8个LA百合品种的核型分析
2018-03-12霍辰思白锦荣窦晓莹郎利新孔滢尚宏忠
霍辰思,白锦荣,窦晓莹,郎利新,孔滢,尚宏忠
(1.山西省农业科学院 果树研究所,山西 太原 030031; 2.北京市辐射中心,北京 100875)
百合是百合科(Liliaceae)百合属(Lilium)植物的总称,其花大色艳,姿态优美,品种繁多,是国内外花卉市场上五大鲜切花之一。百合主要有亚洲百合杂种系(Asiatic hybrids),东方百合杂种系(Oriental hybrids),喇叭百合杂种系(Trumet hybrids),麝香百合杂种系(Longiflorum hybrids)及LA(亚洲百合与麝香百合杂种系间的杂交种)、OA(东方百合杂种系与亚洲百合杂种系间的杂交种)、OT(东方百合杂种系与喇叭百合杂种系间的杂交种)等系间杂种系[1,2]。LA系列在栽培和观赏方面优势突出[3],继承了麝香百合的喇叭花型、强壮的花茎,又遗传了亚洲百合的柔和色调和坚挺的花被,受到市场的广泛欢迎。
植物核型的研究有助于揭示物种间的亲缘关系及遗传进化规律,为新品种选育提供可靠的细胞学依据[4~7]。目前百合属的细胞学研究主要集中于野生种和部分品系中[8~12]。由于面世较晚,LA百合品系的研究相比于其他百合品种较匮乏,对其染色体核型文献报道较少。本研究选取8个LA百合品种作为研究对象,对其染色体数目及核型进行分析,旨在从细胞学层面分析LA百合品系的遗传特点,为百合遗传育种中亲本选择和后代鉴定提供可靠的染色体信息。
1 材料与方法
1.1 材料
供试材料共8份LA百合,分别为:‘Cilesta’、‘Couplet’、‘Eyeliner’、‘Menorca’、‘Serrada’、‘Donau’、‘Desiderio’和‘White sound’,所有品种均购自于荷兰,保存于北京市辐射中心育种资源圃。
1.2 方法
染色体制片采用常规压片法。于2014年4月上午9:00-11:00,取生长旺盛的根尖,蒸馏水洗净后,用放线菌酮(200 mmol·L-1)在室温下暗处理7~8 h。将预处理8 h 的材料用卡诺氏固定液(无水乙醇∶冰醋酸=3∶1)固定过夜,4 ℃保存备用,若长期保存可将材料置于70%酒精中4 ℃下保存。将固定好的根尖用纤维素酶与果胶酶混合液在37 ℃恒温水浴中解离1 h,用蒸馏水冲洗材料后,2%卡宝品红染液进行染色。压片后用Olympus (BX-51) 显微镜进行镜检[13,14]。
每个品种观察20~25个细胞以确定染色体的数目,选择5个分散好、形态清晰的中期分裂相进行染色体分析测量,得到相对应的核型数据。同一条染色体重复3次,求其平均值。核型的分析按照李懋学制定标准进行[15]。染色体的相对长度、臂比以及类型参照Levan等命名系统[16]。核型的类型根据Stebbins的分类标准进行划分[17],核型不对称系数按Arano的方法进行计算[18]。
2 结果与分析
对8个LA百合进行染色体数目及核型分析,染色体参数和核型类型见表1。中期染色体照片、核型图与核型模式图见图1。结果表明,供试的8个LA百合品种均为三倍体,2n=3x=36,其中,7个品种染色体核型类型为3B型,只有‘Serrada’核型类型为3A型。
