水稻籼粳指数鉴定方法及亚种间杂交的分子辅助育种
2018-02-27季文杰李阳生
季文杰 李阳生


摘要:籼粳交F1株高、生育期超亲、结实率低、子粒充实度差是长期困扰水稻(Oryza sativa L.)亚种间杂交育种的几大难题。通过对水稻籼粳指数鉴定方法的发展、遗传距离与杂种优势关系的研究进行总结,并结合株高、抽穗期、育性基因及库源关系等方面的研究进展,讨论分子标记方法在水稻育种工作中的应用前景,以期为水稻籼粳亚种间杂种优势利用提供一定的参考价值。
关键词:水稻(Oryza sativa L.);亞种间杂交;籼粳指数;遗传距离(GD);分子标记辅助选择(MAS)
中图分类号:S512 文献标识码:A
文章编号:0439-8114(2018)24-0041-05
DOI:10.14088/j.cnki.issn0439-8114.2018.24.011 开放科学(资源服务)标识码(OSID):
Abstract: Tall culm,long growth period,low seed setting rate and bad grain filling are the main problems of indica-japonica inter-subspecific hybrid F1 progeny in rice.This article reviewed the advances of indica-japonica index identification technologies,the relationship between parents genetic distance and F1 heterosis,and the progress in research on plant hight,heading date,fertility genes,and source-sink relationship,to investigate the prospect of molecular markers in rice breeding,hoping to provide some references for the utilization of inter-subspecific heterosis.
Key words: rice(Oryza sativa L.); inter-subspecific hybridization; indica-japonica index; genetic distance (GD); molecular marker-assisted selection (MAS)
水稻(Oryza sativa L.)籼粳亚种间杂交有着显著的杂种优势效应,一直以来都是育种家关注的重点,然而普遍偏低的结实率成为限制其超高产潜力实现的主要因素之一。前人研究[1,2]表明,杂交水稻株高优势遵循籼粳交>籼爪交>粳爪交>籼籼交>粳粳交的规律,而结实率趋势则正相反。如何掌握其中的平衡,仍需要大量的尝试和摸索,而在此之前,鉴定试验材料的籼粳属性是非常必要的。典型籼粳交组合的F1通常存在株高偏高、生育期偏长、结实率较低、谷粒充实度较差等问题。在此,结合最新的分子标记技术和量化研究方法,明确材料籼粳指数,优化亲本遗传距离,将有利于选育出优良的亚种间杂交稻组合。
