伐桩注水对毛竹光合蒸腾特性月变化的影响
2018-02-28耿养会谢锦忠
张 磊,杜 澜,童 龙,李 彬,耿养会,谢锦忠*,张 玮
(1.中国林业科学研究院 亚热带林业研究所,浙江 富阳 311400;2.重庆市林业科学研究院,重庆 400036)
光合特性是植物适应环境最重要的特征和物质积累代谢的基本单元,也是植物生长发育的基础和生产力高低的决定性因素[1-2]。光合作用不仅是植物在陆地生态系统吸收固定物质、分配转化及碳水循环的基础环节,同时也是植物生长和一切代谢活动的生理基础[3-4],对植物光合特性进行研究将为解释和预测植物内部与外部环境因子如何相互作用进而影响植物的生长、发育及物质生产过程中能量的吸收、固定、分配与转化起到重要作用。蒸腾作用通常用来反映了植物的水分状况及植物与环境因子之间的关系,是植物与自然界水分循环的主要环节[5]。植物的光合蒸腾作用是一个复杂的生物化学过程,受到很多因素的影响。它不仅受到植物自身遗传特性的影响,同时还受到外界环境因子的制约[6-8]。植物的光合蒸腾作用与土壤含水量之间存在着密切的关系,即使轻微的干旱也能使植物的光合蒸腾作用发生剧烈的变化。光合作用与灌水之间的关系一直是植物生理学、植物生态学的重要研究领域[9-11]。
毛竹(Phyllostachysedulis)隶属禾本科,竹亚科刚竹属植物。是我国竹类资源中栽培面积最大,经营历史最久,加工利用范围最广,对竹产区地方经济发展及竹农收入影响最为深远的集经济、生态、社会效益于一体的竹种,在集约经营条件下可获得较高的经济效益[12-13]。目前关于毛竹方面的研究多集中在不同年龄及不同生产力毛竹林的碳氮磷化学计量特征[14-15],毛竹不同基因的表达与功能及基因的结构与进化分析[16-18],集约经营模式下毛竹的空间分布格局[19],毛竹碳汇造林初期净碳汇量监测与不确定性分析[20]等,而对于不同灌溉条件下毛竹光合蒸腾特性的研究还相对较少,而且目前关于毛竹灌溉方面的研究又主要集中在滴灌和喷灌上,毛竹多生长在高山上,仅靠天然降水已不能满足毛竹的生产需求,而喷灌滴灌等灌溉方式又需要投入大量的人力财力。毛竹作为一种克隆植物,它能够通过地下鞭在克隆分株之间进行水分的运输与分享,从而实现毛竹的水分生理整合。因此本研究采用将毛竹伐桩内部的蜡质层和竹隔打破,并向伐桩中进行注水,使用注水伐桩对毛竹进行灌溉的方式来探讨不同数量注水伐桩对毛竹光合与蒸腾特性的影响及其与环境因子的关系,从而为毛竹林节水灌溉措施的制定和培育生产提供了理论参考。
1 研究区概况与研究方法
1.1 研究区概况
研究地点位于浙江省杭州市富阳区庙山坞自然保护区(30°03′~30°06′N,119°56′~120°02′E),该保护区属亚热带湿润季风气候,冬春季降雨较少。海拔为290~530 m,年平均降水量1 100 mm,年平均气温16.1 ℃,年平均无霜期237 d,土壤为微酸性红壤,总面积816.8 hm2,森林覆盖率93.6%。植物种类丰富,仅木本植物就有71科173属347种,野生植物大约1 000余种。主要植被类型包括天然次生林、毛竹林、人工杉木(Cunninghamialanceolata(Lamb.) Hook.)林和天然马尾松(PinusmassonianaLamb.)林等。
