表达IP10基因重组新城疫病毒的拯救
2018-03-15郭顺利谢美玉宾晓芸石明霞陈安宁陈思宇周素芳
郭顺利,谢美玉,宾晓芸,石明霞,陈安宁,陈思宇,周素芳
(广西医科大学,南宁530021)
近年溶瘤病毒能够增强肿瘤相关抗原的表达,引起肿瘤特异性免疫反应[1]。新城疫病毒(NDV)作为溶瘤病毒家族中的一员也逐渐被人们认识[2]。NDV为副黏病毒科、腮腺炎病毒属的单股负链RNA病毒,核酸长度为15 186 bp,核酸序列为3'-NP-P-M-F-HN-L-5',每个基因都有独立的基因起始区(GS)、基因终止区(GE)和基因间序列[3]。根据对鸟类致病力不同可将病毒分为强毒株、中强毒株、弱毒株。NDV的致病性通常被认为由F基因所决定。F基因产生的F蛋白以F0融合蛋白形式存在,只有经胰蛋白酶作用后才具有重新感染细胞的能力[4]。NDV感染人体后,不依赖于人体细胞分裂,能够独立复制,且不会将基因组整合到人的基因组中。另外,NDV能够诱导肿瘤细胞凋亡,而对人体正常细胞无明显不良反应[5]。虽然中强毒株对肿瘤的杀伤作用更强,但如保存和利用的不恰当,会给鸟类和家禽业带来重大损失;并且中强毒株对正常细胞的杀伤作用也远高于弱毒株。因此,弱毒株组Lasota是溶瘤的最好选择。干扰素γ诱导蛋白10(IP10/CXCL10)是细胞因子,为CXC趋化因子家族的一员。通过与CXCR3配体结合,能够趋化自然杀伤(NK)细胞,活化CD4+T细胞和CD8+T细胞等[6]。并且,IP10具有抑制血管生成的作用,它能够独自[7]或与其他药物共同作用[8]治疗肿瘤。随着反向遗传学技术的迅速发展,为NDV与免疫治疗相结合提供了可能。2016年6月~2017年1月,本研究将具有肿瘤选择性的NDV Lasota株和具有趋化免疫细胞的IP10进行基因序列重组,利用反向遗传学技术拯救重组病毒rNDV-IP10;且验证重组病毒的生长动力学和IP10基因的表达,为以后的研究奠定基础。
1 材料与方法
1.1 细胞、质粒及试剂 能稳定表达T7 RNA聚合酶的BSR-7细胞、原代鸡胚成纤维细胞(CEF细胞)和乳仓鼠肾细胞(BHK-21细胞)均用含10% FBS的DMEM培养,生长环境为37 ℃、5% CO2。用弱毒株Lasota作为重组病毒的载体。重组NDV基因组转录载体pBRN-FL-PmeⅠ及辅助质粒pBS-NP、pBS-P、pBS-L、细胞均由哈尔滨兽医研究所提供。人肝癌细胞低转移株(MHCC-97L细胞)为本实验室保存,使用含10%FBS的高糖DMEM培养。IP10的编码基因序列由长沙优宝生物科技有限公司合成,由pMD18-T载体携带。SPF鸡胚由哈尔滨兽医研究所实验动物中心提供。重组酶和Taq酶均购自诺唯赞公司。PmeⅠ限制性内切酶购自NEB公司。磷酸钙转染试剂盒购自Invitrogen公司。小鼠抗人Ip10单克隆抗体购自Abcam公司。小鼠抗人β-actin抗体购自武汉三鹰。FITC标记山羊抗鼠抗体购自sigma公司。携带IgG-HRP标记的山羊抗小鼠抗体购自北京全式金公司。ECL超敏化学发光检测试剂盒购自索莱宝公司。荧光显微镜购自奥林巴斯公司。
