广西宁明组植物化石中黄铁矿及其沉积意义
2018-02-27马福军吴媛欣刘松孙柏年闫德飞张凤太王秋军
马福军,吴媛欣,刘松,孙柏年,闫德飞,张凤太,王秋军
1.贵州师范学院地理与旅游学院&贵州省地理国情监测重点实验室,贵阳 550018 2.黔东南苗族侗族自治州地质公园管理局,贵州凯里 556000 3.兰州大学地质科学与矿产资源学院,兰州 730000 4.贵州大学资源与环境工程学院&贵州省古生物研究中心,贵阳 550025
0 引言
化石降解后残留的有机生物分子(如植物角质层),或者埋藏时被充填的矿物(如CaCO3、SiO2、FeS2等),保存有生物体原有的组织结构信息,能为认识过去生命体提供最直接的证据[1- 3]。化石的一些降解过程,伴随有一些矿化现象。理解生物降解和矿化过程,有助于认识生物的化石化过程[2,4]。黄铁矿化是生物体化石化的主要途径之一,已被证实可形成特殊保存的动物软体组织和植物内部结构,以植物化石中较常见[2,5- 7]。化石内赋存的黄铁矿,对认识生物化石化过程中的沉积微环境有着重要意义。尽管对植物化石中的黄铁矿研究已取得了一定的进展,但对其形成的机制和指示的沉积微环境分析较少,一些学者根据研究成果提出了不同的推测[1- 2,5,7- 9]。对于植物化石如何黄铁矿化,黄铁矿化是否与硫化细菌、生物聚合物、生物自身的有机分子等相关存在不同的观点[2,10- 13]。探寻植物化石化过程中,黄铁矿形成的沉积微环境,是一个尚未引起重视、但对解释植物化石形成和保存又是十分关键的研究领域[6]。
黄铁矿具多种保存形式,沉积中最常见的是莓状体和单晶[14- 15]。莓状体是由大量(102~106个)等径、同形态的黄铁矿单晶聚集而成的球体或亚球体,直径为1~250 μm;常见为1.0~20.0 μm,直径大于50 μm的莓状体较少见[9,15- 16]。单晶直径为0.2~2 μm,单个莓状体内的单晶形态、大小较一致[15]。沉积地层中莓状体最初的成核、生长和成分,与沉积环境的氧化还原状态密切相关,其形成后的形状、大小和结构都较稳定,甚至不随沉积环境变迁和矿物相变化而变化,其最初的形态特征能保存[6,16- 17]。在贫氧的水体环境(the dysoxic water conditions),黄铁矿能形成直径较大的单晶、莓状体和莓状体聚合体,且莓状体的直径变化幅度相对较大;而在静水—缺氧水体环境(the anoxic- euxinic water conditions),黄铁矿形成大小相对一致,直径变化幅度相对窄的单晶和莓状体[18- 19]。因此,莓状体的大小(以及结构和成份)可用来分析其形成时氧化还原环境的特征,并已获得沉积学研究的证实[7,18,20],如平均直径小于20 μm莓状体,是形成于周围水体氧浓度小于30 μmol/L的贫氧环境[6,16]。
广西宁明组产大量的、保存较好的植物化石[21],化石内部保存有较多的黄铁矿。尽管近年来对该组的沉积环境、孢粉、植物化石等进行了研究[22- 34],但对该地层植物化石的化石化沉积微环境以及保存大量的黄铁矿缺少探讨。本文基于对宁明组植物化石中的黄铁矿研究,进行粒径分布和成份分析,探讨该地层中植物化石化过程中的沉积微环境,为今后解释植物化石形成和保存提供新的信息。
1 材料与方法