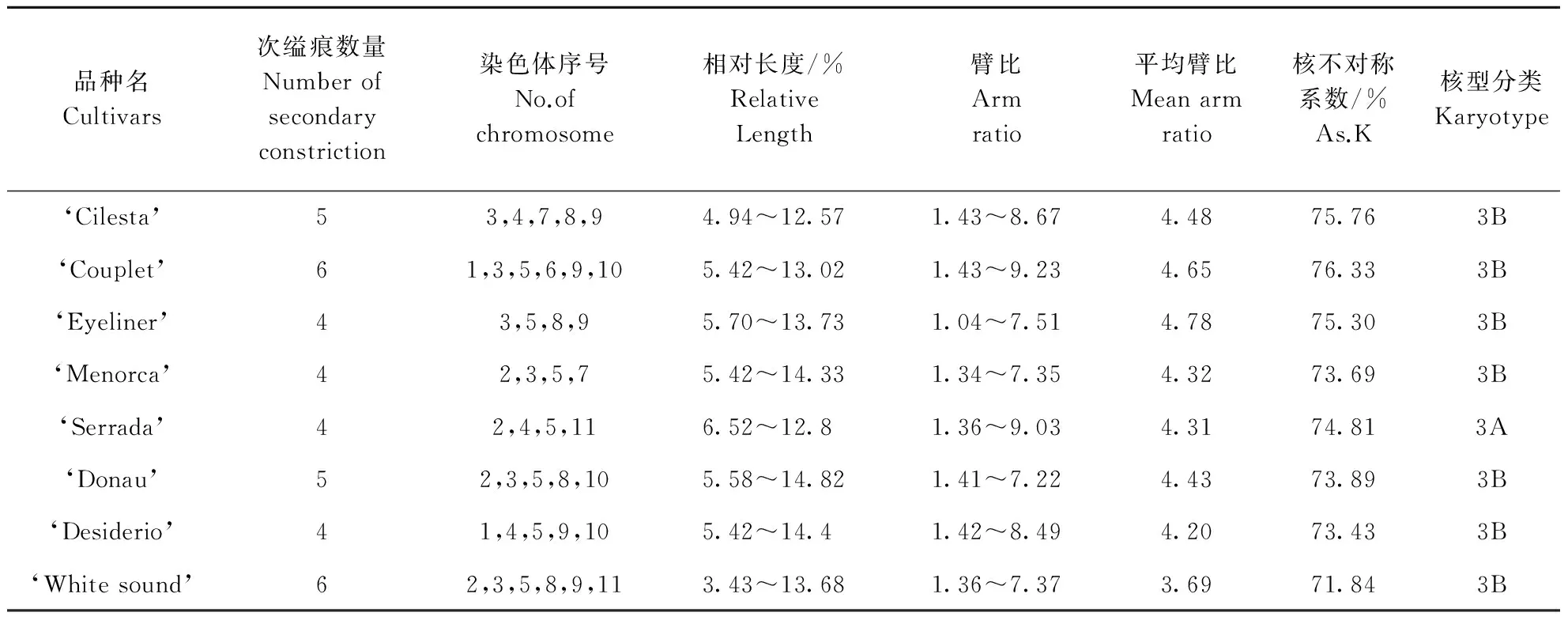
表1 8个LA百合品种的核型分类及相关参数Table 1 The comparison of karyotype on 8 LA hybrids
2.1 ‘Cilesta’的核型分析
‘Cilesta’的核型公式为2n=3x=36=6m+6sm(6SAT)+15st(6SAT)+9t。相对长度的范围为4.94%~12.57%,平均臂比为4.488,最长的与最短的染色体长度比为2.54,臂比值大于2的染色体占总数的75%,核型不对称系数为75.8%,属于3B型(表1)。第1、2对染色体着丝点位置在中部(m),第3、6对染色体着丝点位置在近中部(sm),其余为近端部(st)或端部(t)着丝点染色体。编号3、4、7、8染色体短臂上有次缢痕,编号9染色体长臂端部具次缢痕(图1,a)。
2.2 ‘Couplet’的核型分析
‘Couplet’的核型公式为2n=3x=36=6m+6sm(6SAT)+18st(12SAT)+6t。相对长度范围为5.42%~13.02%,平均臂比为4.647,最长的与最短的染色体长度比为2.4,臂比值大于2的染色体占总数的75%,核型不对称系数为76.32%,属于3B型(表1)。第1、10对染色体是近中部(sm)着丝点染色体,第2、8对染色体是中部着丝点染色体(m),其余均为近端部(st)或端部(t)着丝点染色体。编号1、3、5、6、9、10染色体短臂端部有次缢痕(图1,b)。
2.3 ‘Eyeliner’的核型分析
‘Eyeliner’的核型公式为2n=3x=36=9m(3SAT)+3sm(3SAT)+18st(12SAT)+6t。相对长度范围为5.70%~13.73%,平均臂比为4.778,最长的与最短的染色体长度比为2.42,臂比值大于2的染色体占总数的75%,核型不对称系数为75.31%,属于3B型(表1)。第1、2、7对着丝点位置在中部(m),第3对着丝点位置为近中部(sm),其余均为近端部(st)或端部(t)着丝点染色体。编号3、5染色体长臂上有次缢痕,编号8、9染色体短臂端部有一条次缢痕(图1,c)。

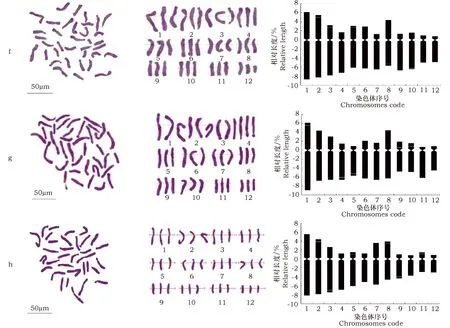
图1 8个LA百合品种中期染色体形态图Fig.1 Metaphase chromosomes of 8 LA hybrids 注:a.‘Cilesta’;b.‘Couplet’;c.‘Eyeliner’;d.‘Menorca’;e.‘Serrada’;f.‘Donau’;g‘Desiderio’;h.‘White sound’Note: a.‘Cilesta’;b.‘Couplet’;c.‘Eyeliner’;d.‘Menorca’;e.‘Serrada’;f.‘Donau’;g‘Desiderio’;h.‘White sound’
2.4 ‘Menorca’的核型分析
