川西北草地沙化对土壤可溶性有机氮及酶活性的影响
2018-03-20舒向阳胡玉福杨雨山佘淑凤贾安都姜正博孙锦杨
舒向阳,胡玉福,杨雨山,何 佳,佘淑凤,贾安都,姜正博,孙锦杨
(1.四川农业大学资源学院, 四川 成都 611130; 2.四川农业大学管理学院, 四川 成都 611130)
氮素是限制植物生长发育的主要营养元素之一,土壤氮素是植物氮素营养的主要来源[1-2]。以往研究认为植物氮素营养主要来源于土壤无机氮,但现有研究表明土壤可溶性有机氮(dissolved organic nitrogen, DON)在植物氮素供给中占据着重要地位[3-5]。土壤酶是具有高度专一催化作用的蛋白质,其直接参与土壤物理能量转化的重要生物化学过程,其在土壤养分矿化分解中扮演着重要角色,同时土壤酶活性也常作为评价土壤质量的重要生物指标[6]。研究表明,土壤脲酶、蛋白酶、硝酸还原酶及精氨酸脱氨酶与土壤中氮转化有着密切关系[7]。同时,硝态氮、铵态氮、微生物量氮及可溶性氮是植物氮素的主要来源,因此研究不同沙化程度土壤氮素及酶活性变化有助于深入了解土壤氮素的供应转化。
沙漠化引起的土壤生态环境破坏对全球干旱、半干旱及半湿润地区社会与环境构成了严重威胁。研究表明,沙漠化会加速地表植被多样性损失,同时会破坏土壤结构、降低土壤肥力,破坏土壤微生物生存环境[8-9]。土壤氮素是植物生长过程中的主要养分来源,其含量会直接决定植物氮素的吸收状况,研究表明,沙化过程不仅会改变土壤氮储量,同时也会影响植物氮素含量[10]。川西北高寒草地位于青藏高原东南缘半湿润地区,是我国长江、黄河两大水系的重要水源涵养地,也是全国五大牧区之一,同时也是全球最大的高原泥炭沼泽湿地,其对维护生态环境安全具有重要意义[11-12]。20世纪70年代以来,由于全球气候变化及人为因素的影响,川西北高寒草地面临严重的退化,草地沙化已严重威胁到该区域的社会经济发展,其生态环境屏障作用正在逐步弱化。统计数据表明,截止2009年川西北草地沙化面积达82.19万hm2,预计2020年川西北草地沙化面积将达到95.38万hm2[13]。本文以川西北高寒沙化草地为研究对象,通过对不同沙化程度高寒草地不同土层土壤可溶性有机氮及酶活性的研究,以期揭示高寒草地沙化土壤氮素及酶活性变化特征,为深入理解高寒草地沙化土壤氮素变化及今后沙地修复工作提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区位于四川阿坝藏族羌族自治州红原县境内,地理坐标N31°51′—33°19′,E101°51′—103°23′,境域分属长江、黄河两大水系。