1.2 研究方法
1.2.1 试验设计 2016年1—12月,在试验毛竹林中选取立地条件、地形以及毛竹生长状况等基本相同的10 m×20 m样地9块,每块样地间隔5 m。为防止周围土壤中的水分通过水平方式输送到相邻样地,在每块样地四周挖深约50 cm、宽约20 cm的土沟;同时为了试验具有一致性,对每块样地中的毛竹进行砍伐,保证各块样地在砍伐后毛竹株数(3 000株/hm2)相同,砍伐后各试验样地立竹平均胸径分别为12.04,12.09,12.13,12.01,12.05,11.98,12.11,12.03,11.96 cm,对应的立竹年龄结构(3 a生∶2 a生∶1 a生)分别为1∶0.53∶1.24,1∶0.75∶1.35,1∶0.65∶1.36,1∶0.73∶1.35,1∶0.66∶1.31,1∶0.74∶1.26,1∶0.57∶1.47,1∶0.83∶1.52和1∶0.61∶1.33。试验设置CK(0个伐桩注水)、T1(12个伐桩注水)和T2(18个伐桩注水),每块样地内的注水伐桩散布尽量均匀,每处理3次重复,对每块相同处理样地设置相同的注水伐桩并用毛竹打桩机将伐桩内部的蜡质层和竹隔打破;伐桩按地径进行选取,且按1cm一个径阶进行划分,所选伐桩的径阶分别为10,11,12,13,14和15 cm;T1处理每个径阶的伐桩选2个,T2处理每个径阶的伐桩选3个,每个相同径阶的伐桩加水量控制相同(各径阶伐桩加水量分别为500,700,1 000,1 300,1 500,1 700 mL),其中每块样地在选择注水伐桩后,其余伐桩不处理,在2015年12月31日对T1和T2处理样地内已打通的伐桩统一进行加水,保证伐桩内始终有水,并将伐桩口用塑料薄膜包扎,以防止水分蒸发。
1.2.2 光合蒸腾特性和环境因子日变化的测定 试验于2016年1—12月,每月月底使用便携式光合仪(Li-Cor 6400,美国Li-Cor 公司)对2 a生标准竹叶片的净光合速率(Pn)与蒸腾速率(Tr)进行原位测定,光强均设为900 μmol/(m2·s)。试验在天气晴朗的白天09:00—11:00进行,每块样地选取标准竹3株,分别在各标准竹上部、中部和下部各选取3个叶片进行测定。由于毛竹的叶片不能充满叶室,试验结束后摘下所测叶片,用拍照法计算所测部分叶面积[21],随后对每个毛竹叶片的光合测定值进行调整。测定主要光合参数为净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)。林内环境因子为空气温度(Ta)、空气相对湿度(RH)、大气CO2浓度(Ca)。其中根据光合参数计算水分利用效率WUE=Pn/Tr和气孔限制值Ls=1-Ci/Ca。
1.2.3 数据处理方法 试验数据在Excel 2003统计软件和Origin 8.0制图软件中进行整理和图表制作,用SPSS 19.0软件进行单因素方差分析(One-way ANOVA)和新复极差法(Duncan)多重比较,设定P<0.05为显著性水平,比较不同处理毛竹各月之间光合蒸腾特性差异。试验中所有数据均采用均值±标准差来表示。
2 结果与分析
2.1 毛竹林内主要环境因子月变化
