β-淀粉样蛋白诱导的阿尔茨海默病大鼠模型中BDNF与TRPC3的关系
2018-03-20张利华张丽艳张小康徐超龙周义
张利华,张丽艳,张小康,徐超龙,周义
(沈阳医学院基础医学院病理生理学教研室,沈阳 110034)
阿尔茨海默病 (Alzheimer’s disease,AD) 又称老年痴呆症,是一种以记忆、学习、认知功能改变为主要特征的神经系统退行性疾病。病因假说很多,其中β-淀粉样蛋白 (β-amyloid protein,A β) 异常沉积占主导地位。其主要病理特征是在患者大脑皮质、海马区出现老年斑和神经元纤维缠结,此外还有神经元缺失、神经元突触异常等病理改变。脑源性神经营养因子 (brain-derived neurotrophic factor,BDNF)是中枢神经系统中分布最广泛的营养因子,在突触可塑性、神经元存活方面起着关键作用。最近的研究[1]报道,AD患者的血清中BDNF水平降低,且发现健康老年受试者的脑脊液中BDNF水平低下可能预示着未来会出现认知衰退。哺乳动物中的经典瞬时受体电位通道蛋白 (canonical transient receptor potential channel,TRPC) 是一种非典型钙离子通道,与神经系统的生长、增殖、分化和凋亡密切相关。BDNF通过激活TRPC3促进树突棘的生长发育[2]。卡洛西汀通过促进TRPC6和磷酸化环磷腺苷效应元件结合蛋白的表达从而治疗脑缺血损伤[3]。在癫痫大鼠海马神经元初代培养中,BDNF和TRPC3表达增加对癫痫引起的细胞损伤起保护作用[4]。在视网膜缺血性损伤的研究中发现,BDNF/酪氨酸激酶受体B通过调节TRPC3/6通道的表达进而发挥神经节细胞的保护作用。
本课题组的前期研究[5]发现,在A β诱导的AD小鼠模型中,TRPC4表达增加。但目前关于BDNF和TRPC3在A β所致的AD模型中的关系及作用机制尚不清楚。本研究采用SD大鼠侧脑室注射寡聚型A β 1-42的方法模拟AD[6],利用逆转录聚合酶链反应 (RTPCR) 和Western blotting检测大鼠海马组织中TRPC3和BDNF mRNA和蛋白表达,探讨它们之间的关系,为AD发病机制研究提供新的证据,同时也为预防和治疗AD提供新的思路。
1 材料与方法
1.1 实验材料
1.1.1 动物:24只5周龄200~220 g的SPF级雄性SD大鼠 (辽宁长生生物技术有限公司) 。
1.1.2 试剂:A β和BDNF兔单克隆抗体 (英国Abcam公司) ;重组人源BDNF (美国Peprotech公司) ;总RNA小量制备试剂盒 (美国Axygen公司) ;逆转录试剂盒 (加拿大Abm公司) ;BCA 蛋白浓度测定试剂盒和SDS-PAGE凝胶配制试剂盒 (上海碧云天公司) ;TRPC3兔多克隆抗体 (台湾Arigo公司) ;β-actin小鼠单克隆抗体和辣根酶标记山羊抗兔IgG (H+L)(美国Proteintech公司) ;辣根酶标记山羊抗小鼠IgG(H+L) (北京中杉金桥公司) 。
1.2 方法
1.2.1 A β 1-42寡聚化处理:注射前,将A β 1-42室温放置1 h,低速离心10 min,溶解在灭菌PBS溶液中 (浓度为1 μ g/μ L) ,并在37 ℃恒温箱中孵育48 h诱导寡聚化,放置冰上备用。
1.2.2 AD模型制备:大鼠经腹腔注射10%水合氯醛(0.4 mL/100 g) 麻醉,头部消毒后将皮肤沿矢状线切开 1 cm,棉签蘸碘伏消毒,剔除骨膜,完全暴露颅骨前囟,置于脑立体定位仪上,侧脑室导管放置位置根据《大鼠脑立体定位图谱》确定,前囟后 0.8 mm,中线向右旁开 1.5 mm,深度3.5 mm。置管后在管周前、左、右3个位置分别拧入小螺丝半固定,涂以牙科水泥固定,晾干后缝合。肌注青霉素5 d预防感染。待手术应激消失后,用微量注射器经导管缓慢注射A β 1-42 10 μ L,注射速度约为2 μ L/min,注射结束留针3 min,使溶液充分弥散。对照组注射同等剂量的PBS溶液。