1 籼粳指数鉴定方法
1.1 形态标记
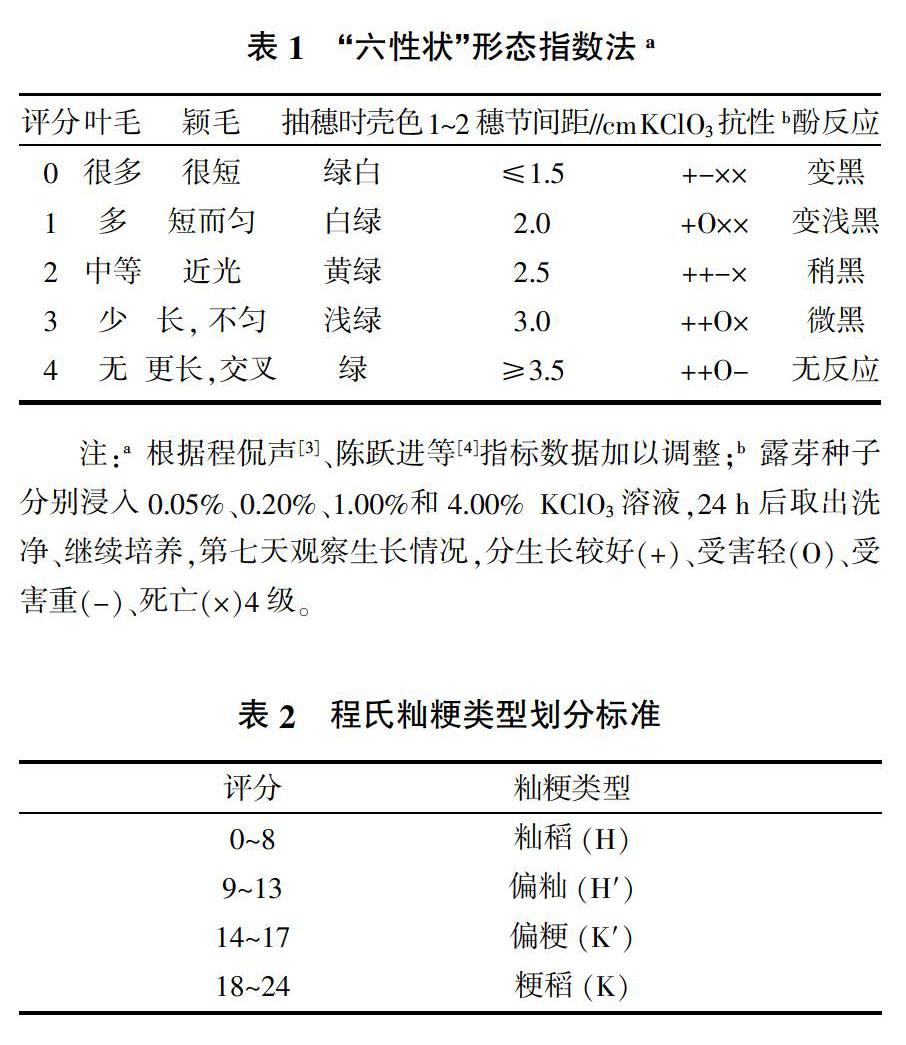
对于田间应用,中国主要采用程氏(六性状)指数法[3];后由陈跃进等[4]改进,增加幼苗KClO3抗性指标(表1、表2)。
1.2 生理生化标记
主要有同工酶法和维管束法。同工酶法,常用酯酶,已鉴别并应用了10多个籼粳特异酯酶同工酶带[5];维管束法,籼稻/粳稻穗颈大小维管束之比分别在1.0和0.5左右,倒二节间和穗颈大维管束之比分别在1.5和2.5左右[6]。
1.3 分子标记
从最开始应用化学酶切方法的RFLP,到酶切与PCR相结合的AFLP、RAPD,及至后期SSR、ISSR、EST、CAPs、InDel、ILP、SRAP、TRAP、SNP等各种PCR标记[7]。常运用于水稻籼粳指数检测的大致有以下几种。
1.3.1 限制性片段长度多态性(RFLP) RFLP是最早采用的一类分子标记方法。李任华等[8]采用42对籼粳特异RFLP探针,结合数学方法鉴定籼粳指数,计算公式为:
式中,Dj表示单位点粳性指数,Fi、Fj分别是此位点某等位基因在籼、粳类群中的基因频率。
式中,TDj表示品种总粳性指数。
此方法采用酶切手段,存在技术繁琐、多态性低等缺点,后续出现了RAPD、AFLP等改进。
1.3.2 简单序列重复(SSR) 又称STR、SSLP、微卫星DNA标记。毛艇等[9]以典型籼稻七山占和典型粳稻秋光及其RILs为对象,以28对籼粳特异SSR标记可以很好地鉴定材料的籼粳特性。计算公式为:
Dj=B位点数/(A位点数+B位点数)×100%
式中,Dj表示偏粳系数,A、B分别表示与七山占或秋光带型相同的位点。并定义Dj>50%为粳型,Dj<50%为籼型。
SSR标记因其共显性、高信息量与可重复性以及较低的技术难度和试验成本得到了很多研究者的青睐,经过多次改进之后,已成为目前最常用的水稻籼粳特性鉴定方法之一。
1.3.3 插入/缺失标记(InDel) Lu等[10]利用基于9311(典型籼稻)和日本晴(典型粳稻)全基因组序列比对获得的34对籼粳特异性InDel标记进行水稻籼粳指数研究(InDel分子指数法)。计算方法为:
Fi、Fj分别表示品种籼性指数和粳性指数;Xii、Xjj、Xij各代表纯合籼、粳以及杂合型位点,是计为1,否计为0。
根据计算得到的Fi、Fj值划分籼粳属性,其分类标准见表3。此方法操作简单、重复性高,并且计算方式科学、籼粳分类细致,是一种较有前景的籼粳指数鉴定手段。
1.3.4 单核苷酸多态性(SNP) 随着测序技术的普及,获得籼、粳稻全基因组序列变得越来越容易,通过全基因组范围比对通常可以发现数以百万计的单碱基替换(SNP),是目前密度最大、应用范围最广的分子标记。SNP标记可结合基因芯片、测序技术,运用生物信息学手段实现数字化分析,这些标记在分子辅助育种、基因克隆工作中很有应用潜力[11],必将成为未来最有希望的分子标记及籼粳指数鉴定方法之一。