地势由东南向西北倾斜,海拔3 210~4 857 m,气候属大陆性高原寒温带季风气候,春秋短促、长冬无夏。年均降雨量791.95 mm,降雨主要集中在5—10月,年均气温为1.1℃,最冷月平均气温-10.3℃,最热月平均气温10.9℃,极端最低气温-36℃,年均积雪期为76 d,无绝对的无霜期。日照充分,太阳辐射强,年均日照时间2 158.7 h,太阳辐射年总量为6 194 MJ·m-2。土地利用现状以草地为主,也有较大面积的沼泽地和沙化地分布,其中沙化土地总面积约为6 915.4 hm2,主要分布邛溪镇和瓦切乡境内。土壤类型以亚高山草甸土为主,高山寒漠土、沼泽化草甸土、岩成土和风沙土等也均匀分布。植被以沙生薹草(Carexpraeclara),垂穗披碱草(Elymussibiricus),华扁穗草(Blysmussinocompressus),线叶蒿草(Kobresiacapillifolia),赖草(Leymussecalinus),淡黄香青(Anaphalisflavescens),黑穗薹草(Carexatrata),木里薹草(Carexmuliensis),细叶亚菊(Ajaniatenuifolia)等为主,植被组合以亚高山草甸为主,沼泽草甸与沼泽植被较为旺盛,植物群落外貌鲜艳,富有季相变化。
1.2 土壤样品采集
本研究于2014年在红原县沙化草地分布较多且集中的瓦切乡选择采样点。本文选择不同沙化程度的草地为研究样地,分别为:未沙化草地(NDG, non-desertification grassland)、轻度沙化草地(LDG, light-desertification grassland)、中度沙化草地(MDG, medium-desertification grassland)和严重沙化草地(HDG, heavy-desertification grassland),详见表1。4种沙化类型草地均选择3处地形和土壤母质一致的样地作为重复,各样地内选取1个面积大小为1 m×1 m的样方用于植被信息调查和土壤样品采集,在选定样方内分别采集0~20 cm、20~40 cm和40~60 cm土壤样品,去除杂物及植物根系、凋落物等。土壤样品分为两份,一份土样冷藏于4℃冰箱,用于测定微生物数量及微生物量碳、氮,另一份土样于室内风干后,保存于密封袋内,用于硝态氮和铵态氮的测定。
1.3 测定方法