1.2.3 BDNF给药:根据说明书溶解和稀释BDNF,分装后置于-40 ℃冰箱低温保存,避免反复冻融。A β 1-42给药14 d后即水迷宫实验前1 d,从第15天起,给予AD+BDNF组大鼠BDNF水溶液,剂量为10 μ L(0.025 μ g/μ L) ,持续给药5 d。
1.2.4 Morris水迷宫行为学检测:A β 1-42给药后第16天,利用MT-200 Morris 水迷宫跟踪分析系统 (成都泰盟科技有限公司) 检测各组大鼠学习和记忆能力。前5 d进行定位航行实验,将待测大鼠头部面向池壁放入水中,让其寻找目标象限内的隐藏平台,所用时间即逃避潜伏期,依据该时间的长短检测大鼠的空间学习能力,3次/d,平台位置不动,象限不重复。第6天进行空间探索实验,将平台撤去,记录大鼠穿越平台次数以及在平台所在象限停留时间,以检测大鼠的空间记忆能力。
1.2.5 RT-PCR 检测TRPC3和BDNF mRNA的表达:Morris水迷宫实验结束后将大鼠断头取海马组织,-80 ℃超低温冰箱保存,以备RT-PCR实验以及Western blotting实验使用。提取大鼠海马组织总RNA后进行逆转录。以逆转录所得的cDNA为模板进行PCR扩增,所需引物由金斯瑞生物科技公司合成。β-actin上游引物5’-AGCGGGAAATCGTGCGTG-3’,下游引物5’-CAGGGTACATGGTGGTGCC-3’;BDNF上游引物5’-CTGAGCGTGTGTGACAGTATTAGCG-3’,下游引物5’-CAGCCTTCCTTCGTGTAACCCAT-3’;TRPC3,上游引物5’-CTGGCCAACATAGAGAAGGAGT-3’,下游引物5’-CACCGATTCCAGATCTCCAT-3’。琼脂糖凝胶成像结果采用Gel Image System软件进行分析。
1.2.6 Western blotting 检测 TRPC3 和BDNF蛋白的表达:向低温保存的海马组织中加入含有蛋白酶抑制剂的RIPA裂解液200 μ L,12 000 r/min离心5 min。取上清液,采用 BCA 法得出标准曲线,根据样品吸光度计算得出蛋白浓度。蛋白上样量为30 μ g/10 μ L,室温下SDS-PAGE凝胶电泳1 h,TRPC3 (分子量91 000) 采用湿转方式转膜2 h,BDNF (分子量15 000) 采用半干转方式转膜20 min,放入5%脱脂牛奶中摇床封闭1 h,加入适度稀释度的一抗TRPC3 (1∶2 000) 、BDNF (1∶2 000)、 β-actin (1∶5 000) 4 ℃孵育过夜。翌日TBST漂洗15 min×3次,加入适度稀释的二抗(1∶2 000) ,冰上孵育2 h,TBST漂洗15 min×3次,最后,用化学发光成像系统 (中国天能公司) 进行ECL显影,用Gel Image System 软件对实验结果进行分析。
1.3 统计学分析
2 结果
2.1 BDNF对AD大鼠空间学习记忆能力有改善作用
侧脑室导管A β 1-42给药14 d后,从第15天开始,BDNF连续给药5 d,期间进行水迷宫实验。第5天大鼠逃避潜伏期AD组较PBS组延长,有统计学差 异[分 别 为(31.06±4.01) s和(24.23±4.62) s,P < 0.05];第6天穿越平台次数AD组较PBS组减少(分 别 为3.25±0.45和6.62±0.46,P < 0.05) ,表 明AD组大鼠的空间学习记忆能力下降。而第5天大鼠逃避潜伏期AD+BDNF组较AD组缩短[分别为(23.30±4.84) s和(31.06±4.01) s,P < 0.05]; 第6天穿越平台次数AD+BDNF组较AD组增加 (分别为5.83±0.75和3.25±0.45,P < 0.05) ,表明BDNF对AD大鼠空间学习记忆能力有改善作用。见图1。
