广州市生鲜鸡中肠炎沙门菌的监测及耐药基因的分子鉴定
2018-03-13孙朋浩邱荣超李书宁郑双双杨伟聪张丽娜詹泽强张建民瞿孝云
孙朋浩,邱荣超,李书宁,郑双双,杨伟聪,张丽娜,符 颖,詹泽强,张建民,廖 明,瞿孝云*
(1.华南农业大学兽医学院人兽共患病防控制剂国家地方联合工程实验室,农业部人畜共患病重点实验室,广东省动物源性人兽共患病预防与控制重点实验室,广州 510642;2.河南牧业经济学院动物科技学院,郑州 450011; 3.云南省西双版纳州动物疫病预防控制中心,景洪 666100)
沙门菌(Salmonella)是一种重要的人兽共患病原菌,也是重要的食源性病原菌[1],据美国疾病预防控制中心统计,在美国每年大约有120万人感染沙门菌而患病,造成约450人死亡[2-3]。在我国每年大约有3亿人因沙门菌感染而患病,占我国病原菌型食源性疾病总数的70%~80%[4]。沙门菌暴发和随机感染的病例中大部分是由于摄入受污染的食物造成的,这些食物主要包括肉类食品、牛奶、鸡蛋、水果和蔬菜等,其中鸡肉被认为是沙门菌的主要载体,是人类感染沙门菌的一个重要源头[5]。广东省是鸡肉的生产和消费大省[6],关于广东省鸡肉中沙门菌的相关报道较少,缺乏相关的数据和信息。
肠炎沙门菌是造成食品污染和人源感染的沙门菌中最常见的血清型[7-8],近年来,食源中沙门菌的耐药性愈发严重[9],人们通过食用鸡肉感染肠炎沙门菌的概率远高于其他肉类产品[5],氟喹诺酮类药物和头孢类药物是目前临床治疗沙门菌感染的一线药物[9],密切关注肠炎沙门菌对β-内酰胺类和喹诺酮类药物的耐药情况,对合理使用抗生素和保障广州市鸡肉食品安全具有重要意义。
1 材料与方法
1.1 材料与试剂
1.1.1 材料 2016年3—12月,每月从广州市白云区、越秀区、天河区、海珠区、荔湾区的农贸市场随机采集现屠宰生鲜鸡样品,采样对象主要是鸡腿、鸡翅、鸡胸脯肉,每次在各个区采集样品约10份,使采样点尽可能的分布在不同档口,最后共采集了316份鸡肉样品。药敏实验质控菌株为大肠埃希菌ATCC25922,沙门菌标准菌株SalmonellaTyphimuriumLT2,均由本实验室保存。
1.1.2 主要试剂 缓冲蛋白胨水(BPW)、木糖-赖氨酸-硫酸四癸钠(XLT4)琼脂、四硫磺酸钠煌绿增菌液(TTB)、沙门菌显色板、LB营养琼脂、LB肉汤、无菌采样袋均购自青岛海博生物技术有限责任公司。沙门菌属诊断血清购自S&A公司。MH琼脂购自广州环凯微生物有限公司。16种药敏纸片:氨苄西林(ampicillin, AMP)、奥格门丁(amoxicillin-clavulanic acid, AMC)、头孢噻肟(cefotaxime, CTX)、头孢他啶(ceftazidime, CAZ)、头孢吡肟(cefepime, FEP)、庆大霉素(gentamicin, CEN)、链霉素(streptomycin, STR)、阿米卡星(amikacin, AMK)、复方新诺明(paediatric