1.2 携带IP10基因质粒的构建 用限制性内切酶切开pBRN-FL-PmeⅠ载体中P和M基因间的PmeⅠ位点。使用IP10上游引物(5′-AGGTCCAACTCTGTTTAAACTTAGAAAAAATACGGGTAGAAGTGCC-ACCATGAATCAAACTGCCATTCTG-3′)和下游引物(5′-ATTGCCGCTTGGGTTTAAACTTAAGGAGATCTT-TTAGACCTTTCC-3′)扩增出携带GN和GE的IP10基因序列。用琼脂糖凝胶DNA回收试剂盒纯化目的基因片段。将纯化的产物用重组酶克隆入pBRN-FL-PmeⅠ载体中,构建出携带IP10基因的质粒。对构建的质粒进行测序,挑选出正确质粒pBRN-FL-IP10。
1.3 重组病毒的拯救 将BSR-7细胞铺于六孔板中,细胞长满六孔板约80%,将pBRN-FL-IP10质粒5 μg、pBS-NP质粒2.5 μg、pBS-P质粒1.25 μg、pBS-L质粒1.25 μg共转染BSR-7细胞。12 h后用10%的DMSO休克细胞2.5 min,加入完全培养基,24 h后换为opti-MEM并加入1 μg/mL甲苯磺酰苯丙氨酰氯甲酮。将细胞放入培养箱继续培养。3 d后刮取细胞,用0.25 μm过滤器过滤混合液,滤液注入9~11日龄SPF鸡胚尿囊腔中(400 μL/枚)。注射后的鸡胚在孵化箱中继续孵育5 d。随后抽取鸡胚尿囊液50 μL,用1%浓度的鸡红细胞测病毒效价。收取效价阳性鸡胚尿囊液,于-80 ℃保存。
1.4 重组病毒生长动力学检测 将NDV和rNDV-IP10以感染复数(MOI)=0.01分别感染24孔板中的CEF细胞1 h,去除病毒稀释液,随后加入含10% FBS的DMEM继续培养。隔24 h收集细胞培养液,将收集的培养液再次感染CEF细胞。病毒滴度使用终点滴定检测和50%组织细胞感染量计算。
1.5 重组病毒上外源基因IP10表达检测 采用间接免疫荧光技术。分别用NDV、rNDV-IP10以MOI=0.000 1感染BHK-21细胞1 h,去除病毒稀释液换成含10% FBS培养基继续培养24 h。弃培养液,用1×PBST洗2次;4%的多聚甲醛室温固定15 min。弃固定液,用1×PBST洗3次,每次5 min。用含1%的BSA溶液室温封闭1 h。小鼠抗人IP10抗体室温孵育1 h;弃抗体稀释液,用1×PBST洗3次,每次5 min;用FITC标记的山羊抗鼠抗体避光孵育30 min。1×PBST洗洗3次,每次5 min。用显微镜观察荧光。
1.6 MHCC-97L细胞内IP10蛋白表达检测 采用Western blotting法。分别用NDV、rNDV-IP10以MOI=0.1感染MHCC-97L细胞1 h,随后加入含10% FBS的高糖DMEM继续培养,36 h后收获细胞。将样品用12% SDS-PAGE分离,然后将凝胶中的样品通过湿转法90 mA、25 min转移至PVDF膜上。5%脱脂牛奶室温封闭1 h。小鼠抗人IP10特异性抗体和小鼠抗人β-actin特异性抗体4 ℃孵育过夜。辣根过氧化物酶标记山羊抗小鼠抗体孵育1 h。ECL发光液浸泡后进行成像分析。
2 结果
2.1 重组病毒rNDV-IP10的产生 IP10基因序列插入重组病毒质粒的P和M基因间,将重组质粒命名为pBRN-FL-IP10。重组质粒pBRN-FL-IP10和pBS-NP、pBS-P和pBS-L共转染BSR-7细胞,拯救出重组病毒rNDV-IP10。收集培养液注入9~ 11日龄鸡胚尿囊腔中扩增,5 d后收取尿囊液测得血凝效价滴度约为1∶256,收取阳性尿囊液。