‘Menorca’的核型公式为2n=3x=36=9m(3SAT)+6sm(6SAT)+12st(9SAT)+9t。相对的长度范围为5.42%~14.33%,平均臂比为4.238,最长的与最短的染色体长度比为2.64,臂比值大于2的染色体占总数的75%,核型不对称系数为73.69%,属于3B型(表1)。第3、9对为近中部(sm)着丝点染色体,第1、2、8对染色体为中部(m)着丝点染色体,其余为近端部(st)或端部(t)着丝点染色体。编号3长臂上有次缢痕,编号2、5、7染色体短臂端部有次缢痕(图1,d)。
2.5 ‘Serrada’的核型分析
‘Serrada’的核型公式为2n=3x=36=6m(3SAT)+9sm(3SAT)+12st(6SAT)+9t。相对长度范围为6.52%~12.8%,平均臂比为4.312,最长的与最短的染色体长度比为1.97,臂比值大于2的染色体占总数的83.3%,核型不对称系数为74.82%,属于3A型(表1)。第3、4、6对染色体为近中部(sm)着丝点染色体,第1、2对染色体为中部(m)着丝点染色体,其余均为近端部(st)或端部(t)着丝点染色体。编号2、11染色体短臂上有次缢痕,编号为4、5的长臂上有次缢痕(图1,e)。
2.6 ‘Donau’的核型分析
‘Donau’的核型公式为2n=3x=36=9m(6SAT)+9sm(6SAT)+12st(3SAT)+6t。相对长度范围为5.58%~14.82%,平均臂比为4.43,最长的与最短的染色体长度比为2.66,臂比值大于2的染色体占总数的75%,核型不对称系数为73.88%,属于3B型(表1)。第1、2、8对染色体为中部(m)着丝点染色体,第3、5、6对染色体为近中部(sm)着丝点染色体,其余均为端部(t)或近端部(st)着丝点染色体。编号2、3、5染色体的短臂上有次缢痕,编号8、10的长臂上有次缢痕(图1,f)。
2.7 ‘Desiderio’的核型分析
‘Desiderio’的核型公式为2n=3x=36=9m(3SAT)+6sm+15st(9SAT)+6t(3SAT)。相对长度范围为5.42%~14.4%,平均臂比为4.194,最长的与最短的染色体长度比为2.65,臂比值大于2的染色体占总数的75%,核型不对称系数为73.42%,属于3B型(表1)。第3、6对染色体为近中部(sm)着丝点染色体,第1、2、8对染色体为中部(m)着丝点染色体,其余均为端部(t)或近端部(st)着丝点染色体。编号4、5的染色体长臂上有次缢痕,9号染色体具有居间随体,编号1、10的染色体短臂上有次缢痕(图1,g)。
2.8 ‘White sound’的核型分析
‘White sound’的核型公式为2n=3x=36=9m(3SAT)+9sm(9SAT)+15st(6SAT)+3t。相对长度范围为3.43%~13.68%,平均臂比为3.695,最长的与最短的染色体长度比为3.98,臂比值大于2的染色体占总数的75%,核型不对称系数为71.83%,属于3B型(表1)。第7、8对染色体为中部(m)着丝点染色体,第1、2、3、11对染色体为近中部(sm)着丝点染色体,其余均为端部(t)或近端部(st)着丝点染色体。编号2、5、8、11的染色体短臂具次缢痕,编号3、9的染色体长臂具次缢痕(图1,h)。