土壤脲酶根据Tabatabai方法测定[14];土壤蛋白酶采用福林比色法[15];土壤硝酸还原酶活性采用Kandeler比色法测定[16];土壤精氨酸脱氨酶采用Kandeler比色法测定[16];土壤可溶性有机氮为溶解性全氮与无机氮的差值。
1.4 数据处理
采用Excel 2013进行数据预处理与图表绘制,采用SPSS19.0软件进行单因素方差分析(One-way ANOVA)及Pearson线性相关系数分析指标相关性。
2 结果与分析
2.1 不同沙化程度草地土壤脲酶变化特征
研究结果表明,不同沙化程度草地土壤脲酶存在显著差异(P<0.05)(表2)。其中,相较于未沙化草地,轻度沙化、中度沙化和重度沙化草地土壤脲酶活性分别降低了57.14%、64.28%和73.21%。在0~20 cm土层,土壤脲酶下降程度尤为明显,相较于未沙化草地,轻度、中度和重度沙化草地土壤脲酶分别下降了13.51%、29.73%和40.54%。在土层剖面上,随土层深度增加,土壤脲酶活性不断降低。
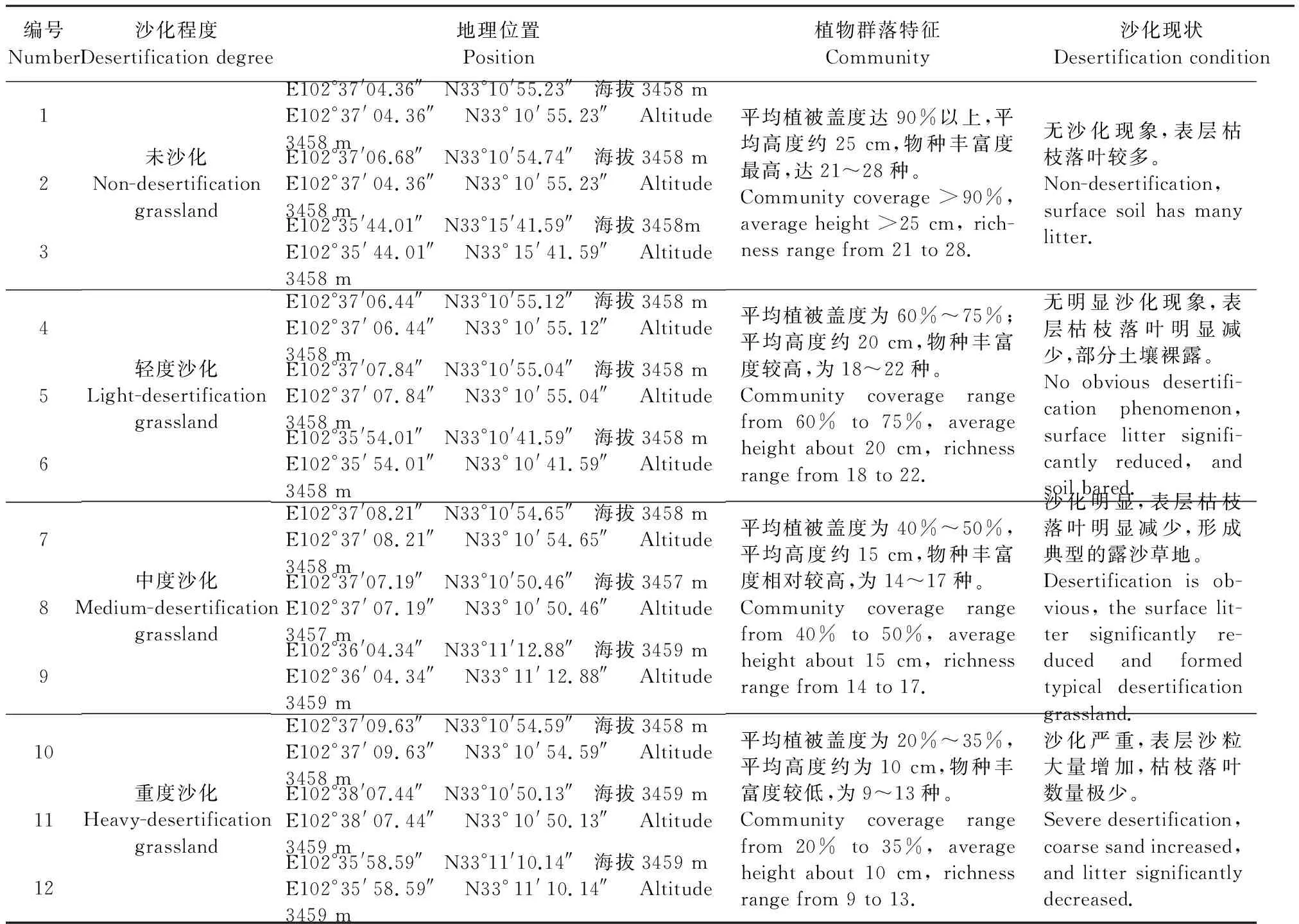
表1 草地样方信息 Table 1 Sampling of desertification grassland quadrat

表2 不同程度沙化草地土壤脲酶活性/(g·kg-1·2h-1) Table 2 Activities of soil urease in different degree of desertifiation grasslands
注:不同小写字母代表不同处理间差异显著水平达到P<0.05。
Note: different lowercases letters indicate significant differences at theP<0.05 level.
2.2 不同沙化程度草地土壤蛋白酶变化特征
由表3可知, 草地沙化导致0~60 cm土壤蛋白酶呈现大幅下降的趋势, 不同沙化草地土壤蛋白酶活性差异显著(P<0.05), 其中, 未沙化草地蛋白酶活性最高可达0.88 mg·kg-1·h-1;相较于未沙化草地,轻度沙化、中度沙化和重度沙化草地土壤蛋白酶分别下降了5.19%、19.48%和33.77%。在0~20 cm土层, 土壤蛋白酶活性下降幅度尤为明显, 随沙化程度增加, 蛋白酶活性呈下降趋势, 相较于未沙化草地, 重度沙化草地降低幅度达30.68%。随土层深度增加, 各沙化草地土壤蛋白酶活性不断降低。
2.3 不同沙化程度草地土壤硝酸还原酶变化特征