本文研究的材料为采自广西崇左市宁明县城中镇西北方向宁明组的一个剖面(图1a)。宁明组主要是浅湖相沉积,其岩性以灰黄色至棕灰色的薄层状泥岩为主,夹浅黄色泥质砂岩、泥质粉砂岩、细砂岩以及多层膨润土[35- 36]。在灰色、浅灰白色和棕灰色的薄层状泥岩以及泥质砂岩中保存有许多植物叶片和果实等压型化石以及孢粉化石。根据岩石地层、孢粉组合以及鱼类化石等研究结果[22- 23,36- 39],宁明组的地质时代被认为是渐新世。
植物化石标本采于宁明组的薄层状泥岩以及泥质砂岩中(图 1b)。植物化石的黄铁矿样品获得过程如下:将植物化石从岩石上剥下,用5%的H2O2试剂均匀的涂抹在标本上,期间用毛刷去掉覆盖的岩石,然后用蒸馏水清洗干净;置于10%HCl溶液中,将吸附和充填在植物化石上的钙质成分完全除去,直至浸泡试剂由黄色变成白色,然后用蒸馏水清洗干净;再经50%的HF溶液浸泡3天,将吸附在植物化石上的硅质成分充分溶解,清除难溶物质,并用蒸馏水多次清洗干净。一部分样品经5%的舒氏液处理后,依照角质层制作方法制成玻片[40- 42],在兰州大学用LeicaDM4000B多功能生物显微镜进行观察;另一部分样品直接喷金后在中科院兰州化学物理研究所和贵州大学分析测试中心用扫描电镜观察和多功能电子能谱仪元素分析。所有标本及处理得到的角质层玻片和一部分电镜座片保存于兰州大学古生物学与地层学研究所标本室,另一部分电镜座片保存于贵州师范学院地理与旅游学院。
2 结果
2.1 黄铁矿形态和大小
光学玻片和电镜样品都可见黄铁矿保存,可见离散型单晶和成群的莓状体(图2a~f)。植物化石内部保存有大量的黄铁矿,为原位保存,后期的充填作用不明显(图2a,b,c),局部可见黄铁矿掉落后的凹坑。离散型单晶和成群的莓状体共同保存,大小成正态分布(表1),莓状体占主体,成群保存,间隙中夹有单晶。莓状体为圆球状,粒径为7.23~14.89 μm,平均约11.18 μm(图2a,b,c,f);每个莓状体由大量等径的单晶密集而成,单晶直径为0.87~1.54 μm,平均约1.16 μm(图2a,b,c),形状为无规则至球状体,晶棱不明显,极少量单晶的表面可见凹点。莓状体周围有粒径较小的无序堆积的单晶,形状与莓状体的单晶较相似,直径为0.83~1.03 μm,平均约0.94 μm(图2a~d)。

图1 样品采集地(a)和宁明组地层柱状图(b.据宁宗善等,1994[36])Fig.1 Map showing the sample locality (a) and the generalized stratigraphic section of the Ningming Formation (b. after Ning et al., 1994)[36]
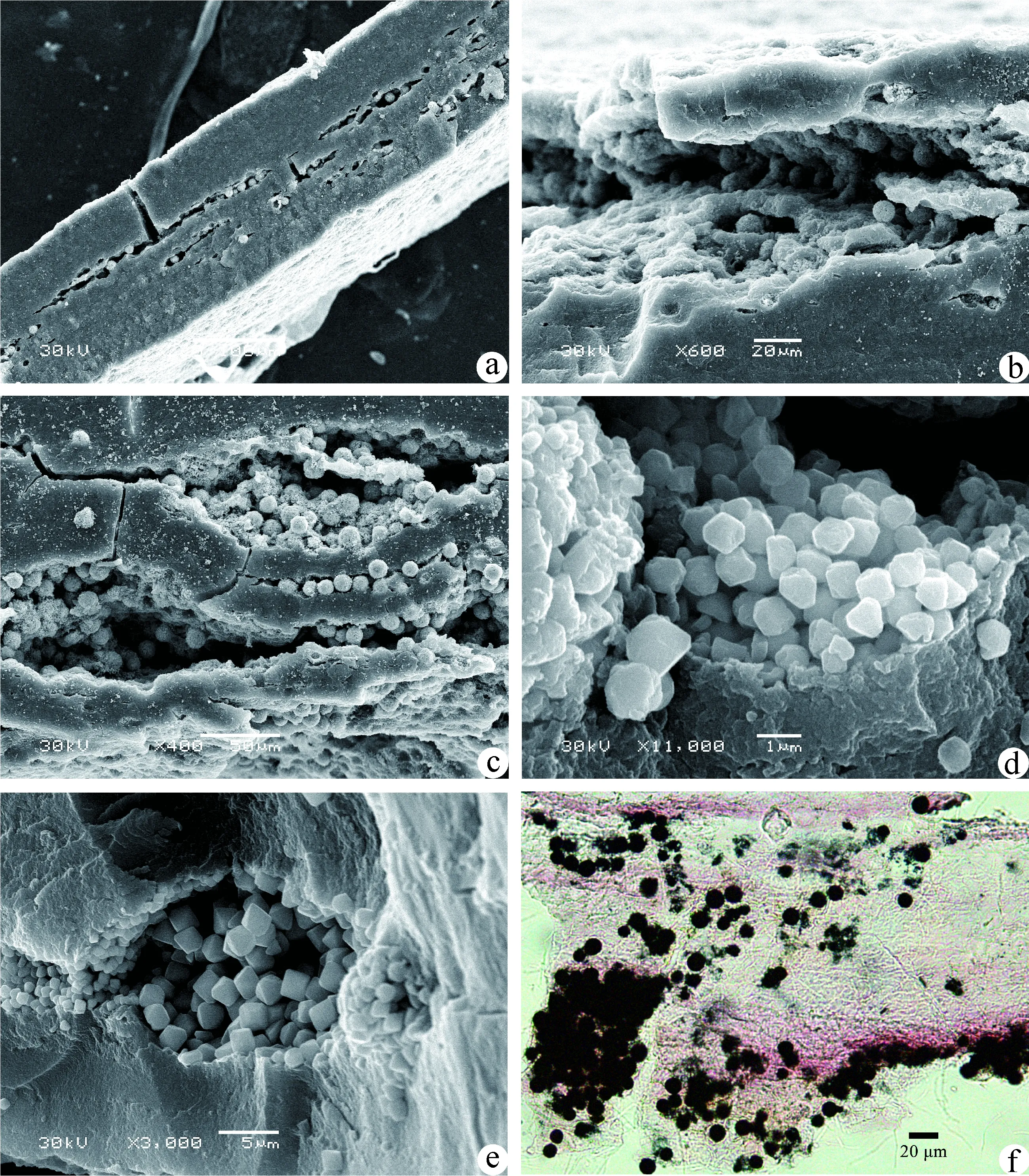
图2 植物化石中的黄铁矿a,b,c.单晶和莓状体层状充填;d.c图中局部放大,显示莓状体周围的离散型单晶;e.八面体的晶体;f.角质层上的黄铁矿Fig.2 Pyrites preserved in plant fossilsa, b, c. dissolved crystals and framboidal clusters preserved in layers between the abaxial and adaxial cuticles of plant fossils; d. enlargement of a portion of ‘c’ showing dissolved crystals scattered around the framboidals; e. crystals in the octahedron shape; f. pyrites adhered to the plant cuticle