由图1可知,测定期间毛竹林内空气温度呈双峰值变化,从1月开始上升,直到8月达到第一个峰值,随后开始下降,在11月空气温度出现略微的回升后,又出现了下降,两次峰值分别出现在8月和11月,分别为34.53 ℃和24.24 ℃,空气温度的最低值出现在1月,为8.34 ℃,测定期间毛竹林内空气温度的年均值为23.08 ℃;测定期间毛竹林内的空气相对湿度和大气CO2浓度月变化均呈多峰曲线,毛竹林内的空气相对湿度1月开始下降,在4月出现一年中的最低值,为24.06%,随后又开始上升,6—12月的变化幅度相对较小,空气相对湿度的最大值出现在8月,为69.69%,测定期间毛竹林内空气相对湿度的年均值为53.65%;测定期间毛竹林内大气CO2浓度在1—10月不同月份之间的变化幅度相对较小,其全年的最小值和最大值分别出现在10月和11月,分别为406.23和457.22 μmol/mol,测定期间毛竹林内大气CO2浓度的年均值为422.68 μmol/mol。
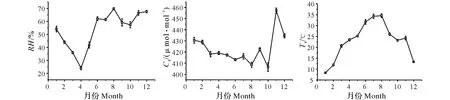
图1 毛竹林内主要环境因子月变化Fig.1 Monthly changes of main environmental factors in Phyllostachys edulis Forest
2.2 不同注水伐桩处理下毛竹净光合速率与蒸腾速率月变化
由图2可知,不同处理下毛竹净光合速率月变化趋势基本相同,均呈多峰曲线。3个处理毛竹净光合速率的4次峰值均分别出现在3月、6月、9月和12月,T2处理毛竹净光合速率在6月出现最大值为7.45 μmol/(m2·s),T1和CK处理毛竹净光合速率的最大值均出现在9月,其值分别为5.92 和4.95 μmol/(m2·s),各处理毛竹净光合速率均在4月、8月和10月出现较小值。不同处理间毛竹净光合速率全年均表现为T2>T1>CK,且T1和T2处理毛竹净光合速率全年均与CK处理达到差异显著水平(P<0.05)。T1、T2处理毛竹净光合速率与CK处理相比均在5月增幅最大,分别为73.66%、96.99%。CK、T1和T2处理毛竹净光合的均值分别为3.61、4.68和5.74 μmol/(m2·s),其均值表现为T2>T1>CK,T1和T2处理毛竹净光合速率均值分别比CK提高了29.64%和59.00%。不同处理间毛竹蒸腾速率全年表现为T2>T1>CK,T1、T2处理与CK处理全年基本均达到差异显著水平(P<0.05)。T2和CK处理毛竹蒸腾速率月变化趋势基本相同,均呈双峰曲线,而T1处理毛竹蒸腾速率月变化呈单峰曲线。T2和CK处理毛竹蒸腾速率的第一个峰值均出现在3月,在4月出现略微降低,随后呈直线上升,且在6月达到一年内的最大值,其最大值分别为3.61和2.43 mmol/(m2·s),6月后又均呈下降变化;T1处理毛竹月变化呈单峰曲线,且在6月出现一年最大值,为3.18 mmol/(m2·s)。其中T1和T2处理毛竹蒸腾速率与CK处理相比最大增幅均出现在5月,分别为70.72%和95.59%。CK、T1和T2处理下毛竹蒸腾速率的年均值分别为1.02、1.24和1.48 mmol/(m2·s),其均值表现为T2>T1>CK,T1和T2处理下毛竹蒸腾速率的年均值分别比CK处理提高了21.57%和45.10%。