由表4可知,相较于未沙化草地,重度沙化0~60 cm土壤硝酸还原酶含量下降了37.11%,随沙化程度增加,土壤硝酸还原酶活性呈显著降低趋势(P<0.05)。其中,在0~20 cm土层,相较于未沙化草地,轻度沙化、中度沙化和重度沙化草地土壤硝酸还原酶活性分别降低了14.29%、27.82%和39.85%。随土层深度增加,土壤硝酸还原酶活性呈不断降低趋势。

表3 不同程度沙化草地土壤蛋白酶活性/(mg·g-1·h-1) Table 3 Activities of soil protease in different degree of desertifiation grasslands
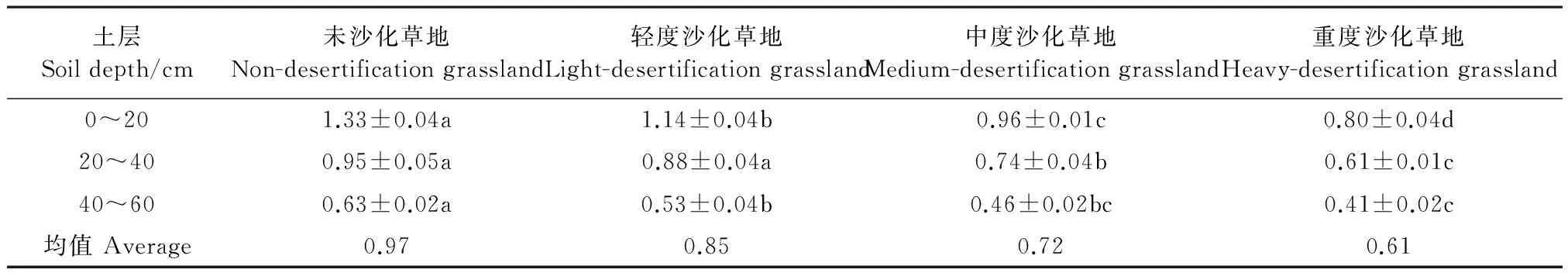
表4 不同程度沙化草地土壤硝酸还原酶活性/(μg·g-1·24h-1) Table 4 Activities of soil nitrate reductase in different degree of desertifiation grasslands
2.4 不同沙化程度草地土壤精氨酸脱氨酶变化特征
由表5可知,草地沙化导致0~60 cm土壤精氨酸脱氨酶呈现大幅下降的趋势,不同沙化草地土壤精氨酸脱氨酶活性差异显著(P<0.05),其中,未沙化草地精氨酸脱氨酶活性最高可达0.25 μg·g-1·3h-1;相较于未沙化草地,轻度沙化、中度沙化和重度沙化草地土壤精氨酸脱氨酶分别下降了15.79%、31.58%和47.37%。在0~20 cm土层,土壤精氨酸脱氨酶活性下降幅度尤为明显,随沙化程度增加,精氨酸脱氨酶活性呈下降趋势,相较于未沙化草地,重度沙化草地降低幅度达44.00%。随土层深度增加,各沙化草地土壤精氨酸脱氨酶活性不断降低。
2.5 不同沙化程度草地土壤可溶性有机氮变化特征
由表6可知,草地沙化导致0~60 cm土壤可溶性有机氮呈现大幅下降的趋势,不同沙化草地土壤可溶性有机氮含量差异显著(P<0.05),其中,未沙化草地可溶性有机氮含量最高可达20.6 mg·kg-1;相较于未沙化草地,轻度沙化、中度沙化和重度沙化草地土壤可溶性有机氮含量分别下降了22.36%、48.72%和67.77%。在0~20 cm土层,土壤可溶性有机氮含量下降幅度尤为明显,随沙化程度增加,可溶性有机氮呈下降趋势,相较于未沙化草地,重度沙化草地降低幅度达64.76%。随土层深度增加,各沙化草地土壤可溶性有机氮含量不断降低。