成群的莓状体离散型单晶大小/μm≤10.010.0~13.0≥13.0≤0.080.08~1.20≥1.20个数21741211298比例/%19.669.211.222.960.416.7
少数黄铁矿的单晶全为八面体,表面光滑,晶棱清晰(图2e)。莓状体粒径为11.56~14.95 μm,平均约12.68 μm;莓状体单晶的直径为1.88~2.36 μm,平均约2.26 μm。莓状体周围有粒径较小、形态相似、无序堆积的离散型单晶,直径为0.79~1.58 μm,平均约1.14 μm。
2.2 莓状体能谱测量
利用能谱仪对莓状体中的单晶进行了测试。图3和表2显示,单晶的主要成份为S和Fe,其质量百分比的和不少于71.98 %;硫、铁的原子个数比值为0.16~2.06,均值为1.54;另含有少量其他元素。
2.3 莓状体的粒径均值和标准差
对同一层位中9个样品的莓状体粒径均值(Mean framboid size,简写M)和标准差(Standard deviation,简写SD)进行了统计(表3),发现不同样品中M和SD存在差别。M为10.6~12.2 μm,SD为0.2~2.2 μm。
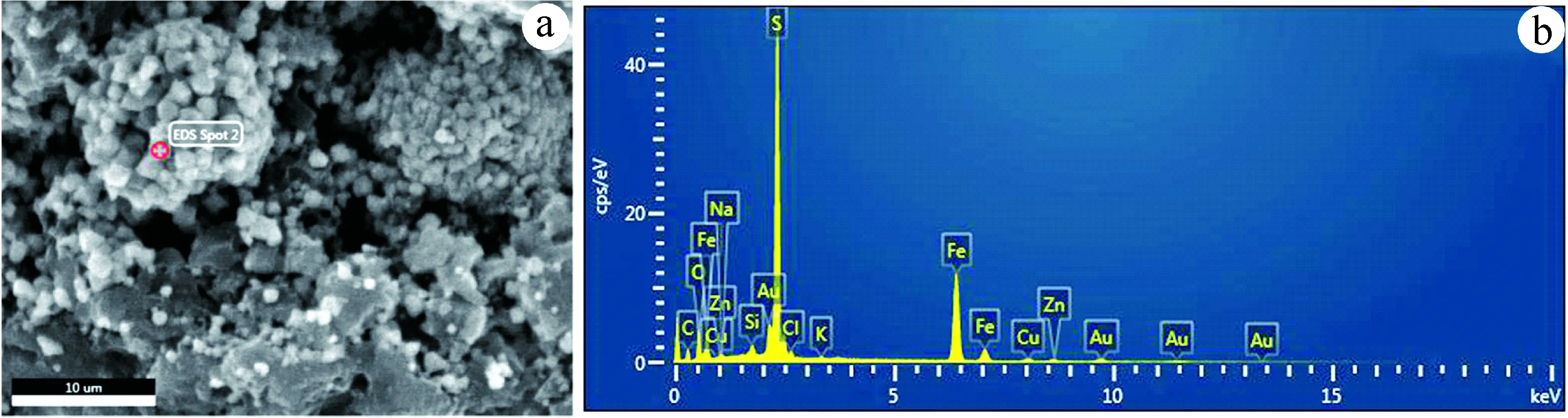
图3 莓状体单晶的能谱示例图a.能谱分析;b.元素含量Fig.3 A set of sample graphs showing the energy spectra (EDS) of a single crystal in a pyrite framboidala. EDS analysis; b. element contents

样点SFe质量比/%原子比/%质量比/%原子比/%S/Fe原子比样点136.2723.3535.7113.241.76样点253.7866.9646.2233.042.03样点346.5043.8039.6821.452.04样点452.5165.8247.4934.181.93样点552.4365.7547.5734.251.92样点639.0029.8233.9214.892.00样点748.8665.0743.5433.291.95样点848.2564.6643.6233.571.93样点950.0665.9143.1932.652.02样点1052.6465.9447.3634.061.94样点1149.2965.4743.1232.891.99样点1247.5664.3543.333.631.91样点1347.6665.4940.3531.832.06样点1431.2544.1868.7555.820.79样点1545.7464.0540.9332.911.95样点1619.7129.9580.2970.050.43样点178.6214.1191.3885.890.16样点1811.2318.0588.7781.950.22样点1910.216.5289.883.480.20平均值1.54

表3 不同样品的莓状体粒径均值和标准差
3 讨论
黄铁矿单晶的大小和形态与结晶时基质浓度密切相关,水体中单硫相溶质的超饱和是控制单晶形成和生长的主要因素[13,18]。由于立方体具有较小的表面自由能,最初的单晶都是小立方体,随溶液中单硫相溶质超饱和的增强,立方体单晶能形成为五角十二面体、八面体和球状体的单晶[18]。通过实验模拟发现[17],在快速的反应时间内,离散型单晶和成群莓状体的粒径和直径变化幅度相对较小,其中莓状体能指示其形成时水体的氧化还原环境。