3 讨论与结论
百合属野生资源大部分为二倍体,仅有卷丹、川百合等少数种为三倍体[19,20],部分品种也有三倍体。三倍体是由于来自亲本的组间杂交F1代产生未减数分裂的2n配子,从而产生三倍体回交一代有性多倍化的过程[21]。
LA百合杂种系为麝香百合与亚洲百合远缘杂交一代后与亚洲百合回交的品种,大部分为异源三倍体。本试验确定供试的8个LA百合品种均为三倍体(2n=3x=36)。百合属的染色体核型多为稳定的3B型[22],少数种类为3A或4B型[23],本研究试验结果与此结论相符。戴小红等发现百合品种‘Pollyanna’为3A型[8]。本试验中除‘Serrada’核型类型为3A型外,其余7个品种染色体核型类型是3B型。
研究表明,对育种亲本的核型背景进行分析,可以为杂交亲本的选配和后代鉴定提供一定的细胞学依据。染色体的随体、次缢痕的特征可做为杂交后代早期鉴定的依据[24]。LA百合杂种系的亲本为亚洲百合和麝香百合,其中,麝香百合多为二倍体,而亚洲百合倍性比较复杂,多数为二倍体,也有三倍体、四倍体以及非整倍体[25]。周桂雪等在对5个亚洲百合品种进行倍性鉴定时,发现‘Brunello’和‘ValdiSole’为四倍体(2n=4x=48),并对这些四倍体百合与二倍体百合进行杂交,结合胚拯救技术,得到少量的三倍体杂交后代[26]。国内外学者利用核型分析中次缢痕的有无对百合杂种进行早期鉴定,如李卫民等报道的福建麝香百合(L.longiflorumThumb.)第6条染色体的短臂上具有次缢痕,核型为3A型[27]。LA百合品系染色体较其亲本染色体比例大,个数多,次缢痕也较其亲本明显。本试验中的8个LA百合品种,除了在其短臂上存在次缢痕,各个品种长臂上也有次缢痕,而其亲本次缢痕多数存在于染色体短臂上。‘Serrada’4、5号染色体长臂上有次缢痕,进一步证明LA百合杂种系较其亲本更加进化。目前,栽培品种的核型分析并不能够完整反映出不同系列百合品种的共同遗传特征,通过核型分析可以得到共性特征,尤其能够找到相似程度较高的带有次缢痕的染色体,这对于种群与种间杂种后代的鉴定以及亲缘关系研究具有十分重要的意义。
Stebbins认为,高等植物核型进化的基本趋势应该是由对称向不对称的方向发展,核不对称系数越高说明其进化程度越高[17]。百合属植物的染色体较大且基数多,本试验对8种LA百合核不对称系数进行分析,结果表明其核型不对称系数均在70% 以上,属于较进化类型,这也是人为干预选择的结果。百合三倍体雄性不育,但是以三倍体LA百合为母本,与二倍体或者四倍体进行杂交授粉,将增加2n配子变异程度,通过百合种质渗入育种,提高后代筛选的机会[26]。大多数LA百合杂种系都含有基因组重组染色体,减数分裂产生2n配子的同时也可以产生其他重组染色体,这些重组染色体对育种研究有重大潜力,研究者可以利用这些非整倍体在杂交后代中筛选出目标性状优良的植株,培育更有经济价值的百合新种质。
[1]龙雅宜,张金政,张兰年,等.百合—球根花卉之王[M].北京:金盾出版社,1999:34-35.