表5 不同程度沙化草地土壤精氨酸脱氨酶活性/(μg·g-1·3h-1) Table 5 Activities of soil arginine deaminase in different degree of desertifiation grasslands

表6 不同程度沙化草地土壤可溶性有机氮含量/(mg·kg-1) Table 6 Contents of soil dissolved organic nitrogen in different degree of desertifiation grasslands
2.6 不同沙化程度草地土壤微生物量氮、铵态氮及硝态氮变化特征
由图1可知,草地沙化导致0~60 cm土壤微生物量氮、铵态氮及硝态氮呈现大幅下降的趋势,不同沙化草地土壤可溶性有机氮含量差异显著(P<0.05)。在0~20 cm土层,土壤微生物量氮、铵态氮及硝态氮含量下降幅度尤为明显。
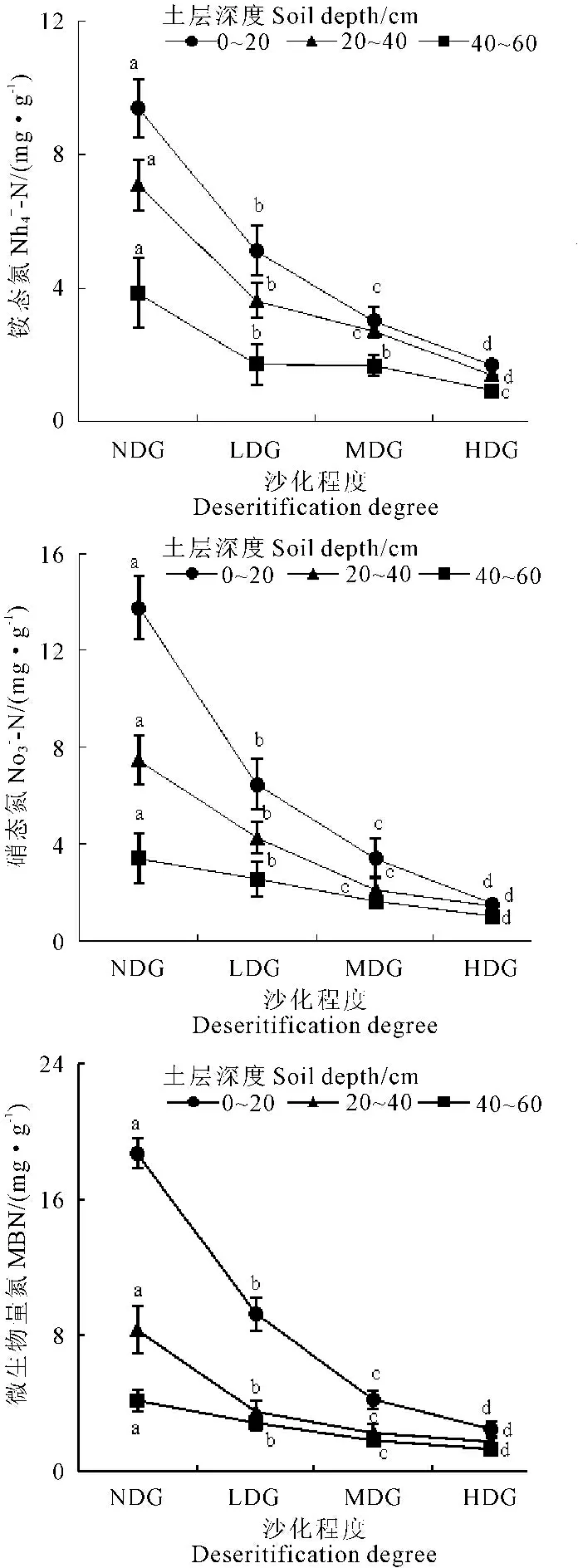
注 Note: NDG—未沙化 non-desertification; LDG—轻度沙化 light-desertification; MDG—中度沙化 medium-desertification; HDG—严重沙化 heavy-desertification.
图1 不同程度沙化草地土壤微生物量氮、铵态氮及硝态氮含量
Fig.1 Contents of soil microbial biomass nitrogen(MBN), NH4+-N and NO3--N in different degree of desertifiation grasslands
2.7 土壤氮素与酶活性相关性分析
相关分析结果表明,蛋白酶、脲酶、硝酸还原酶和精氨酸脱氨酶与铵态氮、硝态氮、微生物量氮和可溶性有机氮均呈极显著正相关(P<0.05)(表7)。其中,硝酸还原酶与铵态氮、硝态氮、微生物量氮和可溶性有机氮的相关系数最高,分别达到0.868、0.850、0.789和0.701,铵态氮与蛋白酶、脲酶、硝酸还原酶、精氨酸脱氨酶相关系数分别达0.737、0.841、0.868和0.838,表明土壤氮素与酶活性密切相关。
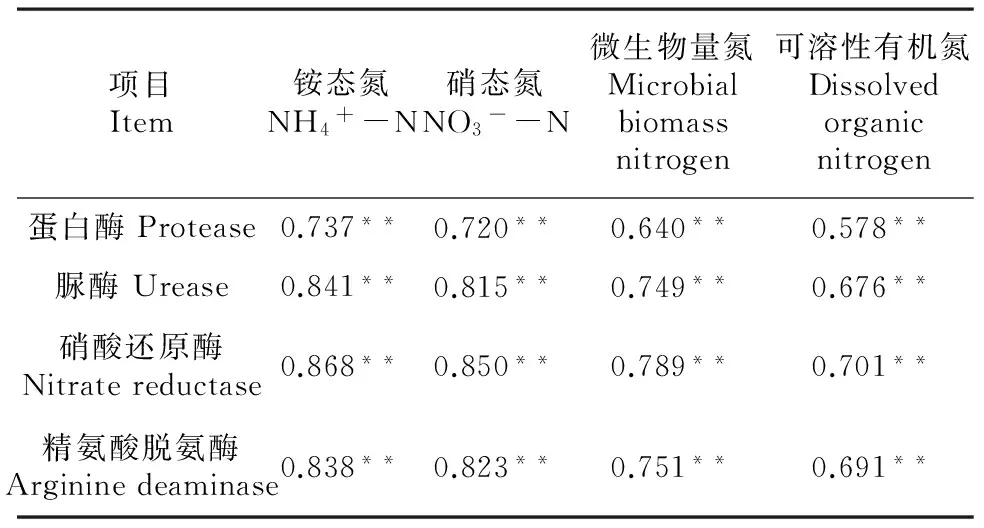
表7 土壤氮素与酶活性相关分析 Table 7 Correlation analysis of soil nitrogen and enzyme activities
注:**在0.01水平(双侧)上显著相关。
Note: **, significant correlation atP<0.01.