沉积地层中,莓状体形成于成岩过程的早期阶段,通常位于沉积层顶面下几厘米处[18,20,43]。在不同的底部水体环境中,能形成不同种类的莓状体[18- 20,44- 45](图4),为成岩黄铁矿(diagenetic pyrites)和同生黄铁矿(syngenetic pyrites):1)在贫氧的水体环境(the dysoxic water conditions),氧化还原层(the redox interface)位于沉积层顶面下,靠近沉积层—水体的接触面,缺氧孔隙水中形成成岩黄铁矿微晶和微晶聚合体,后期埋藏过程中能形成莓状体、莓状聚合体、微晶聚合体,其中莓状体的粒径为4~50 μm,粒径变化幅度相对较大(图4a);2)随氧浓度的降低,水体转变为一种静水—缺氧水体环境(the anoxic- euxinic water conditions),氧化还原层上升至沉积层上的水体中,形成同生黄铁矿微晶和微晶聚合体,短时间内的下降沉淀过程中形成莓状体和莓状聚合体,莓状体的平均粒径较小(<6 μm),粒径变化幅度相对窄(图4b)。实验模拟研究表明[45- 46]:在缺氧水体环境中,莓状体的粒径最大值小于 20 μm。而在富氧的水体环境(the oxic water conditions),黄铁矿只形成少量的单晶,基本不存在莓状体[19]。因此,莓状体粒径大小和变化幅度可应用于生物化石化时沉积微环境分析[6,16,18,47]。
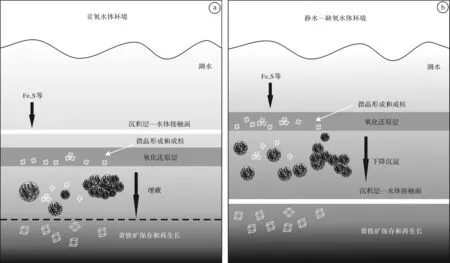
图4 不同水体环境的黄铁矿形成模式(据Wang et al. [18]修改)a.贫氧水体环境;b.静水—缺氧水体环境Fig.4 Relationships between different pyrite forms and water conditions (modified from Wang et al. [18])a. oxic- dysoxic water condition; b. euxinic- anoxic water condition
3.1 宁明组植物化石中黄铁矿沉积环境推测
邝国敦等[23]基于岩性和化石的保存情况,推测宁明组含植物化石的层段为一种湖泊相浅水砂泥坪环境。对于宁明组沉积期间,湖水整体的氧化还原相演化情况一直还未有相关研究;本文拟基于宁明组植物化石中的黄铁矿,探讨该地层中植物化石化过程中的内部沉积微环境。光学显微镜和扫描电镜显示,植物化石内黄铁矿分为单晶和莓状体,其中单晶为无规则至球状体(图2a~d)和八面体(图2e),指示了单晶不同的生长时期。由于单晶直径为0.79~1.58 μm,变化幅度小于0.08 μm;莓状体粒径为7.23~14.95 μm(其均值大于6 μm),变化幅度小于8 μm;且直径和粒径大小均处于低值段(表1);因此单晶直径和莓状体粒径的大小和变化幅度均说明黄铁矿是在短时间内形成的,且植物体上、下表皮间为一种贫氧的水体环境(图4a)。当前莓状体粒径最大值为14.95 μm,小于 20 μm,同样支持了以上对植物体上、下表皮间水体氧化还原环境的推测。
