
[1] James P W, Hawksworth D L, Rose F. Lichen communities in the British Isles: a preliminary conspectus // Seaward M R D, ed. Lichen Ecology. London: Academic Press, 1977: 296- 413.
[2] Schoenwetter T. Epiphytic Lichen Communities on Mountain Beech, Nothofagus Solandri var. Cliffortioides in Craigieburn Forest Park, South Island, New Zealand[D]. New Zealand: Lincoln University, 2010.
[3] Ray D G, Barton J W, Lendemer J C. Lichen community response to prescribed burning and thinning in southern pine forests of the mid-Atlantic coastal plain, USA. Fire Ecology, 2015, 11(3): 14- 32.
[4] 王永健, 陶建平, 彭月. 陆地植物群落物种多样性研究进展. 广西植物, 2006, 26(4): 406- 411.
[5] Primack R B. Essentials of Conservation Biology. 5th ed. Sunderland, MA: Sinauer Associates, Inc., 2010: 1- 215.
[6] 袁蕾, 周华荣, 宗召磊, 叶琴. 乌鲁木齐地区典型灌木群落结构特征及其多样性研究. 西北植物学报, 2014, 34(3): 595- 603.
[7] Armstrong R A, Welch A R. Competition in lichen communities. Symbiosis, 2007, 43(1): 1- 12.
[8] John E A. Fine scale patterning of species distributions in a saxicolous lichen community at Jonas Rockslide, Canadian Rocky Mountains. Ecography, 1990, 13(3): 187- 194.
[9] John E, Dale M R T. Environmental correlates of species distributions in a saxicolous lichen community. Journal of Vegetation Science, 1990, 1(3): 385- 392.
[10] Matthes U, Ryan B D, Larson D W. Community structure of epilithic lichens on the cliffs of the Niagara Escarpment, Ontario, Canada. Plant Ecology, 2000, 148(2): 233- 244.
[11] Pastore A I, Prather C M, Gornish E S, Ryan W H, Ellis R D, Miller T E. Testing the competition-colonization trade-off with a 32-year study of a saxicolous lichen community. Ecology, 2014, 95(2): 306- 315.
[12] Li S, Liu W Y, Wang L S, Ma W Z, Song L. Biomass, diversity and composition of epiphytic macrolichens in primary and secondary forests in the subtropical Ailao Mountains, SW China. Forest Ecology and Management, 2011, 261(11): 1760- 1770.
[13] Li S, Liu W Y, Li D W. Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystems in southwest China. Ecological Indicators, 2013, 29(6): 93- 104.
[14] 王玉良, 郑爱琴, 阿不都拉·阿巴斯. 鹞落坪树生地衣群落数量分类及多样性. 广西植物, 2011, 31(6): 747- 753.
[15] 艾尼瓦尔·吐米尔, 阿不都拉·阿巴斯. 新疆阿勒泰山两河源自然保护区地面生地衣的物种多样性. 生物多样性, 2006, 14(5): 444- 450.
[16] 艾尼瓦尔·吐米尔, 阿不都拉·阿巴斯. 乌鲁木齐南部山区岩面生地衣群落特征的研究. 菌物学报, 2009, 28(2): 178- 188.
[17] 艾尼瓦尔·吐米尔, 阿地力江·阿不都拉, 阿不都拉·阿巴斯. 乌鲁木齐南部山区地生地衣群落分布格局. 生物多样性, 2011, 19(5): 574- 580.
[18] 艾尼瓦尔·吐米尔, 阿不都拉·阿巴斯. 新疆托木尔峰国家级自然保护区岩面生地衣群落数量分类及其与环境相关性. 植物资源与环境学报, 2015, 24(2): 80- 87.
[19] 艾尼瓦尔·吐米尔, 阿不都拉·阿巴斯. 阿尔泰山两河源自然保护区森林生态系统朽木生地衣群落的数量分类. 菌物学报, 2015, 34(3): 357- 365.
[20] 艾尼瓦尔·吐米尔, 阿迪力江·阿不都拉, 阿不都拉·阿巴斯. 新疆阿尔泰山两河源自然保护区岩面生地衣群落特征的研究. 干旱区资源与环境, 2015, 29(9): 74- 79.