compound sulfamethoxazole tablets, SXT)、磺胺复合物(peptidylsulfanilamide, S3)、萘啶酸(nalidixic acid, NAL)、环丙沙星(ciprofloxacin, CIP)、氯霉素(chloramphenicol, CHL)、亚胺培南(imipenem, IPM)、多黏菌素B(polymyxin B sulfate, PB)、四环素(tetracycline, TET)均购自赛默飞世尔科技(中国)有限公司。TaqDNA聚合酶和1 000 bp Marker购自TaKaRa公司。
1.2 方法
1.2.1 沙门菌的分离鉴定 沙门菌分离的方法主要参考国家标准(GB4789.4-2016)。培养后,观察培养平板上菌落的形态,挑取典型菌落用麦康凯培养基进行纯化,取纯化后的沙门菌疑似菌落分别穿刺接种三糖铁和尿素斜面,37 ℃培养过夜,培养结果H2S阳性、尿酶阴性者即被认定为沙门菌,最后用沙门菌属诊断血清进行最终确定。
1.2.2 沙门菌血清型鉴定 鉴定方法按照泰国S&A 公司提供的沙门菌血清诊断操作步骤进行,查阅S&A公司沙门菌抗血清诊断附录,根据测定得到的抗原式确定沙门菌的血清型。
1.2.3 药敏试验 选取16种抗菌药物药敏纸片,根据美国临床实验室标准化协会(Clinical and Laboratory Standards Institute, CLSI)推荐的Kirby-Bauer法,对分离菌株进行操作,质控菌株为大肠埃希菌ATCC25922,结果参照赛默飞世尔科技公司提供的药敏试验参照表和CLSI制定的药敏试验抑菌环执行标准进行判断。
1.2.4 基因检测 本试验选择喹诺酮耐药决定区基因(quinolone resistance determining regions, QRDRs)gyrA、gyrB、parC、parE和质粒介导的诺酮类耐药基因(plasmid-mediated quinolone resistance, PMQR)qnrA、qnrB、qnrS、qnrC、qnrD、qepA为扩增对象,引物合成参照文献记载[10-16];选择β-内酰胺类耐药基因blaTEM、blaCTX-M、blaCMY、blaOXA、blaPSE、blaSHV为扩增对象,引物合成参考相关研究[17-18];选择Ⅰ类、Ⅱ类、Ⅲ类整合子基因盒插入区(可变区)的基因为扩增对象,引物合成参考文献[19],通过PCR方法和基因序列测定方法进行检测。
1.2.5 PCR扩增及序列测定 不同目的基因的PCR反应体系及反应条件略有差异,具体参照文献方法[10-15,17-19]。序列测定由广州天一辉远生物科技公司完成。
2 结 果
2.1 沙门菌分离鉴定及血清型