另外,黄铁矿的S/Fe原子个数比值是黄铁矿铁品位一个重要参数,正常溶解氧条件下S/Fe值为2,浓度略高的溶解氧条件下S/Fe值越高,浓度略低的溶解氧条件下S/Fe值越低[16,18]。宁明组植物化石中莓状体的S/Fe值为0.16~2.06,均值为1.54(小于2),指示黄铁矿形成时,植物上、下表皮组织内溶解氧供应不足,为一种贫氧的水体微环境。
而植物化石上、下表皮间组织结构未保存,而充填有大量的、近似层状分布的黄铁矿(图2a,b,c),表明黄铁矿形成时,上、下表皮间组织结构已发生了较强烈的降解。当前结果支持了化石内莓状体形成与有机质降解之间存在着必然联系的结论[6]。而保存的莓状体形态一致,为圆球状(图2a,b,c,f),说明黄铁矿未发生挤压变形,在地层挤压前已形成。
3.2 莓状体粒径均值—标准差的分布分析
莓状体粒径的均值(M)和标准差(SD)的分布图能为区分贫氧的水体环境和静水—缺氧环境提供信息[19,48- 49]。现代贫氧沉积中,M为5.7~11.9 μm,SD为2.5~7.9 μm;而缺氧沉积中,M范围为4.3~6.1 μm,SD小于2.0 μm[45]。基于对现代和古代沉积中莓状体的分析表明[19- 20,48],莓状体粒径的均值和标准差的关系式为M=-3.3SD+14,该关系式同时是定量界定贫氧和静水—缺氧沉积环境的界线(图5)。

图5 莓状体粒径均值(M)—标准差(SD)分布图(数据来自表3)虚线是界定贫氧和静水—缺氧水体环境的界线(M=-3.3SD+14)Fig.5 Cross- plot diagram showing mean framboid diameter (M) versus standard deviation (SD) (data from Table 3)The dash line separating anoxic- euxinic water conditions from dysoxic water conditions (M =-3.3SD+14)
对来自同层位9个样品中的黄铁矿莓状体粒径均值—标准差的分布进行了分析(表3、图5),其中一个样的M- SD值稍位于界线下,即位于静水—缺氧水体环境区;另一个样的M- SD值位于界线附近;其他7个样的M- SD值均位于界线上,即位于贫氧水体环境区。因此,M- SD分布图整体支持当前的推测:黄铁矿形成时,虽然存在局部微环境为静水—缺氧水体环境,但植物体上、下表皮间整体为一种贫氧的水体环境(图5)。
4 结论
(1) 对广西宁明组植物化石中的黄铁矿进行了研究,单晶绝大多数为无规则至球状体,晶棱不明显,表面可见凹点;少数为八面体,晶棱明显,表面光滑;单晶直径为0.79~1.58 μm;莓状体为圆球状,粒径为7.23~14.95 μm;S/Fe原子个数比值为0.16~2.06,均值为1.54。
(2) 莓状体的粒径大小和变化幅度,S/Fe原子个数比值,以及粒径均值—标准差分布图均表明植物在化石化黄铁矿形成时期,其上、下表皮组织内为一种贫氧的水体环境,并在短时间内形成了莓状体。
(3) 植物化石上、下表皮间组织结构未保存,而充填有大量的、近似层状分布的黄铁矿,表明黄铁矿形成时,上、下表皮间组织结构已发生了较强烈的降解。
)
[1] Garcia- Guinea J, Martinez- Frías J, Harffy M. Cell- hosted pyrite framboids in fossil woods[J]. Naturwissenschaften, 1998, 85(2): 78- 81.
