不同小麦品种颖果背部维管束发育研究
2018-02-13刘大同别同德高德荣
刘大同, 陈 明, 别同德, 张 勇, 高德荣,
(1.江苏里下河地区农业科学研究所/农业部长江中下游小麦生物学与遗传育种重点实验室,江苏扬州 225007;2.扬州大学/江苏省粮食作物现代产业技术协同创新中心,江苏扬州 225009)
小麦(TriticumaestivumL.)颖果中的背部维管束是母体养分运输组织,是光合同化物和水进入颖果的主要通道,与籽粒的灌浆充实速率和品质等密切相关[1-2]。花后叶片制造的光合产物以及茎秆中积累的养分经维管束进入颖果,在颖果背部维管束的筛管发生同化物卸载,然后通过合点和珠心突起传递细胞进入胚乳腔中,再经质外体或共质体途径供应胚和胚乳的发育[1,3-4]。
一般情况下,韧皮部运输光合产物等有机物质,木质部运输水分和无机盐。由于人们普遍关注小麦维管束发育与籽粒灌浆中同化物的运输,因此对韧皮部的结构发育研究较多[5-7];而对颖果中的水分没有足够重视,因此对木质部研究较少。但研究发现,小麦颖果与小花轴维管束连接处的木质部被一组特殊的厚壁细胞“切断”,使小花轴内的维管束导管与颖果内的维管束导管失去连贯性,但韧皮部连贯性不受影响[8-9],因此在灌浆过程中,水分可能是以光合同化物溶剂的形式进入颖果的。Crafts等认为,当运入库器官中的水分超过需求时,多余的水有可能通过木质部流回母体[10];王忠等在研究水稻和小麦维管束向胚乳运送物质时提出,木质部的导管可能是作为排水管来排除灌浆液流中的多余水分[1,11-12],但均未被研究证实。因此,小麦颖果发育过程中维管束的发育特征还有许多尚未阐明的重要问题,值得进一步深入探究。
扬麦16是目前长江中下游麦区种植面积最大的小麦品种,具有灌浆快、脱水快的特点[3,13-14]。研究表明,扬麦16灌浆速率、灌浆完成期(籽粒干质量达最大值的日期)籽粒含水率及此后的籽粒脱水速率均显著高于对照品种扬麦15、宁麦13等品种[3,13]。目前关于扬麦16灌浆快的机理,从颖果显微结构和内源生长素含量等方面进行了解释[3],但关于脱水快的机理尚未见研究报道。本研究以扬麦16和扬麦15为材料,利用树脂切片方法,对花后不同时间的颖果背部维管束的发育进行研究,分析了韧皮部和木质部的形态变化规律,旨在为揭示扬麦16及同类型品种灌浆快、脱水快的机理提供形态学研究基础。
1 材料与方法
1.1 材料
试验于2014—2017年在江苏里下河地区农业科学研究所湾头试验基地(地理位置为119°42′E、32°39′N)进行,该地为亚热带季风气候,年平均气温14.8~15.3 ℃,年平均降水量961~1 048 mm。以长江中下游麦区主栽品种扬麦16和扬麦15为材料,生长期常规化管理,及时防控病虫害。为获取不同发育时间的颖果,在开花期采用记号笔点颖与植株挂牌相结合的方法正确标记开花日期,标记的颖花均为穗中部小穗基部的2朵花。从花后5 d开始取样,直到种子成熟,选取花后5、10、15、20、25 d的颖果进行研究。