2.2 重组病毒生长特征 用无外源基因的亲本NDV作为对照组。两种病毒生长特征相似,约72 h时滴度最高。插入外源基因的rNDV-IP10在CEF细胞中能够如亲本一样正常生长复制。见图1。
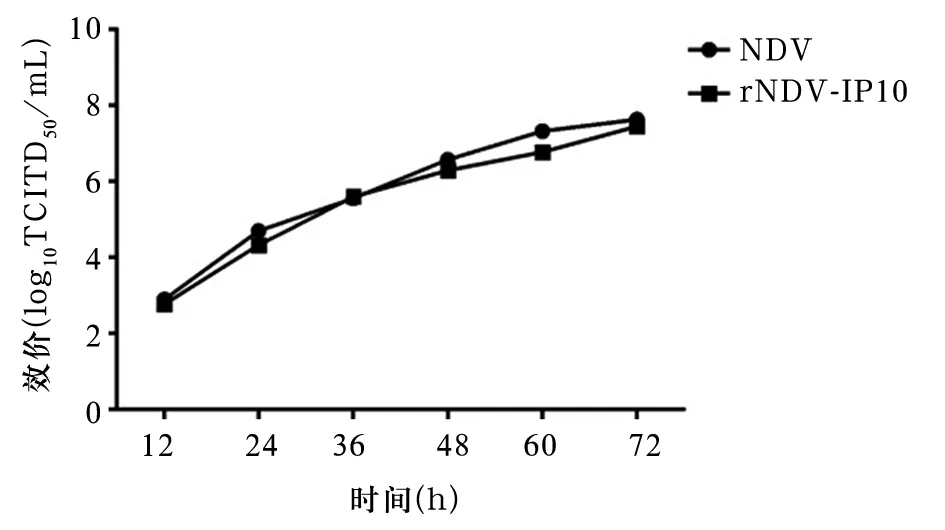
图1 病毒NDV和rNDV-IP10的生长动力曲线
2.3 重组病毒上外源基因IP10表达情况 间接免疫荧光技术证实,感染了亲本NDV的BHK-21细胞未见绿色荧光,而感染了rNDV-IP10病毒的BHK-21细胞可见绿色荧光。由此可鉴,rNDV-IP10病毒能够表达IP10。
2.4 IP10基因在MHCC-97L细胞中的表达 感染了rNDV-IP10的MHCC-97L细胞蛋白样品有特异条带,而感染了NDV的MHCC-97L细胞未出现条带。
3 讨论
NDV对Ⅰ型干扰素较敏感,许多肿瘤细胞缺乏干扰素信号转导通路,因而NDV能在肿瘤细胞中复制而不能在正常细胞中生长[9]。另外,肿瘤细胞也缺乏相应的抗病毒信号通路[10]。数据表明,NDV在肿瘤细胞中的复制能力是正常细胞的10 000倍,并且不具有嗜神经性[11]。临床研究结果表明,即使当人体感染NDV后也仅表现出轻微流感样病理反应,而无其他严重的病理反应[12]。另外,NDV可刺激机体的免疫功能,通过诱导细胞毒性T细胞、NK细胞、单核细胞等来增强机体的抗瘤作用[13]。虽然部分NDV株被用于临床,并取得一定的疗效[14],但是其溶瘤疗效仍需提高。随着反向遗传学技术的成熟,研究者正通过对NDV进行基因改造来增强溶瘤效果。将外源基因插入NDV基因中发现,外源基因的插入不会影响病毒的复制,并且目的基因能够正常表达。更重要的是,重组NDV对肿瘤细胞的选择性杀伤作用改变,并增强了对肿瘤细胞的杀伤作用。重组NDV明显抑制了肿瘤的生长,延长了小鼠的生存时间[15]。由于溶瘤病毒能够打破肿瘤的免疫耐受性,且具有杀伤肿瘤细胞的能力,越来越受人们的关注。
IP10通过与CXCR3配体结合定向趋化CD4+T、CD8+T、NK细胞等抑制肿瘤,并且能够与其他基因或药物联合治疗肿瘤,明显延长了动物生存时间和抑制了肿瘤血管的生成,联合治疗效果明显要好于单独给药[8]。因此,可将IP10的抗瘤作用与NDV的选择性作用相结合治疗肿瘤。
本研究中,通过反向遗传技术将IP10序列插入NDV基因组中。由于IP10基因序列较短,而Lasota载体过大,使用T4连接酶连接效率过低。本研究通过使用重组酶将此连接阳性率增加。重组酶能够简单、高效的将小片段基因连接到大片段载体上,甚至无需对线性化载体及PCR产物进行纯化便有很高的连接效率。通过基因组测序选出正确的重组质粒,最终成功拯救出了表达IP10基因的rNDV-IP10病毒。为以后探究rNDV-IP10的联合抗瘤作用奠定基础。
[1] Vigil A, Martinez O, Chua MA, et al. Recombinant Newcastle disease virus as a vaccine vector for cancer therapy[J].Mol Ther, 2008,16(11):1883-1890.