316份鸡肉样品中,阳性样品为243份,阳性率为76.9%。共鉴定23种血清型,主要血清型为阿贡纳(19.8%)、科瓦利斯(14.0%)、姆班达卡(11.9%)、肯塔基(10.3%)、肠炎(7.8%)、布伦登卢普(7.4%)(表1)。
2.2 药敏试验
肠炎沙门菌对萘啶酸(100.0%)、磺胺复合物(79.0%)、氨苄西林(57.9%)耐药严重,对链霉素(36.8%)、四环素(21.0%)、庆大霉素(21.0%)、头孢噻肟(15.8%)、头孢他啶(5.3%)耐药水平较低,对奥格门丁、头孢吡肟、阿米卡星、复方新诺明、环丙沙星、氯霉素、亚胺培南、多黏菌素B完全敏感,多重耐药率为68.4%(耐3类及其以上的抗生素)(表2~4)。19株肠炎沙门菌的耐药谱中,以AMP+STR+S3+NAL+TET、S3+NAL、NAL最多(15.8%),其次是AMP+STR+S3+NAL、AMP+CEN+S3+NAL+TET(10.5%);最长耐药谱为AMP+CTX+STR+S3+NAL+TET(5.3%)(表3)。
表1 243株沙门菌血清型分布
Table 1 243 strains ofSalmonellaserotype distribution
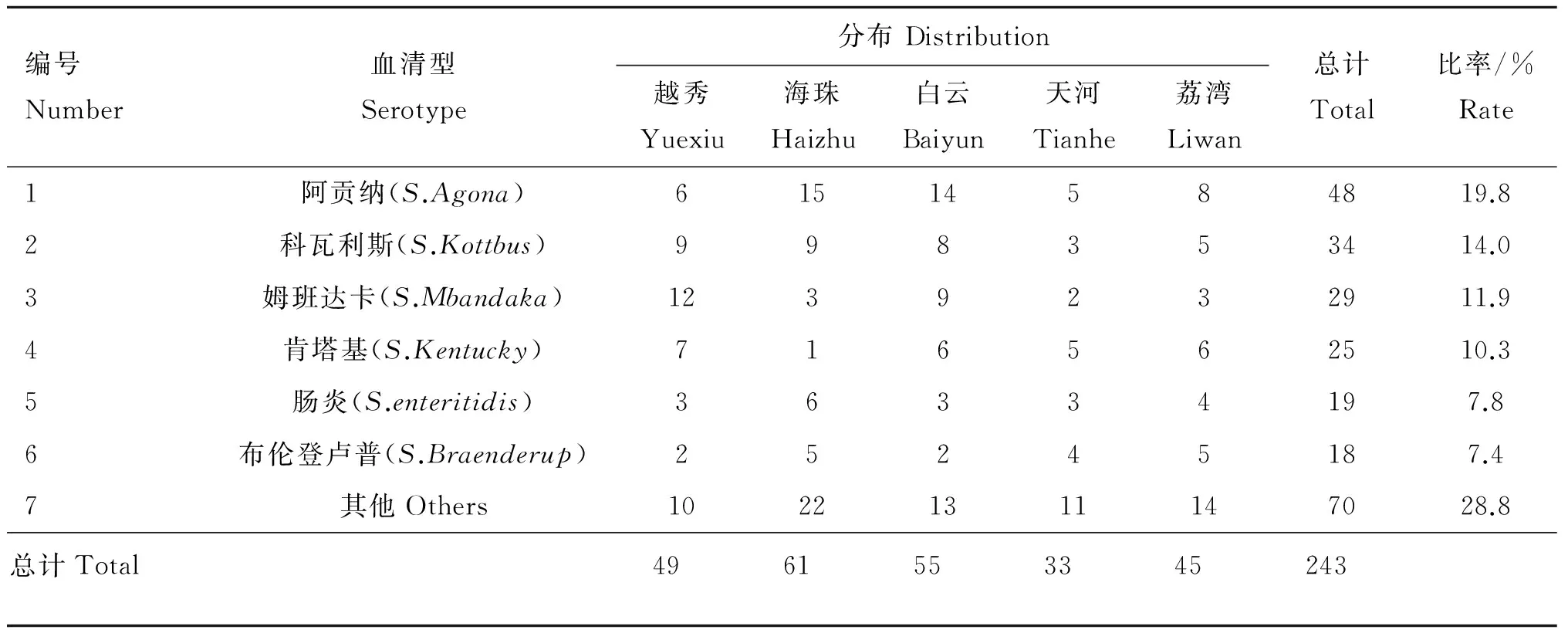
编号Number血清型Serotype分布Distribution越秀Yuexiu海珠Haizhu白云Baiyun天河Tianhe荔湾Liwan总计Total比率/%Rate1阿贡纳(S.Agona)61514584819.82科瓦利斯(S.Kottbus)998353414.03姆班达卡(S.Mbandaka)1239232911.94肯塔基(S.Kentucky)716562510.35肠炎(S.enteritidis)36334197.86布伦登卢普(S.Braenderup)25245187.47其他Others10221311147028.8总计Total4961553345243
表2 19株肠炎沙门菌耐药情况
Table 2 19 strains ofSalmonellaenteritidis antibiotic resistance
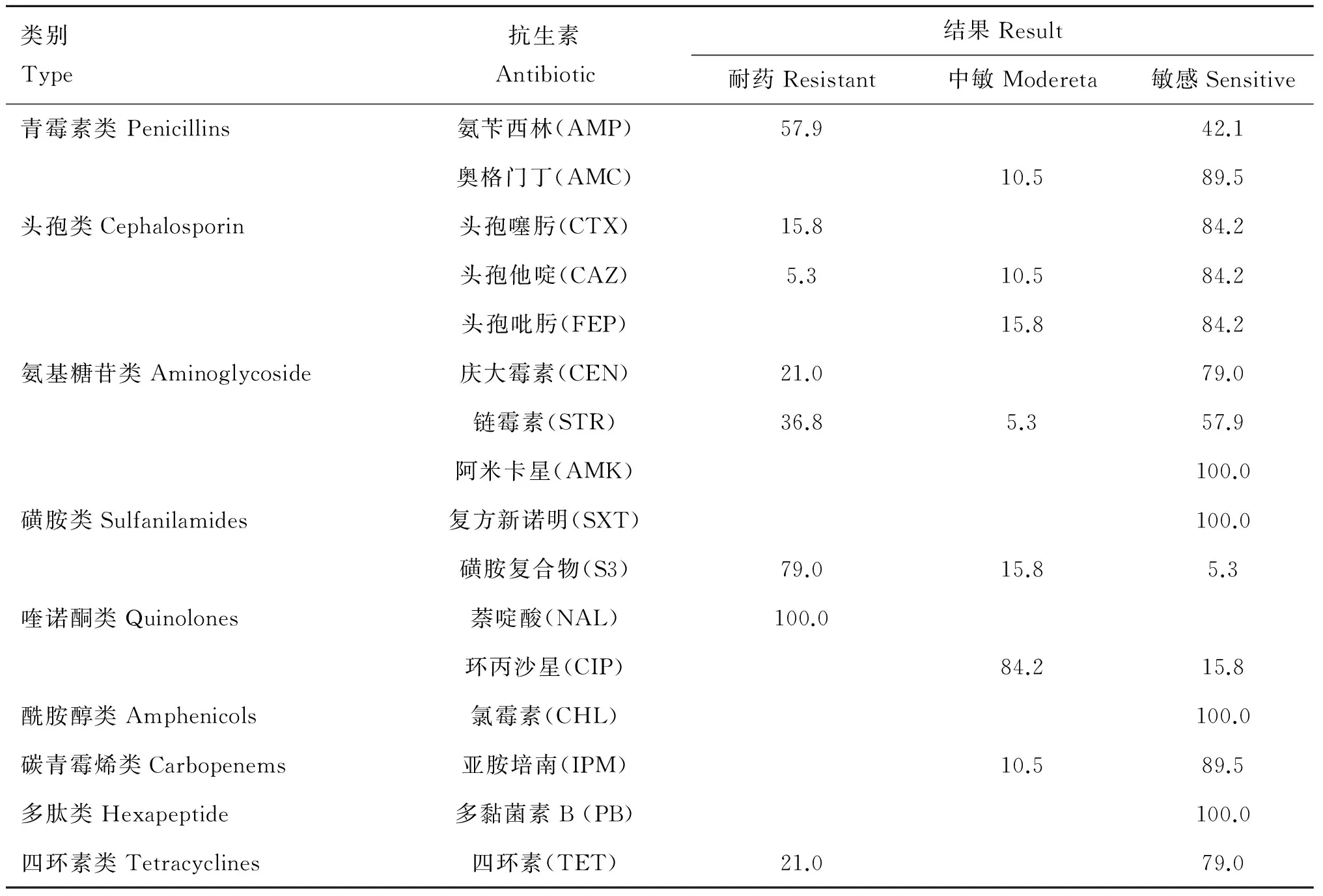
%
表3 19株肠炎沙门菌多重耐药谱
Table 3 Multiple antibiotic resistance profiles of 19 strains ofSalmonellaenteritidis
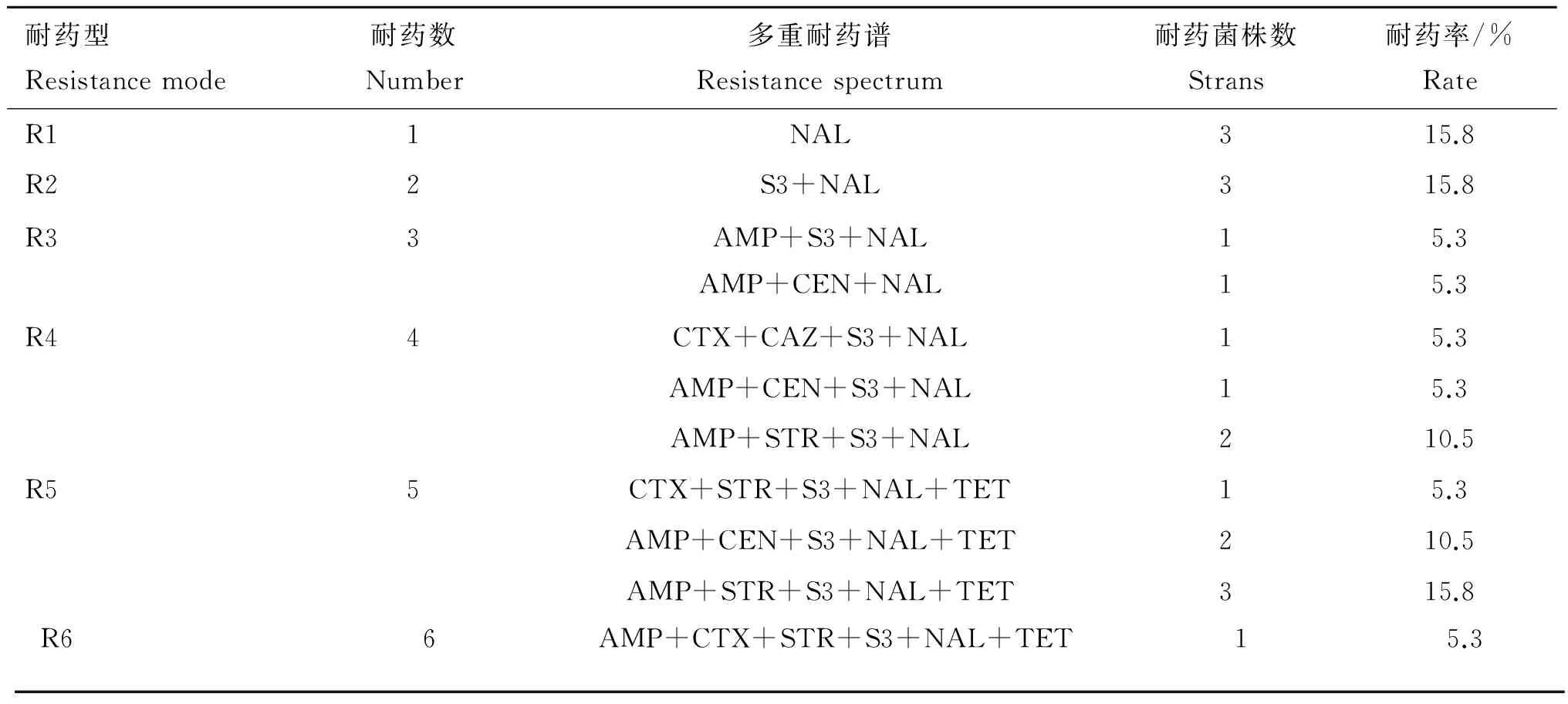
耐药型Resistancemode耐药数Number多重耐药谱Resistancespectrum耐药菌株数Strans耐药率/%RateR11NAL315.8R22S3+NAL315.8R33AMP+S3+NALAMP+CEN+NAL115.35.3R44CTX+CAZ+S3+NAL15.3AMP+CEN+S3+NAL15.3AMP+STR+S3+NAL210.5R55CTX+STR+S3+NAL+TET15.3AMP+CEN+S3+NAL+TET210.5AMP+STR+S3+NAL+TET315.8R66AMP+CTX+STR+S3+NAL+TET15.3