[2] Grimes S T, Brock F, Rickard D, et al. Understanding fossilization: experimental pyritization of plants[J]. Geology, 2001, 29(2): 123- 126.
[3] Ma F J, Wang Q J, Dong J L, et al. A new plant assemblage from the middle Triassic volcanic tuffs of Pingchuan, Gansu, northwestern China and its paleoenvironmental significance[J]. Paläontologische Zeitschrift, 2016, 90(2): 349- 376.
[4] Gabbott S E, Hou X G, Norry M J, et al. Preservation of early Cambrian animals of the Chengjiang biota[J]. Geology, 2004, 32(10): 901- 904.
[5] Grimes S T, Davies K L, Butler I B, et al. Fossil plants from the Eocene London Clay: the use of pyrite textures to determine the mechanism of pyritization[J]. Journal of the Geological Society, 2002, 159(5): 493- 501.
[6] 冷琴,杨洪. 中国东北热河生物群化石及沉积物中的黄铁矿莓状体及其对早期化石化阶段沉积微环境的指示[J]. 自然科学进展,2003,13(4):359- 364. [Leng Qin, Yang Hong. Pyrite framboids associated with the Mesozoic Jehol Biota in northeastern China: implications for microenvironment during early fossilization[J]. Progress in Natural Science, 2003, 13(4): 359- 364.]
[7] Brock F, Parkes R J, Briggs D E G. Experimental pyrite formation associated with decay of plant material[J]. Palaios, 2006, 21(5): 499- 506.
[8] Sugawara H, Sakakibara M, Belton D, et al. Formation process of pyrite polyframboid based on the heavy- metal analysis by micro- PIXE[J]. Environmental Earth Sciences, 2013, 69(3): 811- 819.
[9] Wacey D, Kilburn M R, Saunders M, et al. Uncovering framboidal pyrite biogenicity using nano- scale CNorgmapping[J]. Geology, 2015, 43(1): 27- 30.
[10] Schoonen M A A. Mechanisms of sedimentary pyrite formation[J]. Special Papers- Geological Society of America, 2004, 379: 117- 134.
[11] Folk R L. Nannobacteria and the formation of framboidal pyrite: textural evidence[J]. Journal of Earth System Science, 2005, 114(3): 369- 374.
[12] MacLean L C W, Tyliszczak T, Gilbert P U P A, et al. A high- resolution chemical and structural study of framboidal pyrite formed within a low- temperature bacterial biofilm[J]. Geobiology, 2008, 6(5): 471- 480.
[13] Mozer A. Authigenic pyrite framboids in sedimentary facies of the mount wawel Formation (Eocene), king George Island, West Antarctica[J]. Polish Polar Research, 2010, 31(3): 255- 272.
[14] 余江滨,潘侊. 原始黄铁矿莓体的发现与研究[J]. 矿物学报,1988,8(4):357- 364. [Yu Jiangbin, Pan Guang. Discovery of primary pyrite framboids and their study[J]. Acta Mineralogica Sinica, 1988, 8(4): 357- 364.]
[15] Soliman M F, El Goresy A. Framboidal and idiomorphic pyrite in the upper Maastrichtian sedimentary rocks at Gabal Oweina, Nile Valley, Egypt: formation processes, oxidation products and genetic implications to the origin of framboidal pyrite[J]. Geochimica et Cosmochimica Acta, 2012, 90: 195- 220.
[16] 李洪星,陆现彩,边立曾,等. 有孔虫壳体内草莓状黄铁矿成因及其地质意义:以湖北雁门口地区栖霞组有孔虫化石为例[J]. 高校地质学报,2009,15(4):470- 476. [Li Hongxing, Lu Xiancai, Bian Lizeng, et al. Formation of pyrite framboids in the chamber of foraminiferas and its geological significance: A case study of the foraminiferas fossils in the Qixia Formation in the Yanmenkou area, Hubei province[J]. Geological Journal of China Universities, 2009, 15(4): 470- 476.]
[17] Butler I B, Rickard D. Framboidal pyrite formation via the oxidation of iron (II) monosulfide by hydrogen sulphide[J]. Geochimica et Cosmochimica Acta, 2000, 64(15): 2665- 2672.