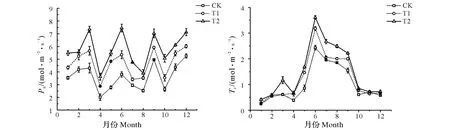
图2 不同注水伐桩处理下毛竹净光合速与蒸腾速率月变化Fig.2 Monthly variation of net photosynthesis and transpiration rate of Phyllostachys edulis with different number of water storage in stumps
2.3 不同注水伐桩处理下毛竹水分利用效率与气孔导度月变化
3个处理下毛竹水分利用效率月变化趋势基本相同,均呈不规则的反抛物线型(图3)。全年毛竹水分利用效率基本均表现为T1和T2处理略高于CK处理,这说明了,在毛竹林中增加注水伐桩在一定程度上可以提高毛竹水分利用效率。各处理毛竹水分利用效率均在1月出现最高值,且以T1处理的水分利用效率最高为15.70 μmol/mmol,随后各处理毛竹水分利用效率均出现降低,T1和CK处理毛竹水分利用效率最小值均出现在8月,其值分别为1.56和1.37 μmol/mmol,而T2处理毛竹水分利用效率最小值出现在7月,其值为1.66 μmol/mmol,各处理毛竹水分利用效率出现最低值后,均又出现小幅度回升,T1、T2处理毛竹水分利用效率与CK处理相比最大增幅分别出现在11月和7月,分别为39.29%和42.38%。CK、T1和T2处理下毛竹水分利用效率年均值分别为5.46、5.97和5.92 μmol/mmol,不同处理间毛竹水分利用效率年均值表现为T1>T2>CK,T1和T2处理下毛竹水分利用效率的年均值分别比CK提高了9.34%和8.42%。各处理毛竹气孔导度全年基本均表现为T1和T2处理高于CK处理,仅在2月和10月出现CK处理毛竹气孔导度分别略高于T2、T1处理的现象。CK、T1和T2处理毛竹全年气孔导度变化均呈多峰曲线,且最大值均出现在6月,分别为0.11、0.15和0.15 mol/(m2·s),最小值均出现在4月,分别为0.02、0.03和0.04 mol/(m2·s)。T1、T2处理毛竹气孔导度与CK处理相比,最大增幅分别出现在5月和10月,且分别增幅了179.96 %和124.07 %。CK、T1和T2处理下毛竹气孔导度的年均值分别为0.07、0.09 和0.10 mol/(m2·s),不同处理间毛竹气孔导度年均值表现为T2>T1>CK,T1和T2处理下毛竹气孔导度年均值分别比CK提高了28.57%和42.86%。
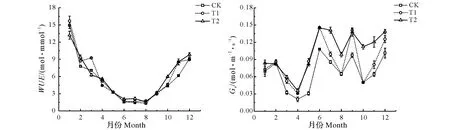
图3 不同注水伐桩处理下毛竹水分利用效率与气孔导度月变化Fig.3 Monthly variation of water use efficiency and stomatal conductance of Phyllostachys edulis with different number of water storage in stumps
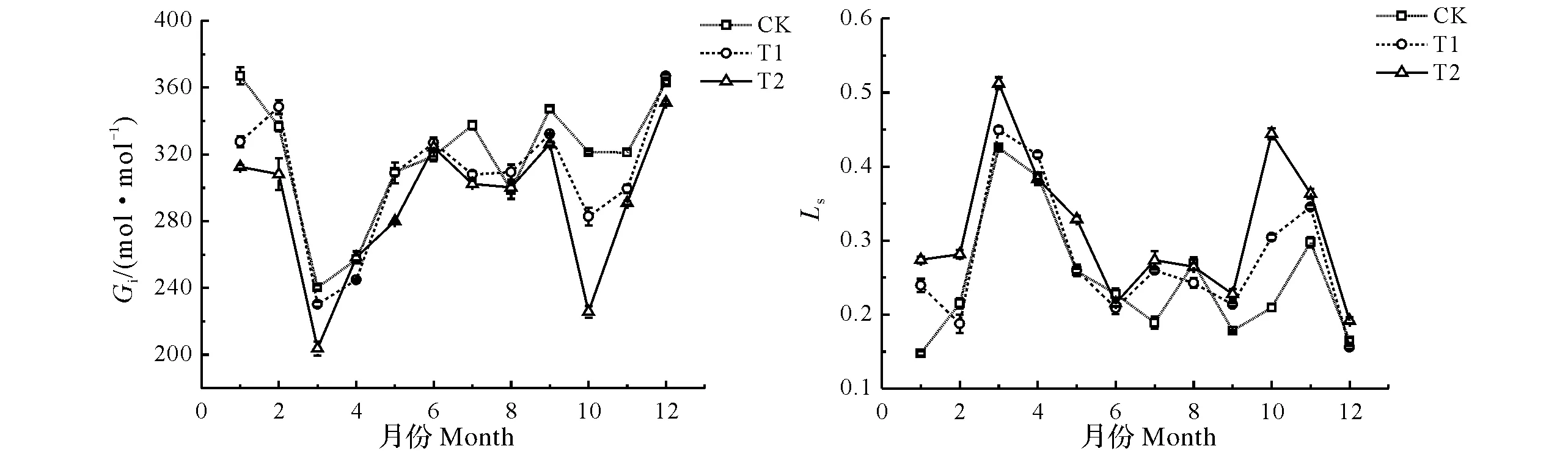