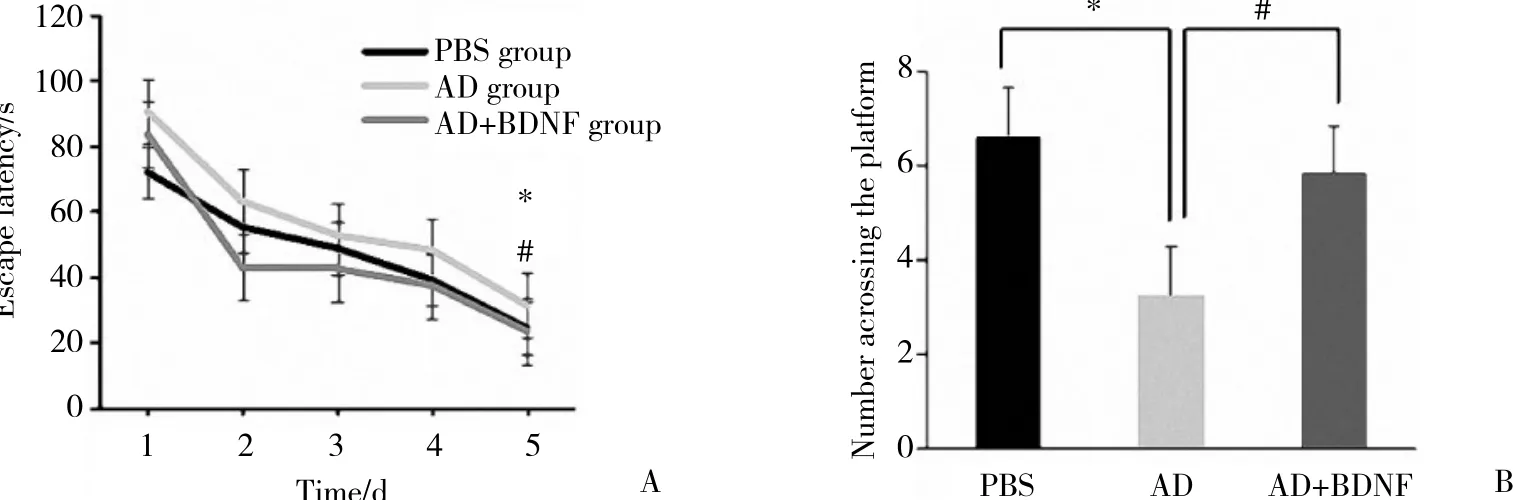
图1 Morris 水迷宫实验中各组大鼠行为学比较Fig.1 Behavioral comparisons of each group of rats in the Morris water maze
2.2 BDNF促进TRPC3 mRNA的表达
RT-PCR结果显示,AD组与PBS组相比,BDNF和TRPC3 mRNA相对含量均降低 (P < 0.05) 。AD+BDNF组与AD组相比,TRPC3 mRNA相对含量增加 (P <0.05) 。见图2。
2.3 BDNF促进TRPC3 蛋白的表达
Western blotting结果显示,AD组与PBS组相比,BDNF和TRPC3 蛋白相对含量均降低 (P < 0.05) ,AD+BDNF组与AD组相比,TRPC3 蛋白相对含量增加 (P < 0.05) 。见图3。
3 讨论
研究[7]表明,A β在脑内沉积形成老年斑是AD最具特征的病理改变,与大脑认知障碍密切相关。A β在AD的发病机制中起着核心作用,但它通过何种机制造成神经元损伤,尚不清楚。在转基因AD小鼠脑组织中观察到BDNF的降低可能会促进神经元萎缩和突触损伤,而BDNF的上调可以控制AD的认知衰退[8]。本研究中,AD组大鼠BDNF表达下降,空间学习记忆能力减退;当给予AD大鼠一定剂量的BDNF时,可明显改善其空间学习记忆能力,与之前ZHANG等[9]的研究结果一致。
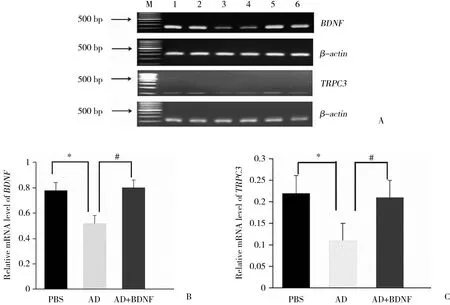
图2 RT-PCR检测大鼠海马区BDNF和TRPC3 mRNA的表达水平Fig.2 Expression of BDNF and TRPC3 mRNA in the hippocampus of rats detected by RT-PCR
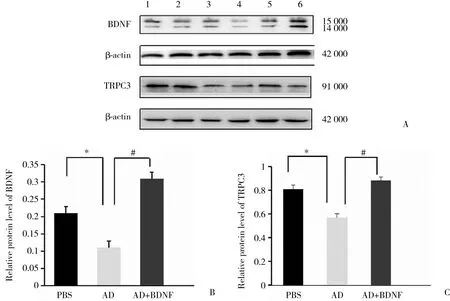
图3 Western blotting检测各组大鼠海马区BDNF 和TRPC3蛋白表达Fig.3 Expression of BDNF and TRPC3 protein in the hippocampus detected by Western blotting
有研究[10]表明,在心肌缺血性损伤及癫痫大鼠模型中,BDNF与TRPC3/6协同作用,对神经元损伤起保护作用。本研究中,AD组大鼠TRPC3表达下降,当给予AD大鼠一定剂量的BDNF时,TRPC3 mRNA和蛋白水平均明显增加,说明BDNF可能通过上调TRPC3从而发挥其改善AD大鼠认知障碍的作用,这在AD模型中尚属首次发现。
有研究[11]表明,BDNF通过钙调蛋白依赖性蛋白激酶激酶激活TRPC5或TRPC6通道从而诱导谷氨酸亚基1翻译和突触生长。也有研究[12]表明,哌嗪类化合物通过激活DAG,进而引起TRPC3/6/7介导的Ca2+内流,发挥BDNF样神经营养作用。但在AD模型中,BDNF通过何种途径调控TRPC3的表达,仍需进一步的试验证实。
综上所述,本研究表明,BDNF可能通过上调TRPC3表达,从而发挥其对AD大鼠认知障碍的改善作用,但其具体机制有待研究。这可能对临床预防和治疗AD有一定帮助。
[1] DINIZ BS,TEIXEIRA AL. Brain-derived neurotrophic factor and Alzheimer’s disease:physiopathology and beyond [J]. Neuromolecular Med,2011,13 (4) :217-222. DOI:10.1007/s12017-011-8154-x.