[18] Wang P K, Huang Y J, Wang C S, et al. Pyrite morphology in the first member of the Late Cretaceous Qingshankou Formation, Songliao Basin, northeast China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 385: 125- 136.
[19] Huang Y G, Chen Z Q, Wignall P B, et al. Latest Permian to Middle Triassic redox condition variations in ramp settings, South China: pyrite framboid evidence[J]. Geological Society of America Bulletin, 2016, 129(1/2): 229- 243.
[20] Wignall P B, Newton R. Pyrite framboid diameter as a measure of oxygen deficiency in ancient mudrocks[J]. American Journal of Science, 1998, 298(7): 537- 552.
[21] 史恭乐. 广西渐新世宁明组植物化石及植物群古气候初步重建[D]. 南京:中国科学院研究生院,2010. [Shi Gongle. Fossil plants from the Oligocene Ningming Formation of Guangxi, and a preliminary palaeoclimatic reconstruction of theora[D]. Nanjing: Nanjing Institute of Palaeontology and Geology, Chinese Academy of Sciences, 2010.]
[22] 王伟铭,陈耿娇,陈运发,等. 广西宁明盆地第三纪孢粉植物群及其地层意义[J]. 地层学杂志,2003,27(4):324- 327. [Wang Weiming, Chen Gengjiao, Chen Yunfa, et al. Tertiary palynostratigraphy of the Ningming Basin, Guangxi[J]. Journal of Stratigraphy, 2003, 27(4): 324- 327.]
[23] 邝国敦,陈耿娇,陈运发,等. 广西宁明盆地第三纪地层研究的新进展[J]. 地层学杂志,2004,28(4):362- 367. [Kuang Guodun, Chen Gengjiao, Chen Yunfa, et al. New information on the Tertiary biostratigraphy of the Ningming basin, Guangxi[J]. Journal of Stratigraphy, 2004, 29(4): 362- 367.]
[24] 李浩敏,陈运发,陈耿娇,等. 广西宁明第三纪类黄杞翅果化石[J]. 古生物学报,2003,42(4):537- 547. [Li Haomin, Chen Yunfa, Chen Gengjiao, et al. Tertiary fossil winged fruits ofPalaeocaryafrom Ningming of Guangxi, S. China[J]. Acta Palaeontologica Sinica, 2003, 42(4): 537- 547.]
[25] Shi G L, Xie Z M, Li H M. High diversity of Lauraceae from the Oligocene of Ningming, South China[J]. Palaeoworld, 2014, 23(3/4): 336- 356.
[26] 宋柱秋,史恭乐,陈运发,等. 广西渐新统宁明组臭椿属(苦木科)的翅果化石及其分类学和生物地理学意义[J]. 古生物学报,2014,53(2):191- 200. [Song Zhuqiu, Shi Gongle, Chen Yunfa, et al. Winged fruits ofAilanthus(Simaroubaceae) from the Oligocene Ningming Formation of Guangxi, and their taxonomic and biogeographic implications[J]. Acta Palaeontologica Sinica, 2014, 53(2): 191- 200.]
[27] Wang Q, Song Z Q, Chen Y F, et al. Leaves and fruits ofBauhinia(Leguminosae, Caesalpinioideae, Cercideae) from the oligocene Ningming Formation of Guangxi, South China and their biogeographic implications[J]. BMC Evolutionary Biology, 2014, 14: 88.
[28] Chen Y F, Manchester S R. Winged fruits ofDeviacerin the Oligocene from the Ningming basin in Guangxi, South China[J]. PLoS One, 2015, 10(12): e0144009.
[29] Dong J L, Sun B N, Ma F J, et al.Paliurusfruits from the Oligocene of South China and their phytogeographic implications[J]. PLoS One, 2015, 10(11): e0140653.
[30] Dong J L, Wang W J, Deng P, et al. New material ofPalaeocaryafrom the Oligocene of Ningming, Guangxi, South China[J]. Paläontologische Zeitschrift, 2016, 90(2): 399- 412.
[31] Ma F J, Sun B N, Wang Q J, et al. A new species ofMeliolinitesassociated withBuxusleaves from the Oligocene of Guangxi, southern China[J]. Mycologia, 2015, 107(3): 505- 511.
[32] Ma F J, Wang Q J, Dong J L, et al.Buxusleaves from the Oligocene of Guangxi, China and their biogeographical significance[J]. Acta Geologica Sinica: English Edition, 2015, 89(5): 1453- 1469.
[33] Wang Q J, Ma F J, Dong J L, et al. Coryphoid palms from the Oligocene of China and their biogeographical implications[J]. Comptes Rendus Palevol, 2015, 14(4): 263- 279.