2.3 QRDRs的突变与PMQR的检测
19株肠炎沙门菌中,喹诺酮耐药决定区基因(QRDRs)的突变结果:gyrA、gyrB、parC基因中共检测出49个突变位点,parE未检测到突变。94.7%的菌株发生gyrA突变,突变类型有天冬氨酸-87→甘氨酸(42.1%)、天冬氨酸-87→色氨酸(31.6%)、丝氨酸-83→色氨酸(21.1%);73.7%的菌株发生gyrB突变,突变类型均为AAC-410→A-C(碱基位点缺失)和苯丙氨酸-414→丝氨酸(73.7%);15.8%的菌株发生parC突变,突变类型有AAC-25→A-C(碱基位点缺失,5.3%)、GAA-140→-AA(碱基位点缺失,10.5%);5.3%的菌株同时发生gyrA、parC突变,68.4%的菌株同时发生gyrA、gyrB突变,10.5%的菌株同时发生gyrA、gyrB、parC突变。19株肠炎沙门菌中,质粒介导的耐药基因(PMQR)检测结果:5.3%的菌株携带qepA,qnrA、qnrB、qnrC、qnrS、qnrD均未检测到;携带qepA的菌株同时发生gyrA、gyrB基因突变(表4)。
表4 19株肠炎沙门菌耐药情况与基因检测情况
Table 4 Drug resistance and gene detection of 19 strains ofSalmonellaenteritidis
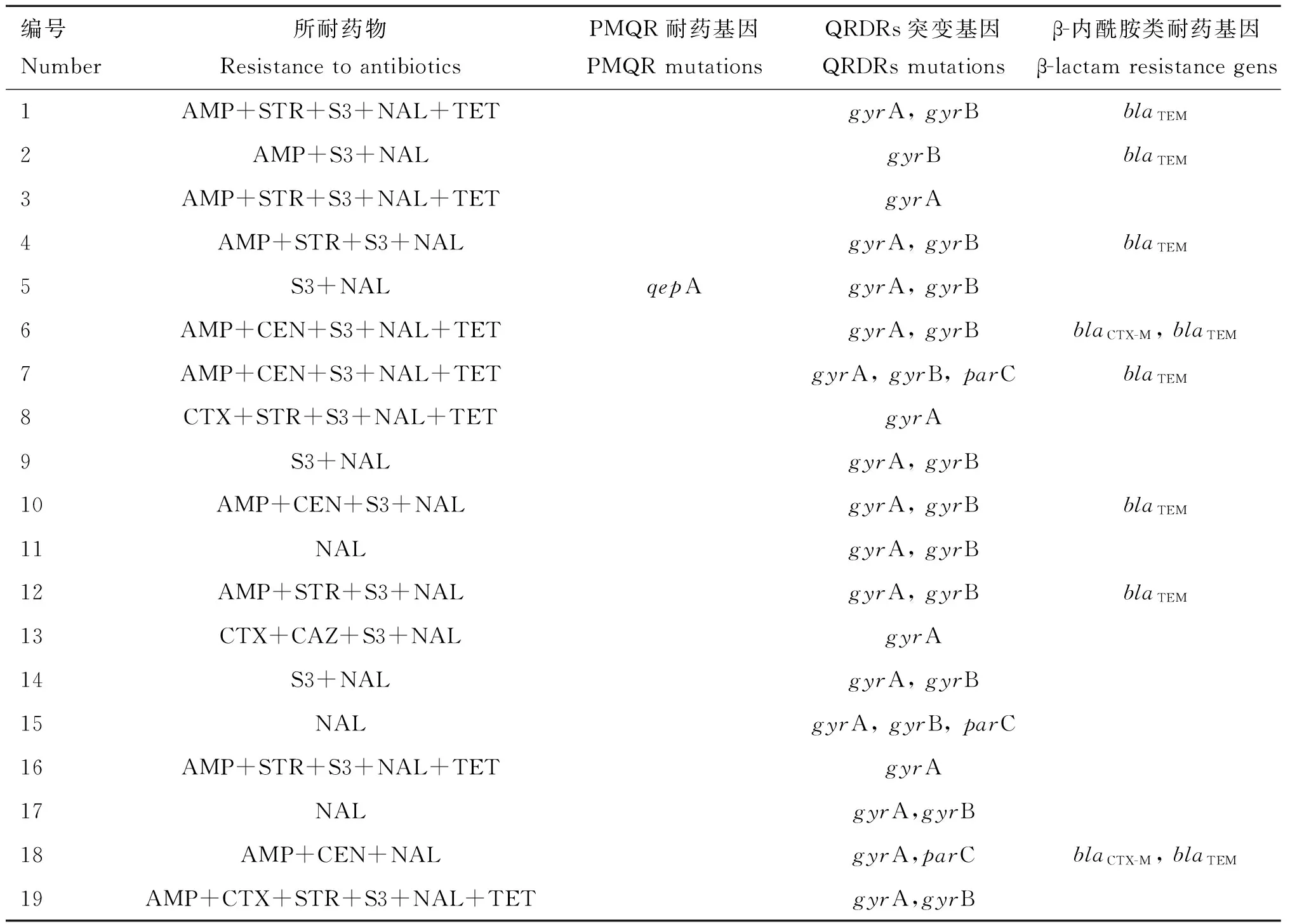
编号Number所耐药物ResistancetoantibioticsPMQR耐药基因PMQRmutationsQRDRs突变基因QRDRsmutationsβ-内酰胺类耐药基因β-lactamresistancegens1AMP+STR+S3+NAL+TETgyrA,gyrBblaTEM2AMP+S3+NALgyrBblaTEM3AMP+STR+S3+NAL+TETgyrA4AMP+STR+S3+NALgyrA,gyrBblaTEM5S3+NALqepAgyrA,gyrB6AMP+CEN+S3+NAL+TETgyrA,gyrBblaCTX-M,blaTEM7AMP+CEN+S3+NAL+TETgyrA,gyrB,parCblaTEM8CTX+STR+S3+NAL+TETgyrA9S3+NALgyrA,gyrB10AMP+CEN+S3+NALgyrA,gyrBblaTEM11NALgyrA,gyrB12AMP+STR+S3+NALgyrA,gyrBblaTEM13CTX+CAZ+S3+NALgyrA14S3+NALgyrA,gyrB15NALgyrA,gyrB,parC16AMP+STR+S3+NAL+TETgyrA17NALgyrA,gyrB18AMP+CEN+NALgyrA,parCblaCTX-M,blaTEM19AMP+CTX+STR+S3+NAL+TETgyrA,gyrB