1.2 方法
1.2.1 颖果的光学显微镜观察 利用树脂半薄切片法观察颖果的显微结构。将小麦颖果从中部横切,取2 mm厚的切段,置于2.5%戊二醛(25%戊二醛溶于pH值为7.2的磷酸缓冲液中稀释)中固定(4 ℃)4 h。固定好的样品经pH值为7.2的磷酸缓冲液清洗(3次,每次10 min)、梯度(20%、40%、60%、70%、80%、90%、95%、100%)乙醇脱水(3次,每次 10 min),环氧丙烷置换,Spurr低黏度树脂浸透与包埋(70 ℃ 聚合10 h)。聚合好的样品块先用玻璃刀在超薄切片机(Ultracut R,Leica,Germany)上切1 μm厚半薄切片,经1%番红和甲基紫重复染色10 min后,在光学显微镜(DMLS,Leica,Germany)下观察并拍照。
1.2.2 颖果横断面的扫描电镜观察 采用固定后的样品经丙酮脱水,乙酸异戊酯过渡,CO2临界点干燥,离子溅射镀金制样后在扫描电子显微镜(Philips XL-30)下观察样品表面和断面结构。
1.2.3 分析作图 图版的制作采用Photoshop图像处理软件(Version CS6,Adobe,USA)。
2 结果与分析
2.1 颖果发育过程中背部维管束区域的解剖结构分析
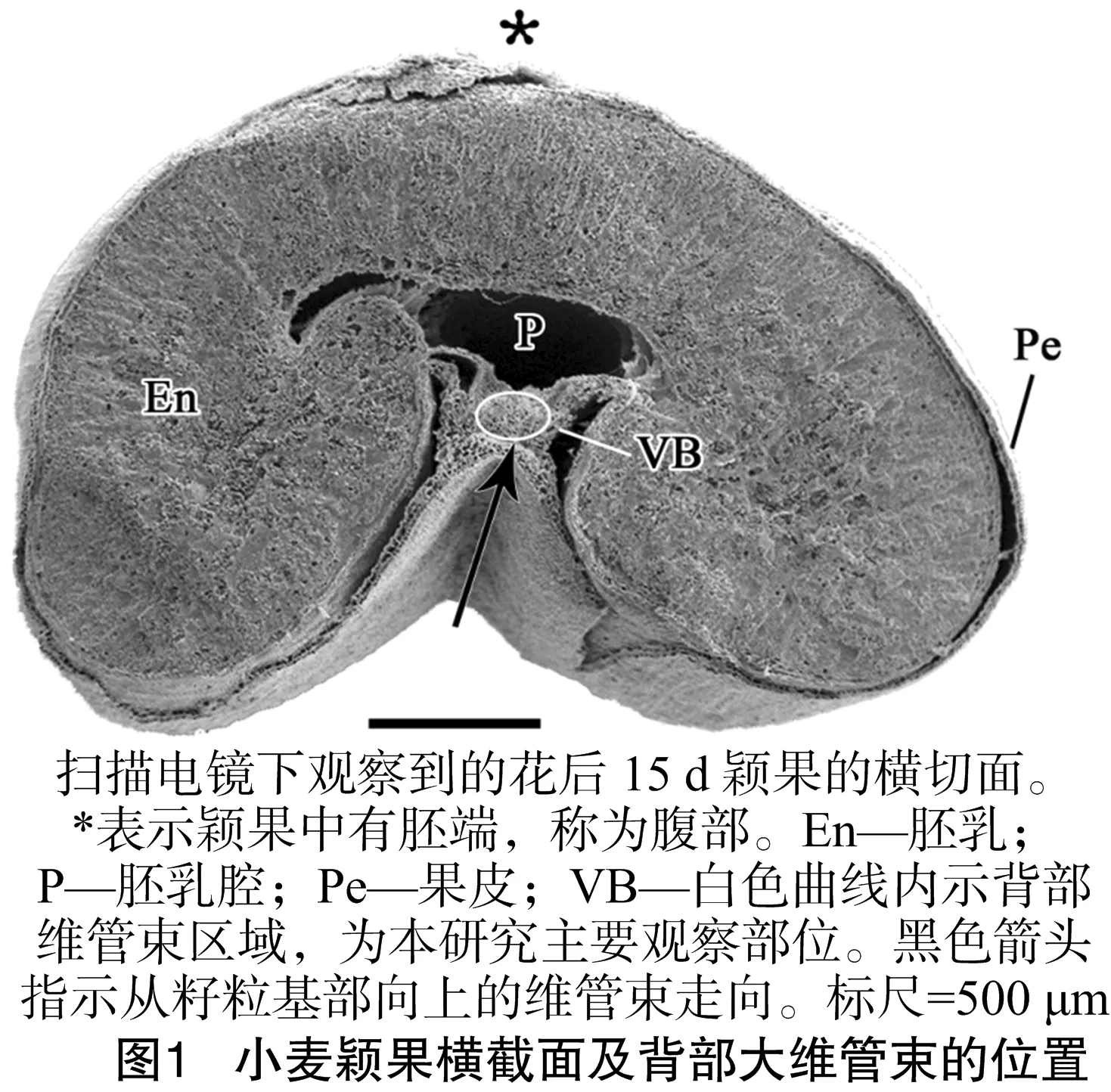
小麦颖果的果皮中有3条维管束,其中颖果两侧通向花柱的2条小维管束在开花后退化,因而不参与灌浆;向颖果输送养分的是背部折痕区的一条大维管束,即背部维管束[15]。在颖果发育的同时,背部维管束发育,由连接小花轴的颖果基部向上贯通到颖果顶端(图1中箭头指示方向),并在侧向上向周围空间卸载有机物,成为灌浆物质的运输通道。由小麦颖果横切面扫描电镜照片可见(图1),背部维管束位于折痕区果皮中央部位,与胚乳之间有质外体空腔——胚乳腔相隔,与胚乳细胞间内没有共质体联系, 因此养分须经过质外体途径才能到达胚乳细胞。
图2是颖果背部折痕区果皮的光镜照片,集中展示了果皮、维管束、合点、珠心突起的发育状况。由图2可见,背部维管束由木质部和韧皮部组成。韧皮部位于外侧,对木质部呈半包围状,且分化早于木质部。花后5~25 d显微图片统计发现,扬麦16和扬麦15的导管数均为6~10个,韧皮部筛管和伴胞数为30~50个,差异不明显。韧皮部距离折痕区果皮的外表皮有5层细胞的厚度。
花后5 d时,颖果背部维管束木质部尚未分化完成,没有观察到明显的导管;韧皮部筛管细胞与周围薄壁细胞已能够清楚区分,但此时筛分子数量较少,周围有大量薄壁细胞,原生质浓厚,细胞核大而明显。此时,果皮细胞中有丰富的淀粉体,体积很小,沿细胞壁分布(图2-A、图2-B)。