[2]刘小溪,吴丽芳,张艺萍,等.百合育种趋势及技术研究进展[J].浙江农业科学,2011,1(2):287-290.
[3]李守丽,石雷,张金政,等.百合育种研究进展[J].园艺学报,2006,33(1):203-210.
[4]王伟涛,杨炜茹,陈晶鑫,等.10种梅花品种及其3种近缘种的核型分析[J].西北植物学报,2013,33(11):2203-2208.
[5]李璇,段青,王祥宁,等.云南泸定百合12个野生居群的核型研究[J].园艺学报,2014,41(5):935-945.
[6]董山平,罗乐,钟军珺,等.榆叶梅及其部分近缘种的染色体核型分析[J].西北植物学报,2015,35(3):608-613.
[7]曾佳诗,王东旭,吴阳青,等.勋章菊品种核型分析[J].园艺学报,2015,42(12):2512-2518.
[8]戴小红,牛立新,张延龙.百合三品系代表品种的核型分析[J].西北林学院学报,2006,21(4):58-61.
[9]杨雪珍,贾月慧,张克中,等.部分中国野生百合的核型分析[J].西北植物学报,2013,33(5):922-930.
[10]席梦利,房磊,张晓玲,等.野百合与金佛山百合的核型分析及45S rDNA在其染色体上的物理定位[J].南京林业大学学报(自然科学版),2013,37(6):153-156.
[11]张艺萍,王继华,吴丽芳,等.百合属野生种和园艺品种的染色体核型[J].福建农林大学学报(自然科学版),2014,43(4):369-373.
[12]段青,崔光芬,贾文杰,等.云南泸定百合8个野生居群的染色体核型分析[J].西北植物学报,2016,36(3):472-478.
[13]李克虎,周桂雪,任贵玲,等.百合品种染色体倍性观察[J].园艺学报,2011,38(5):970-976.
[14]王中轩,魏迟,廉玉芹,等.中国原产4种野生百合的核型分析[J].园艺学报,2013,40(11):2207-2212.
[15]李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:1-37.
[16]Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J]. Hereditas,1964,52(2):201-220.
[17]Stebbins G L.Chromosome evolution in higher plants[M].London:Edward Amoldltd,1971:1-40.
[18]Arano H.The karyotypes and the speciation in subfamily carduoideae of Japan[J].Japanese Journal of Botany,1965:19(3):31-67.
[19]袁国良,徐萍,宫红霞,等.用三倍体作母本实现百合种质渗入育种[J].农业生物技术学报,2013,21(11):1321-1327.
[20]图力古尔,刘立波.吉林省5种百合的核型研究[J].植物科学学报,1996,14(1):6-12.
[21]Barba-Gonzalez R,Lim K-B,Ramanna M S.Occurrence of 2n gametes in the F1hybrids of Oriental×Asiatic lilies(Lilium):Relevance to intergenomic recombination and backcrossing[J].Journal of Euphytica,2005,143(1-2):67-73.
[22]洪德元.植物细胞分类学[M].北京:科学出版社,1990:91-96.
[23]魏迟,王中轩,贾桂霞.OT系百合3个品种的核型分析[J].东北林业大学学报,2013(11):59-62.
[24]曹钦政,贾桂霞,高雪,等.三倍体LA百合远缘杂交亲本的筛选[J].北京林业大学学报,2016,38(2):96-104.
[25]Schmitzer E.A survey of named polyploid lilies of the Asiatic section[J].Quarterly Bulletin of the North American Lily Society,1991,45:6-12.
[26]周桂雪,李克虎,张线线,等.亚洲百合品种倍性、花粉育性及其杂交研究[J].园艺学报,2011,38(4):733-739.
[27]李卫民,高英.中药百合的核型分析[J].中国中药杂志,1991,16(5):268-270.