1.3.5 细胞质类型鉴定 有研究[12]表明,与粳稻相比,多数籼稻品种的叶绿体DNA(cpDNA)在ORF100内有69 bp缺失,可作为籼粳叶绿体分型的标记;孙传清等[13]使用17种内切酶组合的RFLP标记将118份普通野生稻和76份亚洲栽培稻线粒体DNA(mtDNA)分成5类,并证实籼粳分化依然是其中的主流。
以上结果表明,除核基因组以外,水稻在细胞质水平上同样存在着籼粳分化,为研究亚洲栽培稻的起源、亚种间杂交工作提供了新的思路。
2 遗传距离与杂种优势
有关亲本分化远近或称遗传距离(Genetic distance,GD)与杂种优势大小关系的研究很早就已经开始了。对籼型杂交稻的标记分析[14]表明,其亲本遗传距离狭窄(0.059 8~0.391 3,平均0.2左右),而汕优63所表现出的强大优势很可能起因于其亲本明恢63带有部分粳型血缘、双亲间GD较大,选择遗传关系较远的品种进行杂交显得尤为必要。甬优系列超级稻的开创者通过对甬优6号亲本的分析[15]发现,母本含有89.5%粳稻组分、父本含有84.2%籼稻组分,证明籼粳成分按一定比例配组可以克服F1育性障碍,继而育成高产超级杂种。
另一方面,有很多研究者一直在致力于寻找籼粳亲本最优遗传距离,并希望给出一个数量范围。李亚莉等[16]利用SSR标记对滇型杂交稻的分析认为,亲本遗传距离与杂种产量优势无线性关系,但适中的差异容易产生正向极端组合,便于筛选强优势品种。严钦泉等[17]以程氏指数法为标准,构建包含15个不同籼粳指数父母本的不完全双列杂交群组,结果显示,亲本程氏指数差值为9时(典型粳稻满分24),产量超亲优势有一峰值。Li等[18]使用42对RFLP探针,以培矮64s为母本(夏士健等[19]确定其籼性指数为0.76),认为父本总粳性指数在20~25(换算为百分值50%~60%)时,F1结实率与产量优势出现极大值。Dan等[20]采用34对InDel分子标记,检测并选出12种不同来源的籼粳稻亲本,组建完全双列杂交群组,结合表型分析数据,认为当双亲遗传分化指数(GDI)在0.37左右时,F1单株产量杂种优势最高。于亚辉等[21]选取142对SSR标记引物,用一套籼粳RILs与粳型两用不育系GB028S(Dj=0.875)进行杂交,对亲本偏粳系数及杂种表型进行分析,结果表明父本(RILs)Dj在0.55~0.65时,F1有形成较高产量优势的潜力。以上研究中F1出现产量优势峰值的双亲遗传距离均分布在0.3~0.4(略有一些浮动),在育种实践中或可借鉴。
当然,也有一些研究者持有不同观点。分子标记可分为一般性标记(随机标记)和特异性标记(与产量等QTL连锁)。Zhang等[22]根据混合标记的分析结果,认为F1杂种优势与标记的一般杂合度相关性不显著,与特殊杂合度相关显著;倪先林等[23]的结论则正相反。罗小金等[24]研究显示,不同亲本材料、遗传距离范围内,GD与F1单株产量及产量相关性状的杂种优势关联性各不相同,关系复杂,缺乏可统一的模式。