1~19分别为19株肠炎沙门菌编号
The number that from 1 to 19 is the number ofSalmonellaenteritidisstrains
2.4 β-内酰胺类耐药基因的检测
19株肠炎沙门菌中,β-内酰胺类耐药基因检测结果:blaTEM、blaCTX-M的检出率分别为42.1%、10.5%,blaTEM和blaCTX-M的共同检出率为10.5%;blaCMY、blaOXA、blaPSE、blaSHV均未检测到。
2.5 Ⅰ类、Ⅱ类、Ⅲ类整合子的检测
Ⅰ类、Ⅱ类、Ⅲ类整合子均未检测到。
3 讨 论
沙门菌作为食源性疾病的头号致病菌而备受关注,世界卫生组织、美国和丹麦等国家相继建立沙门菌监测系统,近年来,我国也日益重视对沙门菌的监测和研究工作。本次试验中,鸡肉中沙门菌检出率为76.9%,此前,杨保伟(B.W.Yang)等[20]报道的在陕西省鸡肉中沙门菌的检出率为54%,林兰等[21]报道北京市生鲜整鸡中沙门菌的检出率为49.9%,通过比较不同地区间鸡肉中沙门菌的检出率,可以发现广州市鸡肉中沙门菌的污染率远高于其他地区,原因可能在于广州地处华南地区,常年温度偏高,气候湿润,沙门菌易于生存,同时华南地区是鸟类迁徙的集散地,鸟类的迁徙活动促进了沙门菌的传播,这要求我们应密切监控华南地区沙门菌在食源中的流行情况。据报道,日、美等发达国家发生的食物中毒事件中40%~80%是由禽沙门菌引起的,其中主要病原为肠炎沙门菌。1989年3月世界卫生组织专门召开了一次有关家禽及禽蛋中肠炎沙门菌污染问题的紧急国际会议。许少洪等[22]对广州海珠地区2002—2011年人源感染沙门菌的血清型分布的研究表明,广州市临床人源感染的沙门菌中,肠炎沙门菌占据相当大的比例。人源沙门菌感染的病例中大部分是由于摄入受污染的食物造成的,其中鸡肉被认为是沙门菌的主要载体[5],因此密切关注广州市鸡肉中肠炎沙门菌的流行情况和耐药情况,为保障广州市的公共卫生安全具有重要意义。
通过对19株肠炎沙门菌进行药敏试验,发现肠炎沙门菌对萘啶酸(100.0%)、磺胺复合物(79.0%)、氨苄西林(57.9%)严重耐药,这与国内外有关报道基本一致[23-24],原因可能在于这些传统抗菌药物长期以来被广泛应用于畜禽养殖中,加之不规范的用药方法,抗菌药物的选择压力使细菌产生了较高的耐药性;对头孢噻肟(15.8%)、头孢他啶(5.3%)较为敏感,对头孢吡肟、环丙沙星表现为完全敏感,这表明三代头孢和氟喹诺酮类药物对肠炎沙门菌仍具有较好的抑制杀灭效果,我们在选择用药时可以参考。