[34] Sun B N, Wang Q J, Konrad W, et al. Reconstruction of atmospheric CO2during the Oligocene based on leaf fossils from the Ningming Formation in Guangxi, China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 467: 5- 15.
[35] 广西壮族自治区地质矿产局. 广西壮族自治区区域地质志[M]. 北京:地质出版社,1985:273- 274. [Bureau of Geology and Mineral Resources of Guangxi Zhuang Autonomous Region. Regional geology of Guangxi Zhuang Autonomous Region[M]. Beijing: Geological Publishing House, 1985: 273- 274.]
[36] 宁宗善,胡炎坤,周铁明,等. 中国油气区第三系(Ⅶ)滇桂油气区分册[M]. 北京:石油工业出版社,1994:13- 20. [Ning Zongshan, Hu Yankun, Zhou Tieming, et al. Tertiary in petroliferous regions of China (Ⅶ)[M]. Beijing: Petroleum Industry Press, 1994: 13- 20.]
[37] 陈耿娇,陈运发,邝国敦,等. 广西宁明盆地第三纪鱼类化石生物地层[J]. 古脊椎动物学报,2004,42(1):81- 85. [Chen Gengjiao, Chen Yunfa, Kuang Guodun, et al. Biostratigraphy of Tertiary fossil fishes from Ningming Basin, Guangxi[J]. Vertebrata Palasiatica, 2004, 42(1): 81- 85.]
[38] Chen G J, Fang F, Chang M M. A new cyprinid closely related to cultrins+xenocyprinins from the mid- Tertiary of South China[J]. Journal of Vertebrate Paleontology, 2005, 25(3): 492- 501.
[39] Chen G J, Chang M M. A new early cyprinin from Oligocene of south China[J]. Science China Earth Sciences, 2011, 54(4): 481- 492.
[40] 马福军,徐小慧,李瑞云,等. 内蒙古乌尼特煤田下白垩统分散角质层研究[J]. 微体古生物学报,2013,30(3):244- 262. [Ma Fujun, Xu Xiaohui, Li Ruiyun, et al. Dispersed cuticles from the lower Cretaceous Wunite coal field in Inner Mongolia[J]. Acta Micropalaeontologica Sinica, 2013, 30(3): 244- 262.]
[41] 王秋军,徐小慧,金培红,等. 基于银杏类化石Baierafurcata气孔参数定量重建中生代古大气CO2浓度变化[J]. 地质论评,2013,59(6):1035- 1045. [Wang Qiujun, Xu Xiaohui, Jin Peihong, et al. Quantitative reconstruction of Mesozoic paleoatmospheric CO2based on stomatal parameters of fossilBaierafurcataof Ginkgophytes[J]. Geological Review, 2013, 59(6): 1035- 1045.]
[42] Wang Q J, Ma F J, Yang Y, et al. Bamboo leaf and pollen fossils from the late Miocene of eastern Zhejiang, China and their phytogeological significance[J]. Acta Geologica Sinica (English Edition), 2014, 88(4): 1066- 1083.
[43] Wilkin R T, Barnes H L. Formation processes of framboidal pyrite[J]. Geochimica et Cosmochimica Acta, 1997, 61(2): 323- 339.
[44] Sawlowicz Z. Pyrite framboids and their development: a new conceptual mechanism[J]. Geologische Rundschau, 1993, 82(1): 148- 156.
[45] Wilkin R T, Barnes H L, Brantley S L. The size distribution of framboidal pyrite in modern sediments: an indicator of redox conditions[J]. Geochimica et Cosmochimica Acta, 1996, 60(20): 3897- 3912.
[46] Wilkin R T, Barnes H L. Pyrite formation by reactions of iron monosulfides with dissolved inorganic and organic sulfur species[J]. Geochimica et Cosmochimica Acta, 1996, 60(21): 4167- 4179.
[47] Hethke M, Fürsich F T, Jiang B Y, et al. Oxygen deficiency in Lake Sihetun; formation of the lower Cretaceous Liaoning Fossillagerstätte (China)[J]. Journal of the Geological Society, 2013, 170(5): 817- 831.
[48] Bond D P G, Wignall P B. Pyrite framboid study of marine Permian- Triassic boundary sections: A complex anoxic event and its relationship to contemporaneous mass extinction[J]. Geological Society of America Bulletin, 2010, 122(7/8): 1265- 1279.
[49] Tian L, Tong J N, Algeo T J, et al. Reconstruction of Early Triassic ocean redox conditions based on framboidal pyrite from the Nanpanjiang Basin, South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 412: 68- 79.