籼粳交亲本遗传距离与F1产量优势的平衡是一个十分复杂的问题,值得投入更多人力物力进行研究。同时,也可以利用已有成果进行一些尝试,大胆假设,小心求证。
3 分子标记辅助育种
典型籼粳亚种间杂交存在四大难题,针对这几个方面已经逐渐形成一些技术思路,而分子标记的发展则提供了更多选择。
3.1 株高偏高与矮秆基因的克隆
水稻株高的建成主要與赤霉素(GA)、油菜素内酯(BR)、独脚金内酯(SL)等植物激素有关,形成一个互作网络[25]。水稻中第一个被克隆的株高控制基因是“绿色革命”基因sd1[26],编码赤霉素GA20氧化酶,与另一个主效基因D18[27]的产物GA3氧化酶,是水稻株高调控的两个主要靶标。目前水稻中克隆赤霉素通路基因近20个,加上其他相关的生长因子通路,则克隆株高调控基因已达70余个[28],而理想株型基因IPA1[29]同样被证明会影响株高。已经有人尝试设计并应用了sd1位点的分子标记[30]对其他非等位矮秆基因进行选择和聚合,相信将会越来越多应用于籼粳交品种的株高控制。
3.2 生育期偏长与抽穗期相关基因
水稻的生育期,即从播种到成熟的整个生育进程,可以分为营养生长期、生殖生长期、成熟期3个阶段,其中生殖生长期和成熟期时间比较固定,受光温等环境因素影响较小。因此,对水稻生育期控制的研究大多集中在播种到始穗之前,即所谓抽穗期(Heading date)阶段。
水稻中有3条独立的光周期调控路径,分别为OsGI-Hd1-Hd3a[31]、Ghd7-Ehd1-Hd3a/RFT1[32]、DTH2-Hd3a/RFT1[33],感知日照长短对开花时间进行着精细的调控。关于籼粳交F1生育期超亲的问题,很多研究都提到了可以利用不同生态类型水稻的感光等位基因来控制,部分育种家已经做出了有益的尝试[34]。据吴子帅[35]总结,截至2016年,已克隆水稻抽穗期调控基因24个、定位QTL逾700个。结合分子标记辅助选择(MAS)和遗传工程手段对这些基因位点进行合理搭配与运用是未来的主要研究方向之一。
3.3 结实率低与广亲和材料的应用
水稻亚种间杂交F1结实率低起因于杂种不育现象,而以雌/雄配子败育最为常见。水稻中第一个定位的杂种不育基因是广亲和基因S5n[36],之后一系列育性相关位点陆续被报道,其中部分基因已被克隆并得到了较为深入的研究[37]。
关于如何利用亲和基因提高籼粳杂交F1结实率,一般有3种策略。第一种是培育含有S5n(及其他中性基因位点)的广亲和不育系、恢复系,与高配合力籼/粳品种杂交,得到产量性状优良的杂种后代。其典型应用首推以两优培九为代表的两系超级杂交稻(分别以培矮64s、Y58s等优秀广亲和不育系为母本)[38],以及采用“粳不籼恢”育种策略的优良三系杂交稻组合,包括甬优系列、浙优系列等(使用具有广亲和特性的籼粳中间型恢复系作为父本)[39]。第二种是通过导入Si或Sj基因培育粳型亲籼系或者籼型亲粳系,这样育成的品种如育种常用广亲和不育系509s[40]。第三种是利用RNAi技术抑制杂种不育相关基因的表达。
3.4 子粒充实度差与“源-库-流”关系理论
对水稻子粒充实过程的研究一般分为3个部分,分别为干物质生产能力、收获潜力及实际产量,即“源-库-流”的关系。籼粳杂交多数源、库比较充足,抽穗期物质转运障碍是导致充实度差的主要原因[41]。