抗生素靶位编码基因突变导致表达产物的空间构型与理化性质发生变化,进而致使药物的结合作用下降或消失而产生耐药性是常见的耐药机制[25]。沙门菌对喹诺酮类药物的耐药性主要与其细胞中QRDRs基因(gyrA、gyrB、parC、parE)的突变有关[26]。本试验中,通过对QRDRs基因检测发现,100%肠炎沙门菌至少有一个QRDRs亚单位发生基因位点突变,值得注意的是,本试验中肠炎沙门菌对萘啶酸的耐药率为100%,QRDRs的突变结果与沙门菌对喹诺酮类药物的耐药结果具有高度的一致性。细菌对喹诺酮类的敏感性主要由作用靶位决定,主要靶位的点突变可引起细菌对喹诺酮类敏感性的降低,多个点突变或同时存在次要靶位的突变使耐药程度进一步上升[25]。与此同时,耐药质粒也是导致沙门菌对喹诺酮类药物产生抗性的重要机制[27-28],它们可以通过接合或转导作用在不同的细菌之间进行转移,介导细菌产生低水平耐药。本试验中qnrA、qnrB、qnrS、qnrC、qnrD均未被检出,qepA的检出率为5.3%,qepA基因被认为是新的质粒介导的喹诺酮类耐药的分子机制,可通过外排泵机制介导细菌对亲水性喹诺酮类药物的耐药性[29],为肠炎沙门菌对氟喹诺酮类药物产生耐药性创造了潜在的条件。分析QRDRs和PMQR的检测结果,可以推测:QRDRs位点突变导致细菌合成氨基酸的改变进而引起合成蛋白质的空间构型与理化性质发生变化是本试验中肠炎沙门菌对萘啶酸完全耐药的重要原因。
沙门菌通过产生β-内酰胺酶,水解β内酰胺环使药物失活,是沙门菌对β-内酰胺类药物产生耐药性的主要原因。本试验中,β-内酰胺类耐药基因的检出率为42.1%,这与冯彩峰[30]之前报道的结果基本一致。结合表4分析发现,β-内酰胺类的耐药基因的检出结果与肠炎沙门菌对氨苄西林的耐药结果高度一致(100%),β-内酰胺类耐药基因的携带为菌株耐受三代头孢类药物创造了潜在条件,这给我们在生产用药中以警示。整合子在整合酶的催化下,通过特异性结合位点捕获外源基因(特别是耐药基因)并使之表达,同时整合子又可整合到质粒或染色体上,或自身作为转座子的一个组成部分而参与转移,使耐药基因在同种和不同种属细菌间广泛传播[31]。本试验中,Ⅰ类、Ⅱ类、Ⅲ类整合子的检测结果均为阴性,这与前面所介绍的喹诺酮类耐药质粒的低检出率相互印证。
4 结 论
广州地区鸡肉中沙门菌的污染率高,血清型类型复杂。肠炎沙门菌仍是目前重要的血清型,对传统抗生素的耐药情况较为严重,对头孢吡肟、环丙沙星等药物完全敏感,喹诺酮耐药决定区基因(QRDRs)突变结果和β-内酰胺类耐药基因的检出结果与肠炎沙门菌的耐药结果高度一致。
[1] ZHANG J, JIN H, HU J, et al. Serovars and antimicrobial resistance of non-typhoidalSalmonellafrom human patients in Shanghai, China, 2006-2010[J].EpidemiolInfect, 2014, 142(4): 826-832.