3 讨 论
研究结果表明,草地沙化会加速地表植被多样性降低和土壤理化性质破坏,引起土壤生产潜力损失,进而破坏草地生态系统平衡[17-20]。尤全刚等[21]研究表明,高寒草甸草地退化不仅会导致草地植被群落特征改变,同时会改变土壤水热条件,如降低土壤持水量、饱和电导率及增加导热率,进而加速地表水热交换。贺凤鹏等[22]研究温带草地退化土壤剖面微生物学特征表明,土壤表层微生物生物量及酶活性均随退化程度加剧而不断降低,在0~10 cm表层差异尤为明显。本文研究结果表明,随着沙化程度加剧,川西北草地地表植被高度、盖度及物种丰富度大幅度下降,同时土壤表层枯枝落叶也大幅下降;在土壤微生物特性方面,草地沙化会导致氮素相关的脲酶、蛋白酶、硝酸还原酶和精氨酸脱氨酶活性显著降低,在0~20 cm土层影响尤为明显。而随土层深度增加,土壤酶活性逐步降低。其原因主要是由于草地沙化过程中地表植被破坏及土壤微生物生存环境日趋恶化[23],而土壤酶主要来源于植被和土壤微生物分泌释放,进而使得土壤酶活性受到影响。
本文研究同时表明,草地沙化将会导致土壤可溶性有机氮含量不断降低。以往研究认为植物生长中主要以吸收土壤无机氮为主,而今诸多研究表明土壤可溶性有机氮也是植物生长吸收的主要氮素来源[24-25],同时研究表明地表植被生物多样性与可溶性有机氮有着密切关系[26-27]。张玉霞等[28]研究表明,草地在不同干扰生境下土壤铵态氮和硝态氮含量变化与植被地上生物量呈显著正相关。王文颖等[29]研究高寒人工草地土壤可溶性有机氮和无机氮结果表明,退化草地中土壤硝态氮和可溶性有机氮占优势,且在高寒区人工草地中土壤可溶性有机氮是植物可利用氮的重要组成部分。
4 结 论
川西北高寒草地退化过程中,草地生态系统中的氮素相关酶活性以及可溶性有机氮会发生相应变化。本文研究表明,土壤脲酶、蛋白酶、硝酸还原酶及精氨酸脱氨酶活性均随沙化加剧而显著降低;土壤可溶性有机氮含量也大幅下降;土壤氮素相关酶与土壤无机氮和溶解性有机氮呈极显著正相关,其中,铵态氮与氮素相关酶活性相关系数最大,硝酸还原酶与土壤氮素相关系数最高。综上可见,在沙化草地修复过程中,应添加氮肥以提高土壤氮素含量。
[1] Mueller K E, Hobbie S E, Tilman D, et al. Effects of plant diversity, N fertilization, and elevated carbon dioxide on grassland soil N cycling in a long-term experiment[J]. Global Change Biology, 2013,19(4):1249-1261.
[2] Ceulemans T, Merckx R, Hens M, et al. Plant species loss from European semi-natural grasslands following nutrient enrichment-is it nitrogen or is it phosphorus?[J]. Global Ecology and Biogeography, 2013,22(1):73-82.
[3] Hasselquist N J, Metcalfe D B, Marshall J D, et al. Seasonality and nitrogen supply modify carbon partitioning in understory vegetation of a boreal coniferous forest[J]. Ecology, 2016,97(3):671-683.
[4] Jiang L, Wang S P, Luo C Y, et al. Effects of warming and grazing on dissolved organic nitrogen in a Tibetan alpine meadow ecosystem[J]. Soil and Tillage Research, 2016,158:156-164.
[5] 林 丽,李以康,崔 莹,等.青藏高原典型植被土壤溶解性氮与植物量相关性分析[J].山地学报,2012,30(6):721-727.
[6] Burns R G, DeForest J L, Marxsen J, et al. Soil enzymes in a changing environment: current knowledge and future directions[J]. Soil Biology and Biochemistry, 2013,58(3):216-234.