对子粒灌浆与内源激素关系的研究发现,籼粳交水稻存在强烈的穗尖顶端优势,中下部穗粒发育受到抑制,呈现阶梯式灌浆现象;对子粒灌浆化学基础的研究发现,籼、粳基因的不协调性(相对于亲和性而言)导致杂种小穗中内源激素、ATP及淀粉合成酶等含量少且活性低,即子粒生理活性的下降,使得淀粉合成缓慢、胚乳发育不良,是引起穗部物质转入率低、谷粒充实度差的最主要原因[42]。关于育种,根据袁隆平“以饱攻饱”的原则[43],应当选择子粒充实度高、一般配合力好的亲本配组;同时,对于子粒充实性相关互补基因的发掘和利用仍是最有前途的方向之一。
4 展望
籼粳亚种间超高杂种优势的利用一直以来都是育种家努力追求的方向,但由于F1结实率低以及其他一些问题使得典型籼粳交杂种优势的直接利用难以实现。中国水稻学界先后提出部分利用籼粳有利性状[44]、籼粳架桥[45]、利用广亲和基因等多种方案,而对籼粳交亲本遗传距离与F1产量杂种优势关系的研究也是其中比较有希望的方向之一。根据对前人研究的总结,作者在此提出一个0.3~0.4的双亲最优遗传距离范围,以供各位指正与讨论,并希望为今后的籼粳杂交育种亲本选择工作提供一些借鉴。
在籼粳指数鉴定过程中应用了多种分子标记方法,结合对水稻株高、抽穗期、育性、灌浆活化基因的克隆与深入研究,利用分子标记辅助选择(MAS)方法,相信亚种间杂种优势利用的障碍将会逐步解决。
随着水稻基因组和生物信息学技术的快速发展,正进入一个数字化分析的全新时代。利用测序数据,一些软件已经可以做到初步籼粳聚类[46-48],另有一些研究则开始尝试应用程序手段开发和优化分子标记[49,50];下一步要做的就是解放双手,使用计算机代替PCR和电泳,直接实现全基因组范围内分子标记基因型的自动化分析,快速、准确、高效地鉴定材料籼粳指数,并结合最新的基因工程技术,实现水稻亚种间甚至是种间杂种优势的直接利用。
参考文献:
[1] 袁隆平.超级杂交水稻育种研究的进展[J].致富天地,2008,6(1):1-3.
[2] 肖国樱,袁隆平.水稻籼爪交和粳爪交杂种优势的研究Ⅰ.杂种农艺性状的表现[J].作物学报,2003,29(2):169-174.
[3] 程侃声.亚洲稻籼粳亚种的鉴别[M].昆明:云南科技出版社,1993.56-69.
[4] 陈跃进,丁效华,杨长寿,等.水稻粳型亲籼系籼粳型属性的程氏指数鉴定[J].湖南农业大学学报(自然科学版),2002,28(4):284-286.
[5] 段中岗,梁程邺.改良聚丙烯酰胺凝胶电泳法测定籼粳分化的同工酶带[J].兰州大学学报(自然科学版),2005,41(4):30-33.
[6] 徐正进,陈温福,张龙步,等.水稻穗颈维管束性状的类型间差异及其遗传的研究[J].作物学报,1996,22(2):l67-172.
[7] 许旭明.水稻籼粳亚种间杂交衍生系遗传基础的研究[D].福州:福建农林大学,2009.
[8] 李任华,徐才国,何予卿,等.水稻亲本遗传分化程度与籼粳杂种优势的关系[J].作物学报,1998,24(5):564-576.