[2] FOLEY S L, LYNNE A M. Food animal-associatedSalmonellachallenges: pathogenicity and antimicrobial resistance[J].JAnimSci, 2008, 86(14S): E173-E187.
[3] 胡 燕. 2008-2009美国鼠伤寒沙门菌疫情调查报告[J]. 现代预防医学, 2010, 37(10): 1964-1965. HU Y. Investigation information for outbreak ofSalmonellatyphimurium infections in United States from 2008 to 2009[J].ModernPreventiveMedicine, 2010, 37(10): 1964-1965. (in Chinese)
[4] YANG B W, CUI Y, SHI C, et al. Counts, serotypes, and antimicrobial resistance ofSalmonellaisolates on retail raw poultry in the People’s Republic of China[J].JFoodProt, 2014, 77(6): 894-902.
[5] ARVANITIDOU M, TSAKRIS A, SOFIANOU D, et al. Antimicrobial resistance and R-factor transfer ofSalmonellaeisolated from chicken carcasses in Greek hospitals[J].IntJFoodMicrobiol, 1998, 40(3): 197-201.
[6] 李世聪, 闻 剑, 邓小玲, 等. 广东省城市居民食物和营养素摄入现状及变化趋势[J]. 中国公共卫生, 2014, 30(9): 1109-1112. LI S C, WEN J, DENG X L, et al. Status and change trend of food consumption and nutrients intake in urban population of Guangdong province[J].ChineseJournalofPublicHealth, 2014, 30(9): 1109-1112. (in Chinese)
[7] 梁兆铭, 柯碧霞, 邓小玲, 等. 2010-2011年广东地区人源主要血清型沙门菌喹诺酮耐药特征分析[J]. 华南预防医学, 2013, 39(6): 27-32. LIANG Z M, KE B X, DENG X L, et al. Characteristics of main human-sourceSalmonellaserovars resistant to quinolone in Guangdong Province, 2010-2011[J].SouthChinaJournalofPreventiveMedicine, 2013, 39(6): 27-32. (in Chinese)
[8] 陈玉贞, 邵 坤, 关 冰, 等. 2003-2010年山东省食源性沙门菌血清分型及药敏分析[J]. 中国食品卫生杂志, 2012, 24(1): 9-13. CHEN Y Z, SHAO K, GUAN B, et al. An analysis on the serotypes and antibiotic sensitivities of foodborneSalmonellain Shandong province from 2003 to 2010[J].ChineseJournalofFoodHygiene, 2012, 24(1): 9-13. (in Chinese)
[9] 炊慧霞, 张秀丽, 廖兴广, 等. 河南省2003-2008年食源性沙门菌耐药性变迁研究[J]. 现代预防医学, 2010, 37(18): 3542-3543, 3547. CHUI H X, ZHANG X L, LIAO X G, et al. Study on the resistance changes of foodborneSalmonellain Henan from 2003 to 2008[J].ModernPreventiveMedicine, 2010, 37(18): 3542-3543, 3547. (in Chinese)
[10] O′REGAN E, QUINN T, PAGS J M, et al. Multiple regulatory pathways associated with high-level ciprofloxacin and multidrug resistance inSalmonellaentericaserovar enteritidis: involvement oframAand other global regulators[J].AntimicrobAgentsChemother, 2009, 53(3): 1080-1087.
[11] ROBICSEK A, JACOBY G A, HOOPER D C. The worldwide emergence of plasmid-mediated quinolone resistance[J].LancetInfectDis, 2006, 6(10): 629-640.
[12] GAY K, ROBICSEK A, STRAHILEVITZ J, et al. Plasmid-mediated quinolone resistance in Non-Typhi serotypes ofSalmonellaenterica[J].ClinInfectDis, 2006, 43(3): 297-304.
[13] WANG M H, GUO Q L, XU X G, et al. New plasmid-mediated quinolone resistance geneqnrC, found in a clinical isolate ofProteusmirabilis[J].AntimicrobAgentsChemother, 2009, 53(5): 1892-1897.
[14] CAVACO L M, HASMAN H, XIA S, et al.qnrD, a novel gene conferring transferable quinolone resistance inSalmonellaentericaserovar Kentucky and Bovismorbificans strains of human origin[J].AntimicrobAgentsChemother, 2009, 53(2): 603-608.
[15] 陈孝杰. 不同来源大肠杆菌中喹诺酮外排泵基因oqxAB和qepA的传播机制研究[D]. 广州: 华南农业大学, 2016. CHEN X J. The transmission mechanisms of plasmid-mediated quinolone resistance efflux pumps genesoqxABandqepAinEscherichiacoliisolates from different sources[D]. Guangzhou: South China Agricultural University, 2016. (in Chinese)
[16] KEHRENBERG C, FRIEDERICHS S, DE JONG A, et al. Identification of the plasmid-borne quinolone resistance geneqnrSinSalmonellaentericaserovar Infantis[J].JAntimicrobChemother, 2006, 58(1): 18-22.