[21] 张元明, 曹同, 潘伯荣. 新疆博格达山地面生苔藓植物物种多样性研究. 应用生态学报, 2003, 14(6): 887- 891.
[22] 中国科学院植物研究所, 中国科学院新疆综合考察队. 新疆植被及其利用. 北京: 科学出版社, 1978: 92- 223.
[23] Gates D M. Radiant energy, its receipt and disposal // Waggoner P E, Gates D M, Webb E K, van Wijk W R, Businger J A, Crawford T V, Hendershott C H, Moss D N, Johnson H D, Chepil W S, Henson W R, Gardner W R, Slavík B, Thornthwaite C W, Hare F K, McQuigg J D, eds. Agricultural Meteorology. Meteorological Monographs. Boston, MA: American Meteorological Society, 1965, 6(28): 1- 26.
[24] Campbell G S, Norman J M. An Introduction to Environmental Biophysics. 2nd ed. New York: Springer-Verlag, 1997: 247- 277.
[25] 阿不都拉·阿巴斯, 吴继农. 新疆地衣. 乌鲁木齐: 新疆科技卫生出版社, 1998: 25- 161.
[26] Dobson F S. Lichens—— An Illustrated Guide to the British and Irish Species. 6th ed. Slough: The Richmond Publishing Co, 2011: 1- 496.
[27] Brodo I M. Keys to Lichens of North America: Revised and Expanded. New Haven: Yale University Press, 2016: 1- 424.
[28] Kooch Y, Jalilvand H, Bahmanyar M A, Pormajidian M R. Application of two way indicator species analysis in Lowland plant types classification. Pakistan Journal of Biological Sciences, 2008, 11(5): 752- 757.
[29] Guvenc S, Oran S, Ozturk S. The epiphytic lichens on Anatolian black pine [PinusnigraArnd. subsp.pallasiana(Lamb.) Holmboe] in Mt. Uludag (Bursa-Turkey). Journal of Applied Biological Sciences, 2009, 3(2): 157- 161.
[30] 欧光龙, 彭明春, 和兆荣, 王崇云, 王浩波, 向伶. 高黎贡山北段植物群落TWINSPAN数量分类研究. 云南植物研究, 2008, 30(6): 679- 687.
[31] 余敏, 周志勇, 康峰峰, 欧阳帅, 米湘成, 孙建新. 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释. 植物生态学报, 2013, 37(5): 373- 383.
[32] 赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990: 147- 192.
[33] 阳含熙, 卢泽愚. 植物生态学的数量分类方法. 北京: 科学出版社, 1981: 1- 420.
[34] 郭水良, 于晶, 陈国奇. 生态学数据分析——方法、程序与软件. 北京: 科学出版社, 2015: 1- 272.
[35] Armstrong R A. The response of lichen growth to transplantation to rock surfaces of different aspect. New Phytologist, 1977, 78(2): 473- 478.
[36] Bjelland T. The influence of environmental factors on the spatial distribution of saxicolous lichens in a Norwegian coastal community. Journal of Vegetation Science, 2003, 14(4): 525- 534.
[37] Scarciglia F, Saporito N, La Russa M F, Le Pera E, Macchione M, Puntillo D, Crisci G M, Pezzino A. Role of lichens in weathering of granodiorite in the Sila uplands (Calabria, southern Italy). Sedimentary Geology, 2012, 280(12): 119- 134.
[38] Allen A, Hilton B. Distribution and zonation of maritime lichens in Sark. La Société Guernesiaise, 1987, 22(2): 234- 257.
[39] MacDonald A M, Lundholm J T, Clayden S R. Saxicolous lichens on a Nova Scotian Coastal Barren. Northeastern Naturalist, 2011, 18(4): 475- 488.
[40] Chen J, Blume H P, Beyer L. Weathering of rocks induced by lichen colonization—a review. CATENA, 2000, 39(2): 121- 146.
[41] Johansson P. Consequences of disturbance on epiphytic lichens in boreal and near boreal forests. Biological Conservation, 2008, 141(8): 1933- 1944.