[9] 毛 艇,徐 海,郭艳华,等.利用SSR分子标记进行水稻籼粳分类体系的初步构建[J].华北农学报,2009,24(1):119-124.
[10] LU B R,CAI X X. Efficient indica and japonica rice identification based on the InDel molecular method:Its implication in rice breeding and evolutionary research[J].Prog Nat Sci,2009, 19(10):1241-1252.
[11] 李 筠.SNP分子標记及其在水稻研究中的应用[J].农业与技术,2016,36(7):32-33.
[12] KANNO A,WATANABE N,NAKAMURA I,et al. Variation in chloroplast DNA from rice(Oryza sativa):Differences between deletions mediated by short direct-repeat sequences within a single species[J].Theor Appl Genet,1993,86(5):579-584.
[13] 孫传清,王象坤,吉村淳,等.普通野生稻和亚洲栽培稻线粒体DNA的RFLP分析[J].遗传学报,1998,25(1):40-50.
[14] 唐 梅.籼型杂交水稻杂种优势群的RAPD和AFLP分析[J].乐山师范学院学报,2005,20(5):59-61.
[15] 马荣荣,许德海,王晓燕,等.籼粳亚种间杂交稻甬优6号超高产株形特征与竞争优势分析[J].中国水稻科学,2007,21(3):281-286.
[16] 李亚莉,马继武,杨晓曦,等.利用SSR分析滇型杂交稻亲本籼粳分化与杂种优势的关系[J].西南农业学报,2012,25(2):347-353.
[17] 严钦泉,阳菊华,伏 军.两系杂交稻亲本籼粳程度与配合力及杂种优势的关系[J].湖南农业大学学报,2001,27(3):163-166.
[18] LI R,XU C G,YANG Z Y,et al. The extent of parental genotypic divergence determines maximal heterosis by increasing fertility in inter-subspecific hybrids of rice(Oryza sativa L.)[J].Molecular Breeding,1998,4(3):205-214.
[19] 夏士健,张启军,杨 杰,等.水稻不育系的广亲和基因检测及籼粳型分析[J].杂交水稻,2010,27(S1):698-703.
[20] DAN Z W,LIU P,HUANG W C,et al. Balance between a higher degree of heterosis and increased reproductive isolation:A strategic design for breeding inter-subspecific hybrid rice[J].PLoS ONE, 2014,9(3):e93122.
[21] 于亚辉,刘 郁,李振宇,等.亲本籼粳成分与两系杂交粳稻杂种优势的关系及遗传基础[J].作物学报,2016,42(5):648-657.
[22] ZHANG Q F,GAO Y J,SAGHAI MAROOF M A,et al. Molecular divergence and hybrid performance in rice[J].Molecular Breeding,1995,1(2):133-142.
[23] 倪先林,张 涛,蒋开锋,等.SSR分子标记与水稻杂种优势的相关性研究[J].安徽农业科学,2009,37(23):10913-10916.
[24] 罗小金,贺浩华,彭小松,等.利用SSR标记分析水稻亲本间遗传距离与杂种优势的关系[J].植物遗传资源学报,2006,7(2):209-214.
[25] WANG Y,ZHAO J,LU W,et al. Gibberellin in plant height control:Old player,new story[J].Plant Cell Reports,2017,36(3):391-398.
[26] ASHIKARI M,SASAKI A,UEGUCHI-TANAKA M,et al. Loss-of-function of a rice gibberellin biosynthetic gene,GA20 oxidase(GA20ox-2),led to the rice ‘Green Revolution[J].Breeding Science,2002,52(2):143-150.
[27] ITOH H, UEGUCHI-TANAKA M,SAKAMOTO T,et al. Modification of rice plant height by suppressing the height-controlling gene,D18,in rice[J].Breeding Science,2002,52(3):215-218.
[28] LIU F,WANG P,ZHANG X,et al. The genetic and molecular basis of crop height based on a rice model[J].Planta,2017, 247(1):1-26.