图4 不同注水伐桩处理下毛竹胞间CO2浓度与气孔限制值月变化Fig.4 Monthly variation of intercellular CO2 concentration and stomatal limitation of Phyllostachys edulis with different number of water storage in stumps
2.4 不同注水伐桩处理下毛竹胞间CO2浓度与气孔限制值月变化
由图4可知,3个处理下毛竹胞间CO2浓度与气孔限制值呈相反的变化趋势,且分别呈多谷和多峰曲线变化。不同处理间毛竹胞间CO2浓度全年基本表现为CK>T1>T2,CK处理毛竹胞间CO2浓度最大值出现在1月,其值为366.97 μmol/mol,而T1和T2处理毛竹胞间CO2浓度最大值均出现在12月,其值分别为366.87 和351.04 μmol/mol,CK、T1和T2处理毛竹胞间CO2浓度最小值均出现在3月,其值分别为240.33、230.27和203.87 μmol/mol,T1、T2处理毛竹胞间CO2浓度与CK处理相比,最大降幅均出现在10月,分别降低了12.02 %、29.73 %。CK、T1和T2处理下毛竹胞间CO2浓度年均值分别为318.14、307.10和290.19 μmol/mol,不同处理间毛竹胞间CO2浓度年均值表现为CK>T1>T2,T1和T2处理下毛竹胞间CO2浓度的年均值分别比CK处理降低了3.45%和8.79%。不同处理间毛竹气孔限制值全年基本表现为T2>T1>CK,CK、T1和T2处理毛竹气孔限制值的最大值均出现在3月,其值分别为0.43、0.45和0.51,而CK、T1和T2处理毛竹气孔限制值的最小值分别出现在1月、2月和6月,其值分别为0.15、0.19和0.22,T1、T2处理毛竹气孔限制值与CK处理相比,分别出现在1月和10月,分别增加了63.14 %、112.21 %。CK、T1和T2处理下毛竹气孔限制值年均值分别为0.25、0.27和0.31,不同处理间毛竹气孔限制值年均值表现为T2>T1>CK,T1和T2处理下毛竹蒸腾速率的年均值分别比CK升高了8.00%和24.00%。
2.5 不同注水伐桩处理下毛竹各指标与林内环境因子相关分析
由表1可知,CK处理毛竹净光合速率与大气CO2浓度显著正相关(P<0.05),与空气温度和空气相对湿度相关性不显著,CK处理下毛竹的蒸腾速率与大气温度极显著正相关(P<0.01),与空气相对湿度和与大气CO2浓度相关性不显著;T1和T2处理下毛竹净光合速率与各环境因子均相关均不显著,T1和T2处理下毛竹蒸腾速率均与大气CO2浓度显著正相关(P<0.05),与空气温度极显著正相关(P<0.01)。由此可知,在毛竹林中增加注水伐桩能够在一定程度上改变毛竹净光合速率和蒸腾速率与环境因子之间的相关关系。
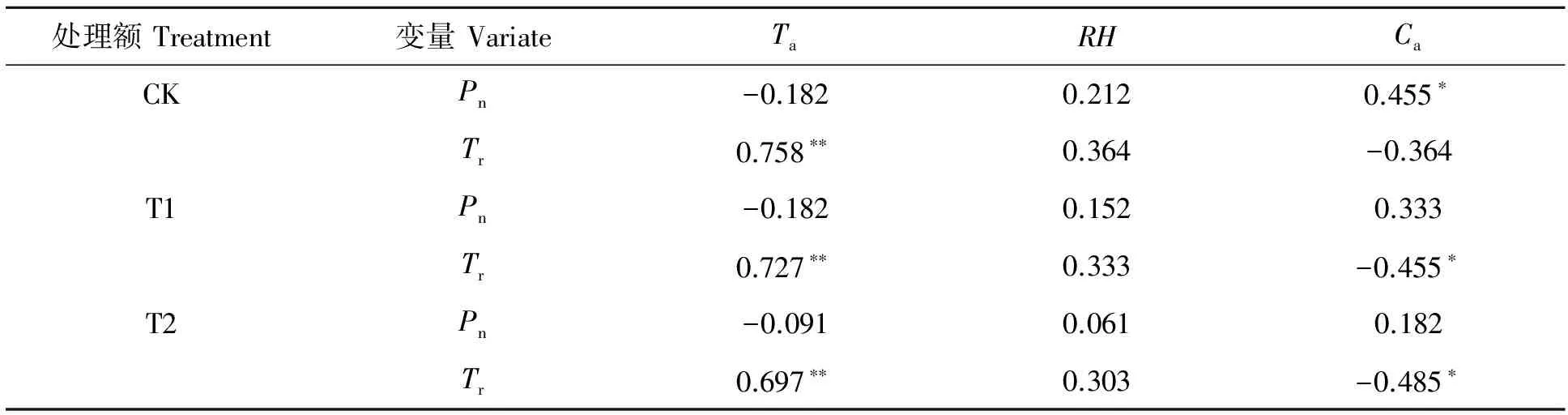
表1 不同注水伐桩处理毛竹净光合速率和蒸腾速率月变化与林内环境因子月变化相关分析
*表示在P<0.05 水平显著相关,**表示在P<0.01 水平显著相关。
* represents significant correlation at 0.05 level,** represents significant correlation at 0.01 leve.