[2] AMARAL M D,POZZOMILLER L. TRPC3 channels are necessary for brain-derived neurotrophic factor to activate a nonselective cationic current and to induce dendritic spine formation [J]. Neurosci,2007,27( 19) :5179-5189. DOI:10.1523/JNEUROSCI.5499-06.2007.
[3] CHAO G,MA Y,MA S,et al. The role of TRPC6 in the neuroprotection of calycosin against cerebral ischemic injury[ J]. Sci Rep,2017,7(1) :3039. DOI:10.1038/s41598-017-03404-6.
[4] YANG ZW,WU F,ZHANG SL. Effects of ganoderic acids on epileptiform discharge hippocampal neurons:insights from alterations of BDNF,TRPC3 and apoptosis[ J]. Pharmazie,2016,71( 6) :340-344.DOI:10.1691/ph.2016.5889.
[5] 张改改,张丽艳,邹丹,等. 经典瞬时受体电位通道蛋白在β淀粉样蛋白诱导的阿尔茨海默病小鼠海马区的表达[J]. 中国医科大学学报,2016,45( 12) :1100-1104. DOI:10.12007/j.issn.0258-4646.2016.12.010.
[6] 于方,拓西平,吕建勇. 侧脑室注射β-淀粉样蛋白制作阿尔茨海默病样大鼠模型[J]. 实用老年医学,2010,24( 6) :475-478. DOI:10.3969/j.issn.1003-9198.2010.06.011.
[7] WEI Z,CHEN XC,SONG Y,ET AL. Amyloid β protein aggravates neuronal senescence and cognitive deficits in 5XFAD mouse model of Alzheimer’s disease[ J]. Chin Med J ( Engl) ,2016,129( 15) :1835-1844. DOI:10.4103/0366-6999.186646.
[8] NAGAHARA AH,MATELING M,KOVACS I,et al. Early BDNF treatment ameliorates cell loss in the entorhinal cortex of APP transgenic mice[ J]. Neurosci,2013,33( 39) :15596-15602. DOI:10.1523/JNEUROSCI.5195-12.2013.
[9] ZHANG L,FANG Y,LIAN Y,et al. Brain-derived neurotrophic factor ameliorates learning deficits in a rat model of Alzheimer’s disease induced by A β 1-42[ J]. PLoS One,2015,10( 4) :e0122415. DOI:10.1371/journal.pone.0122415.
[10] HANG P,ZHAO J,CAI B,ET al. Brain-derived neurotrophic factor regulates TRPC3/6 channels and protects against myocardial infarction in rodents[ J]. Int J Biol Sci,2015,11( 5) :536-545. DOI:10.7150/ijbs.10754.
[11] FORTIN DA,SRIVASTAVA T,DWARAKANATH D,et al. BDNF activation of CaM-kinase kinase via TRPC channels induces the translation and synaptic incorporation of GluA1 containing calcium-permeable AMPARs[ J]. Neurosci,2012,32( 24) :8127-8137.DOI:10.1523/JNEUROSCI.6034-11.2012.
[12] SAWAMURA S,HATANO M,TAKADA Y,et al. Screening of transient receptor potential canonical channel activators identifies novel neurotrophic piperazine compounds[ J]. Mol Pharmacol,2016,89( 3) :348-363. DOI:10.1124/mol.115.102863.