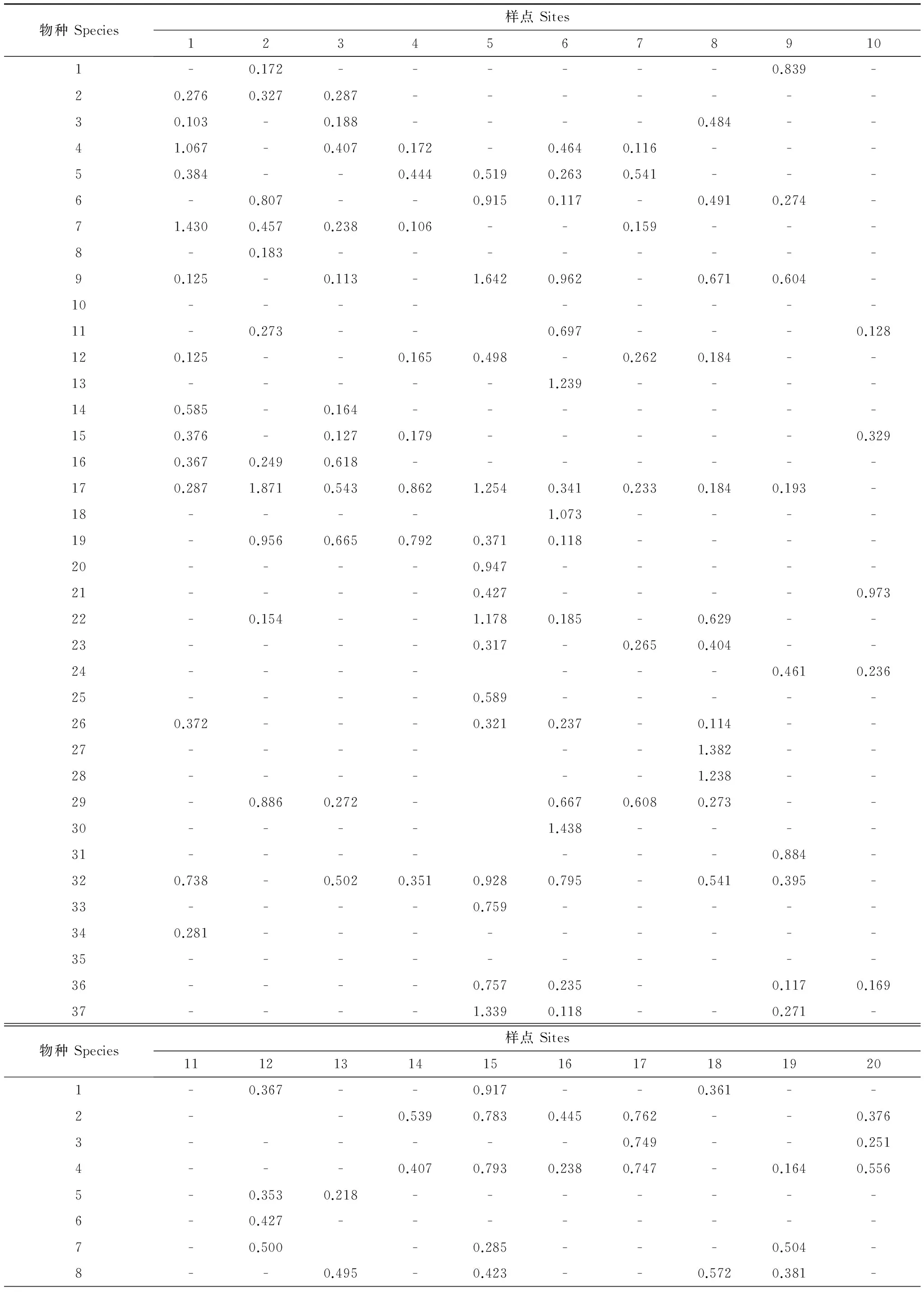
附录Ⅰ 博格达山37种岩面生地衣在20个样点中的相对盖度/%