3 讨论与结论
光合作用是一切植物生命活动的基础,同时光合作用与灌水量之间存在着密切的关系。相关研究显示,在模拟干旱环境下,毛竹净光合速率随注水伐桩数量的增加而增加[22]。本研究发现与此相同的结果,3个处理毛竹净光合速率全年的变化趋势基本相同,全年均表现为随着灌水量的增加(注水伐桩水量的增加),毛竹的净光合速率增加,T1和T2处理毛竹净光合速率全年均CK处理存在显著差异(P<0.05)。仍有研究表明,植物的净光合速率与灌水量之间并非总是存在着正相关关系,灌水量与植物净光合速率之间存在一个阈值,不同植物之间的阈值的大小也存在一定的差异,当灌水量未超过这个阈值时,植物的净光合速率与灌水量正相关,但当灌水量超过这个阈值,植物的净光合速率反而随着灌水量的增加而下降[23]。本研究不同处理下毛竹的净光合速率全年均表现为随着注水伐桩数量的增加而增加,未出现随着注水伐桩数量增加其净光合速率降低的现象,这可能是在增加18个注水伐桩的情况下,全年灌水量并未超过毛竹净光合速率与灌水量之间的这个阈值。因此,毛竹净光合速率与注水伐桩数量之间的关系有待进一步深入的研究,但在实际生产中,增加18个注水伐桩的处理已接近达到极限水平,因为在竹林抚育过程中单位面积采伐的老竹数量是有限度的。在本研究中T1、T2处理毛竹净光合速率与CK处理毛竹净光合速率相比均在5月增幅最大,说明在毛竹林中增加注水伐桩对毛竹5月分的光合产物的积累最为有利。
蒸腾是植物重要的生理活动之一[24]。植物蒸腾速率的大小与灌水量密切相关。在本研究中,毛竹蒸腾速率全年均表现为随着灌水量(注水伐桩数量)的增加而增加,这与张磊[25]和赵文赛[26]等在植物蒸腾速率与水分之间关系中的研究结果相同。而王慧[27]和李杨[28]等的研究发现,植物的蒸腾速率随着灌水量的增加呈先增加后降低的现象,其研究仍表明不同植物蒸腾速率响应灌水量降低的临界值存在差异。本研究中,毛竹的蒸腾速率全年均表现为随着注水伐桩的增加而增加,未出现灌水量与毛竹蒸腾速率呈负相关的现象,这可能是18个注水伐桩的灌水量还未超过能够引起毛竹蒸腾速率降低的范围。因此毛竹蒸腾速率与注水伐桩之间的关系还需要进一步研究。本研究发现,4—10月T1和T2处理毛竹蒸腾速率均与CK处理毛竹蒸腾速率间差异较大,且达到差异显著水平(P<0.05),其他月份不同处理间毛竹蒸腾速率差异较小,说明在毛竹林中增加注水伐桩对4—10月毛竹的蒸腾速率影响作用较大。
在本研究中,不同处理下毛竹水分利用效率月变化趋势基本相同,均呈不规则的反抛物线型,不同处理间毛竹的水分利用效率差异不大,且全年均基本以T2处理毛竹的水分利用效率较高,这说明了增加注水伐桩可以提高毛竹的水分利用效率。不同处理间毛竹气孔导度全年基本表现为T2>T1>CK,说明在毛竹林中增加注水伐桩后,可以使得毛竹的气孔导度增加。不同处理间毛竹胞间CO2浓度全年基本表现为CK>T1>T2,通过结合毛竹净光合速率的变化可以看出,T1和T2处理下毛竹胞间CO2的利用能力得到了增强,这可能是在增加注水伐桩后毛竹叶肉细胞的光合能力得到了改善,从而增加了对毛竹胞间CO2的利用。由各处理下毛竹全年气孔限制值的变化可知,CK处理毛竹气孔限制值全年均处于较低的水平,T1和T2处理全年均处于较高的水平,这可能是由于增加注水伐桩后改善了毛竹叶肉细胞的光合能力,表现为增强了对其胞间CO2的利用,使CO2处于相对较低的水平,从而使得气孔限制值较大。在本研究中,不同处理下毛竹的蒸腾速率均与空气温度极显著正相关(P<0.01),同时CK处理毛竹净光合速率及T1和T2处理毛竹蒸腾速率均与大气CO2浓度显著正相关(P<0.05)。在一年中大多环境因子的变化很难通过一定的措施进行改变如光合有效辐射、空气温度和空气相对湿度,但是大气CO2浓度可以通过一定的技术措施来加以控制,如在毛竹林中通过施用有机肥来促进CO2的生成,或者通过合理砍伐毛竹实现通风透光,均可以达到提高毛竹林内大气CO2浓度的目的,从而可以进一步提高毛竹的光合能力。
不同处理下毛竹的净光合速率、蒸腾速率、水分利用效率、气孔导度和气孔限制值全年均基本表现为T2>T1>CK;毛竹胞间CO2浓度全年基本均表现为CK>T1>T2。在毛竹林中增加注水伐桩能够在一定程度上改变环境因子与毛竹净光合速率与蒸腾速率之间的相关关系。增加注水伐桩能够显著提高毛竹的光合蒸腾能力,增加毛竹光合产物的积累,同时能够改善毛竹的生长状况,毛竹全年光合蒸腾特性与注水伐桩数量间的关系还有待进一步研究。
[1] Gago J,Coopman R E,Cabrera H M,et al.Photosynthesis limitations in three fern species[J].Physiologia Plantarum,2013,149(4):599-611.
[2] Collatz G J.Influence of certain environmental factors on photosynthesis and photorespiration inSimmondsiachinensis[J].Planta,1977,134(2):127-132.