[17] QIAO J, ZHANG Q, ALALI W Q, et al. Characterization of extended-spectrumβ-lactamases (ESBLs)-producingSalmonellain retail raw chicken carcasses[J].IntJFoodMicrobiol, 2017, 248: 72-81.
[18] NGUYEN D T A, KANKI M, NGUYEN P D, et al. Prevalence, antibiotic resistance, and extended-spectrum and AmpCβ-lactamase productivity ofSalmonellaisolates from raw meat and seafood samples in Ho Chi Minh City, Vietnam[J].IntJFoodMicrobiol, 2016, 236: 115-122.
[19] 伍 好. 肉鸡源性肠杆菌科细菌中耐药基因、整合子及ISCR1元件的研究[D]. 济南: 山东大学, 2016. WU H. The research on the antibiotic resistance genes, integrons and ISCR1 elements amongEnterobacteriaceaefrom chicken broilers[D]. Jinan: Shandong University, 2016. (in Chinese)
[20] YANG B W, QU D, ZHANG X L, et al. Prevalence and characterization ofSalmonellaserovars in retail meats of marketplace in Shaanxi, China[J].IntJFoodMicrobiol, 2010, 141(1-2): 63-72.
[21] 林 兰, 王晔茹, 徐 潇, 等. 零售生鲜整鸡中沙门氏菌分离与耐药性分析[J]. 药物生物技术, 2014, 21(4): 343-348. LIN L, WANG Y R, XU X, et al. Isolation and antimcirobial resistance ofSalmonellaentericafrom retail chicken carcasses[J].PharmaceuticalBiotechnology, 2014, 21(4): 343-348. (in Chinese)
[22] 许少洪, 孙凤琪, 曾爱芳, 等. 广州海珠地区2002年-2011年沙门菌血清型分布[J]. 中国卫生检验杂志, 2013, 23(2): 466-469. XU S H, SUN F Q, ZENG A F, et al. A study on the distribution ofSalmonellaserotypes in Haizhu district of Guangzhou during 2002 and 2011[J].ChineseJournalofHealthLaboratoryTechnology, 2013, 23(2): 466-469. (in Chinese)
[23] 杨保伟, 申进玲, 席美丽, 等. 2007-2008年西安地区鸡肉源沙门氏菌相关特性分析[J]. 食品科学, 2011, 32(19): 130-136. YANG B W, SHEN J L, XI M L, et al. Antibacterial susceptibility and subtypes ofSalmonellaisolates from retail chicken in Xi’an in 2007-2008[J].FoodScience, 2011, 32(19): 130-136. (in Chinese)
[24] ZIECH R E, LAMPUGNANI C, PERIN A P, et al. Multidrug resistance and ESBL-producingSalmonellaspp. isolated from broiler processing plants[J].BrazilJMicrobiol, 2016, 47(1): 191-195.
[25] 郝宏珊, 杨保伟, 师俊玲, 等. 鸡肉源沙门氏菌对喹诺酮和氟喹诺酮类抗生素耐药状况及相关基因[J]. 微生物学报, 2011, 51(10): 1413-1420. HAO H S, YANG B W, SHI J L, et al. Drug resistance and related genes of chickenborneSalmonellato quinolone and fluoroquinolones[J].ActaMicrobiologicaSinica, 2011, 51(10): 1413-1420. (in Chinese)
[26] CASIN I, BREUIL J, DARCHIS J P, et al. Fluoroquinolone resistance linked to gyrA, gyrB, and parC mutations inSalmonellaentericatyphimurium isolates in humans[J].EmergInfectDis, 2003, 9(11): 1455-1457.
[27] AMBLER J E, PINNEY R J. Positive R plasmid mutator effect on chromosomal mutation to nalidixic acid resistance in nalidixic acid-exposed cultures of Escherichia coli[J].JAntimicrobChemother, 1995, 35(5): 603-609.
[28] SJÖLUND-KARLSSON M, FOLSTER J P, PECIC G, et al. Emergence of plasmid-mediated quinolone resistance among non-TyphiSalmonellaentericaisolates from humans in the United States[J].AntimicrobAgentsChemother, 2009, 53(5): 2142-2144.
[29] 余 华, 李从荣, 李 艳, 等. 质粒介导的喹诺酮类耐药基因qepA的流行现状及耐药机制研究[J]. 中华检验医学杂志, 2011, 34(6): 526-528. YU H, LI C R, LI Y, et al. Prevalent situation and mechanism for plasmid-mediated quionlone resistance geneqepA[J].ChineseJournalofLaboratoryMedicine, 2011, 34(6): 526-528. (in Chinese)
[30] 冯彩峰. 食品动物源沙门氏菌耐药性及耐β-内酰胺类基因的检测[D]. 雅安: 四川农业大学, 2014. FENG C F. The detection of resistance andβ-lactams resistance genes inSalmonellaisolates from food-animals[D]. Ya'an: Sichuan Agricultural University, 2014. (in Chinese)
[31] SUNDE M, NORDSTROM M. The prevalence of, associations between and conjugal transfer of antibiotic resistance genes inEscherichiacoliisolated from Norwegian meat and meat products[J].JAntimicrobChemother, 2006, 58(4): 741-747.