[2] Li H, Peng KW, Russell SJ. Oncolytic measles virus encoding thyroidal sodium iodide symporter for squamous cell cancer of the head and neck radiovirotherapy[J]. Hum Gene Ther, 2012,23(3):295-301.
[3] Zhao H, Janke M, Fournier P, et al. Recombinant Newcastle disease virus expressing human interleukin-2 serves as a potential candidate for tumor therapy[J]. Virus Res, 2008,136(1-2):75-80.
[4] Gotoh B, Ohnishi Y, Inocencio NM, et al. Mammalian subtilisin-related proteinases in cleavage activation of the paramyxovirus fusion glycoprotein: superiority of furin/PACE to PC2 or PC1/PC3[J]. J Virol, 1992,66(11):6391-6397.
[5] Vigil A, Park MS, Martinez O, et al. Use of reverse genetics to enhance the oncolytic properties of Newcastle disease virus[J]. Cancer Res, 2007,67(17):8285-8292.
[6] Nishimura F, Dusak JE, Eguchi J, et al. Adoptive transfer of type 1 CTL mediates effective anti-central nervous system tumor response: critical roles of IFN-inducible protein-10[J]. Cancer Res, 2006,66(8):4478-4487.
[7] Giese NA, Raykov Z, DeMartino L, et al. Suppression of metastatic hemangiosarcoma by a parvovirus MVMp vector transducing the IP-10 chemokine into immunocompetent mice[J]. Cancer Gene Ther, 2002,9(5):432-442.
[8] Wang X, Zhang FC, Zhao HY, et al. Human IP10-scFv and DC-induced CTL synergistically inhibit the growth of glioma in a xenograft model[J]. Tumour Biol, 2014,35(8):7781-7791.
[9] Krishnamurthy S, Takimoto T, Scroggs RA, et al. Differentially regulated interferon response determines the outcome of Newcastle disease virus infection in normal and tumor cell lines[J]. J Virol, 2006,80(11):5145-5155.
[10] Wilden H, Fournier P, Zawatzky R, et al. Expression of RIG-I, IRF3, IFN-beta and IRF7 determines resistance or susceptibility of cells to infection by Newcastle Disease Virus[J]. Int J Oncol, 2009,34(4):971-982.
[11] Reichard KW, Lorence RM, Cascino CJ, et al. Newcastle disease virus selectively kills human tumor cells[J]. J Surg Res, 1992,52 (5):448-453.
[12] Laurie SA, Bell JC, Atkins HL, et al. A phase 1 clinical study of intravenous administration of PV701, an oncolytic virus, using two-step desensitization[J]. Clin Cancer Res, 2006,12(8):2555-2562.
[13] Fournier P, Arnold A, Wilden H, et al. Newcastle disease virus induces pro-inflammatory conditions and type I interferon for counter-acting Treg activity[J]. Int J Oncol, 2012,40(3):840-850.
[14] Freeman AI, Zakay-Rones Z, Gomori JM, et al. Phase I/II trial of intravenous NDV-HUJ oncolytic virus in recurrent glioblastoma multiforme[J]. Mol Ther, 2006,13(1):221-228.
[15] Wu Y, He J, An Y, et al. Recombinant Newcastle disease virus (NDV/Anh-IL-2) expressing human IL-2 as a potential candidate for suppresses growth of hepatoma therapy[J]. J Pharmacol Sci, 2016,132(1):24-30.