[3] 韩忠明,王云贺,林红梅,等.吉林不同生境防风夏季光合特性[J].生态学报,2014,34(17):4874-4881.
Han Z M,Wang Y H,Lin H M,et al.Photosynthetic characteristics ofSaposhnikoviadivaricatain different habitats in summer[J].Acta Ecologica Sinica,2014,34(17):4874-4881.
[4] James B.Photosynthetic energy conversion:natural and artificial[J].The Royal Society of Chemistry,2009,38:185-196.
[5] 赵平,邹绿柳,饶兴权,等.成熟马占相思林的蒸腾耗水及年际变化[J].生态学报,2011,31(20):6038-6048.
Zhan P,Zou L L,Rao X Q,et al.Water consumption and annual variation of transpiration in matureAcaciamangiumPlantation[J].Acta Ecologica Sinica,2011,31(20):6038-6048.
[6] Lewis L D,Lucash M,Olszyk D,et al.Seasonal patterns of photosynthesisi in douglas fir seedlings during the third and fourth year of exposure to elevated CO2and temperature[J].Plant,Cell and Environment,2011,24(5):539-548.
[7] 张津林,张志强,查同刚,等.沙地杨树人工林生理生态特性[J].生态学报,2006,26(5):1523-1532.
Zhang J L,Zhang Z Q,Zha T G,et al.Physiological regulations of photosynthesis in a poplar plantation on a sandy soil[J].Acta Ecologica Sinica,2006,26(5):1523-1532.
[8] 靳甜甜,傅伯杰,刘国华,等.不同坡位沙棘光合日变化及其主要环境因子[J].生态学报,2011,31(7):1783-1793.
Jin T T,Fu B J,Liu G H,et al.Diurnal changes of photosynthetic characteristics ofHippophaerhamnoidesand the relevant environment factors at different slope locations[J].Acta Ecologica Sinica,2011,31(7):1783-1793.
[9] 冯强,胡聃,李娜,等.典型城区与郊区环境下大叶黄杨光合特性的比较[J].城市环境与城市生态,2008,21(2):13-16.
Feng Q,Hu D,Li N,et al.Comparative analysis photosynthetio characteristics ofEuymusjapinicusthumb in typical urban and suburban environment[J].Urban environment and urban ecology,2008,21(2):13-16.
[10] Xu D Q,Shen Y G.Diurnal variations in the photosynthetic efficiency in plants[J].Acta Phytophysiologica Sinaca,1997,23(4):410-416.
[11] 王孟本,李洪建,柴宝峰,等.树种蒸腾作用、光合作用和蒸腾效率的比较研究[J].植物生态学报,1999,23(5):401-410.
Wang M B,Li H J,Cai B F,et al.A comparision of transpiration,photosynthesis and trabspiration efficiency in four tree species in the loess region[J].Acta Phytoecologica Sinica,1999,23(5):401-410.
[12] 金爱武.现代毛竹培育技术及其传播:问题和方法[M].北京:中国农业出版社,2006.
Jin I W.Modern bamboo breeding technology and its propagation:problems and methods[M].Beijing:China Agriculture Press,2006.
[13] 应叶青,郭璟,魏建芬,等.水分胁迫下毛竹幼苗光合及叶绿素荧光特性的响应[J].北京林业大学学报,2009,31(6):128-133.
Ying Y Q,Guo J,Wei J F,et al.Photosynthetic and chlorophyll fluorescent responses ofPhyllostachyspubescensseedlings to water deficiency stress[J].Journal of Beijing Forestry University,2009,31(6):128-133.
[14] 郭宝华,刘广路,范少辉,等.不同生产力水平毛竹林碳氮磷的分布格局和计量特征[J].林业科学,2014,50(6):1-9.
Guo B H,Liu G L,Fan S H,et al.Distribution patterns and stoichiometry characteristics of C,N,P inPhyllostachysedulisforests of different productivity levels[J].Scientia Silvae Sinicae,2014,50(6):1-9.
[15] 范少辉,刘广路,郭宝华,等.不同年龄毛竹碳氮磷化学计量特征[J].热带作物学报,2016,37(2):279-285.
Fan S H,Liu G L,Guo B H,et al.The carbon,nitrogen and phosphorus contents ofPhyllostachyseduliswith different ages[J].Chinese Journal of Tropical Crops,2016,37(2):279-285.
[16] 张庆波,胡尚连,徐刚,等.毛竹bZIP转录因子的基因结构与进化分析[J].森林与环境学报,2016,57(1):54-61.