[29] JIAO Y,WANG Y D,WANG J,et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J].Nature Genetics,2010,42(6):541-544.
[30] 徐建军,张云辉,顾 啸,等.水稻半矮秆基因sd1功能标记的开发与利用[J].上海农业学报,2016,32(4):22-24.
[31] TSUJI H,TAOKA K I,SHIMAMOTO K. Florigen in rice:Complex gene network for florigen transcription,florigen activation complex,and multiple functions[J].Curr Opin Plant Biol,2013, 16(2):228-235.
[32] SONG Y H,SHIM J S,KINMONTHSCHULTZ H A,et al. Photoperiodic flowering:Time measurement mechanisms in leaves[J].Annu Rev Plant Biol,2015,66(1):441-464.
[33] WU W,ZHENG X M,LU G,et al. Association of functional nucleotide polymorphisms at DTH2 with the northward expansion of rice cultivation in Asia[J].PNAS,2013,110(8):2775-2780.
[34] 杜雪树,戚华雄,廖世勇,等.水稻抽穗期分子生物学研究进展[J].湖北农业科学,2013,52(24):5958-5962.
[35] 吴子帅.水稻早抽穗特性的遗传分析及QTL初步定位[D].南宁:广西大学,2016.
[36] YANG J Y,ZHAO X B,CHENG K,et al. A killer-protector system regulates both hybrid sterility and segregation distortion in rice[J].Science,2012,337(6100):1336-1340.
[37] 欧阳亦聃.水稻籼粳杂种不育与广亲和[J].科学通报,2016, 61(35):3833-3841.
[38] 吴 俊,邓启云,袁定阳,等.超级杂交稻研究进展[J].科学通报,2016,61(35):3787-3796.
[39] 宋昕蔚,林建荣,吴明国. 水稻籼粳亚种间杂种优势利用研究进展与展望[J].科学通报,2016,61(35):3778-3786.
[40] CHEN L,ZHAO Z, XI L,et al. Marker-assisted breeding of a photoperiod-sensitive male sterile japonica rice with high cross-compatibility with indica rice[J].Molecular Breeding,2011, 27(2):247-258.
[41] 朱庆森,张祖建,杨建昌,等.亚种间杂交稻产量源库特征[J].中国农业科学,1997,30(3):52-59.
[42] 陶龙兴,王 熹.亚种间杂交稻结实障碍的生理学研究进展[J].中国农业科技导报,2002(1):54-59.
[43] 袁隆平.选育水稻亚种间杂交组合的策略[J].杂交水稻,1996(2):1-3.
[44] 杨守仁.籼粳稻杂交育种研究——二十年来的经验体会提要[J].遗传学通讯,1973(2):34-38.
[45] 杨振玉,张忠旭,華泽田,等.不同类型籼粳亚种间杂种F1可利用和非可利用杂种优势的评价利用[J].中国水稻科学,1991, 4(2):49-55.
[46] PRICE A L,PATTERSON N J,PLENGE R M,et al. Principal components analysis corrects for stratification in genome-wide association studies[J].Nature Genetics,2006,38(8):904-909.
[47] TAMURA K,PETERSON D,PETERSON N,et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Mol Biol Evol,2011,28(10):2731-2739.
[48] RAJ A,STEPHENS M,PRITCHARD J K. fastSTRUCTURE:variational inference of population structure in large SNP data sets[J].Genetics,2014,197(2):573-589.
[49] GUPTA S,BHARALEE R,DAS R,et al. Bioinformatics tools for development of fast and cost effective simple sequence repeat (SSR),and single nucleotide polymorphisms (SNP) markers from expressed sequence tags(ESTs)[J].African Journal of Biotechnology,2013,12(30):4713-4721.
[50] LIU J,LI J,QU J,et al. Development of genome-wide insertion and deletion polymorphism markers from next-generation sequencing data in rice[J].Rice,2015,8(1):27.
猜你喜欢
——和田盘羊