[7] Garcla-Gil J C, Plaza C, Soler-Rovira P, et al. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass[J]. Soil Biology and Biochemistry, 2000,32(13):1907-1913.
[8] Tang Z S, An H, Shangguan Z P. The impact of desertification on carbon and nitrogen storage in the desert steppe ecosystem[J]. Ecological Engineering, 2015,84(11):92-99.
[9] Allington G R H, Valone T J. Reversal of desertification: the role of physical and chemical soil properties[J]. Journal of Arid Environments, 2010,74(8):973-977.
[10] Zhao H L, He Y H, Zhou R L, et al. Effects of desertification on soil organic C and N content in sandy farmland and grassland of Inner Mongolia[J]. Catena, 2009,77(3):187-191.
[11] Xiang S, Guo R Q, Wu N, et al. Current status and future prospects of Zoige Marsh in eastern Qinghai-Tibet Plateau[J]. Ecological Engineering, 2009,35(4):553-562.
[12] Li J C, Wang W L, Hu G Y, et al. Changes in ecosystem service values in Zoige Plateau, China[J]. Agriculture, Ecosystems & Environment, 2010,139(4):766-770.
[14] Tabatabai M A. Soil Enzymes. Methods of Soil Analysis:Part 2-microbiological and Biochemical Properties[M]. Madison, WL, USA: Soil Science Society of America, 1982:775-833.
[15] Ladd J N, Butler J H A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates[J]. Soil Biology and Biochemistry, 1972,4(1):19-30.
[16] Kandeler E. Methods in Soil Biology[C]//Schinner F, Ohlinger R, Kandeler E, et al. Book Methods in Soil Biology. New York: Springer-Verlag, Heidelberg, 1996:168-170.
[17] Li F R, Zhao W Z, Liu J L, et al. Degraded vegetation and wind erosion influence soil carbon, nitrogen and phosphorus accumulation in sandy grasslands[J]. Plant and Soil, 2009,317(1-2):79-92.
[18] Martinez-Mena M, Rogel J A, Castillo V, et al. Organic carbon and nitrogen losses influenced by vegetation removal in a semiarid mediterranean soil[J]. Biogeochemistry, 2002,61(3):309-321.
[19] Zuo X A, Zhao H L, Zhao X Y, et al. Vegetation pattern variation, soil degradation and their relationship along a grassland desertification gradient in Horqin Sandy Land, northern China[J]. Environmental geology, 2009,58(6):1227-1237.
[20] 秦嘉海,张 勇,赵芸晨,等.祁连山黑河上游不同退化草地土壤理化性质及养分和酶活性的变化规律[J].冰川冻土,2014,36(2):335-346.
[21] 尤全刚,薛 娴,彭 飞,等.高寒草甸草地退化对土壤水热性质的影响及其环境效应[J].中国沙漠,2015,(5):1183-1192.
[23] 舒向阳,胡玉福,蒋双龙,等.川西北沙化草地植被群落,土壤有机碳及微生物特征[J].草业学报,2016,25(4):45-54.
[24] Alexandre A, Balesdent J, Cazevieille P, et al. Direct uptake of organic carbon by grass roots and allocation in leaves and phytoliths: 13C labeling evidence[J]. Biogeosciences Discussions, 2015,12(23):19751-19780.
[25] Moran-Zuloaga D, Dippold M, Glaser B, et al. Organic nitrogen uptake by plants: reevaluation by position-specific labeling of amino acids[J]. Biogeochemistry, 2015,125(3):359-374.
[26] Van Kessel C, Clough T, van Groenigen J W. Dissolved organic nitrogen: an overlooked pathway of nitrogen loss from agricultural systems?[J]. Journal of Environmental Quality, 2009,38(2):393-401.
[27] Oelmann Y, Wilcke W, Temperton V M, et al. Soil and plant nitrogen pools as related to plant diversity in an experimental grassland[J]. Soil Science Society of America Journal, 2007,71(3):720-729.
[28] 张玉霞,姚 拓,王国基,等.高寒生态脆弱区不同扰动生境草地植被及土壤无机氮变化特征[J].草业学报,2014,23(4):245-252.
[29] 王文颖,李文全,周华坤,等.高寒人工草地土壤可溶性有机氮库和无机氮库动态变化[J].生态环境学报,2016(1):30-35.