Zhang Q B,Hu S L,Xu G,et al.Analysis on gene structure and evolution of bZIP transcription factor ofPhyllostachysedulis[J].Journal of Forest and Environment,2016,57(1):54-61.
[17] 孙化雨,娄永峰,李利超,等.毛竹TIPs基因家族成员组织表达模式研究[J].林业科学研究,2016,29(4):521-528.
Sun H Y,Lou Y F,Li L C,et al.Tissue expression pattern analysis of TIPs genes inPhyllostachysedulis[J].Forest Research,2016,29(4):521-528.
[18] 董丽莉,赵韩生,王丽丽,等.毛竹PeSCR基因的表达与功能[J].林业科学,2016,52(6):35-42.
Dong L L,Zhao H S,Wang L L,et al.Expression and function of PeSCR gene fromPhyllostachysedulis[J].Forest Research,2016,52(6):35-42.
[19] 顾琪,陈霜霜,彭悦,等.集约经营模式下毛竹的空间分布格局[J].南京林业大学学报(自然科学版),2016,40(1):162-168.
Gu Q,Chen S S,Peng Y,et al.Spatial distribution pattern ofPhyllostachysedulisunder the pattern of intensive faring[J].Journal of Nanjing Forestry University(Natural Sciences),2016,40(1):162-168.
[20] 俞淑红,周国模,施拥军,等.毛竹碳汇造林初期净碳汇量监测与不确定性分析[J].浙江农林大学学报,2016,33(5):807-815.
Yu S H,Zhou G M,Shi Y J,et al.Net carbon sinks in the initial stages of Moso bamboo stands[J].Journal of Zhejiang A& F University,2016,33(5):807-815.
[21] 王忠芝,张金瑞.基于图像处理的叶面积测量方法[J].微计算机应用,2010,31(5):68-72.
Wang Z Z,Zhang J R.A measurement approach of leaf area based on digital image processing[J].Microcomputer Applications,2010,31(5):6872.
[22] 张磊,谢锦忠,张玮,等.模拟干旱环境下伐桩注水对毛竹光合蒸腾特性的影响[J].南京林业大学学报(自然科学版),2017,41(2):43-51.
Zhang L,Xie J Z,Zhang W,et al.Effects of stump water storage on the photosynthesis and transpiration characteristics ofPhyllostachysheterocyclacv.Pubescensunder a simulated drought environment[J].Journal of Nanjing Forestry University(Natural Sciences),2017,41(2):43-51.
[23] 夏江宝,张光灿,孙景宽,等.山杏叶片光合生理参数对土壤水分和光照强度的阈值效应[J].植物生态学报,2011,35(3):322-329.
Xia J B,Zhang G C,Sun J K,et al.Threshold effects of photosynthetic and physiological parameters inPrunussibiricato soil moisture and light intensity[J].Chinese Journal of Plant Ecology,2011,35(3):322-329.
[24] Jones H G.Plants and Microclimate.2nd edn[M].Cambridge,UK.Cambridge University Press,1983.
[25] 张磊,谢锦忠,张玮,等.伐桩储水下毛竹光合蒸腾特性[J].生态学杂志,2017,36(3):655-663.
Zhang L,Xie J Z,Zhang W,et al.Photosynthesis and transpiration characteristics ofPhyllostachysedulisstands with water storage in their stumps[J].Chinese Journal of Ecology,2017,36(3):655-663.
[26] 赵文赛,孙永林,刘西平.干旱-复水-再干旱处理对玉米光合能力和生长的影响[J].植物生态学报,2016,40(6):594-603.
Zhao W S,Sun Y L,Liu X P.Effects of drought-rewatering-drought on photosynthesis and growth of maize[J].Chinese Journal of Plant Ecology,2016,40(6):594-603.
[27] 王慧,周广胜,蒋延玲,等.降水与CO2浓度协同作用对短花针茅光合特性的影响[J].植物生态学报,2012,36(7):597-606.
Wang H,Zhou G S,Jiang Y L,et al.Interactive effects of changing precipitation and elevated CO2concentration on photosynthetic parameters ofStipabreviflora[J].Chinese Journal of Plant Ecology,2012,36(7):597-606.
[28] 李杨,黄建辉.库布齐沙漠中甘草对不同水分和养分供应的光合生理响应[J].植物生态学报,2009,33(6):1112-1124.
Li Y,Huang J H.Photosynthetic physiological responses of glycyrrhiza uralensis under different water and nutrient supplies in Kubuqi Desert,China[J].Chinese Journal of Plant Ecology,2009,33(6):1112-1124.