花后10 d,筛管数量增加,且聚集在一起呈簇状,在颖果横切面中呈左右对称结构,容易辨认。韧皮薄壁细胞上方为木质部,可见多个细胞壁明显增厚、染色深的导管和许多薄壁细胞。此时,2个小麦品种已经发育出较为完善的输导组织。珠心突起特化出传递细胞,细胞壁增厚,细胞间隙增大,适应养分运输需求(图2-C、图2-D)。
花后15 d,扬麦16果皮细胞中淀粉体的体积持续增大,扬麦15中变化不明显;扬麦16韧皮部维管薄壁细胞变形扭曲,细胞壁发生内陷或凸出,而扬麦15的同类细胞未发生明显改变。扬麦16薄壁细胞中的淀粉体数量和体积均大于扬麦15。合点连接内外珠被,是养分进入胚囊的必经之路。扬麦16合点区细胞内有多个小液泡,由于积累丹宁酸等不透水物质而边缘呈黑色,对养分运输有阻碍作用;扬麦15合点细胞中多为1个大液泡,且液泡边缘颜色较浅。由于扬麦16薄壁细胞的变形和压缩,使得韧皮部到合点的距离减小,可能更有利于同化物快速通过合点到达胚乳腔(图2-E、图2-F)。
花后20 d,扬麦16的维管薄壁细胞进一步变形、压缩,甚至发生破损解体,细胞间隙增加;韧皮部更加靠近合点区,且呈线形排布,与周围细胞接触面也更大,同化物进出的通道更加丰富;合点细胞中液泡积累更多丹宁类物质而黑色加深,小液泡融合形成大液泡;珠心突起传递细胞壁也进一步加厚(图2-G)。扬麦15韧皮部外侧的细胞发生形变,但从韧皮部通往合点之间的薄壁细胞形态没有明显改变,筛管距离合点较远(图2-H)。
花后25 d,扬麦16合点细胞中液泡积累更多丹宁类物质而黑色加深、面积增大,形成了1道未完全封闭的屏障,共质体运输途径受到阻碍;扬麦15合点区细胞的液泡仅边缘为黑色,未形成明显屏障。2个品种的维管束周围薄壁细胞均发生了变形扭曲,珠心突起传递细胞壁均显著增厚,更为突出的是,韧皮部细胞被部分填充,染色加深(图2-I、图2-J)。
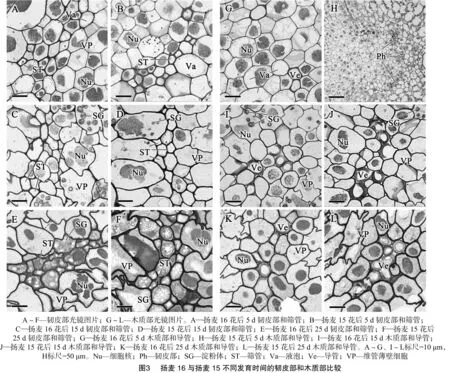
2.2 背部维管束韧皮部和木质部发育的比较
由图3可见,花后5 d,韧皮部和木质部的维管薄壁细胞结构清晰完整,原生质浓厚,细胞核较大且完整,液泡明显。与韧皮部薄壁细胞相比,筛管-伴胞和中间细胞较小,内含物少,细胞壁较厚。此时,扬麦16的木质部已可见类似导管形态的细胞,而扬麦15木质部分化不明显(图3-A、图3-B、图 3-G、图3-H)。
花后15 d,扬麦16韧皮部薄壁细胞扭曲变形,细胞壁内陷或向旁边的细胞外突,筛管形状也有一定改变;木质部中与导管不相邻的薄壁细胞也发生了扭曲,但程度较轻。细胞外形的不规则可能使细胞膜表面积增大,有利于养分跨膜运输。与之相比,扬麦15的薄壁细胞排列较规则,未见明显的细胞形状改变。2个品种靠近果皮的薄壁细胞中均有淀粉体存在,且为复粒淀粉,但扬麦15淀粉体数量少,体积也较小(图3-C、图3-D、图3-I、图3-J)。
花后25 d,2个品种维管薄壁细胞均发生扭曲变形,且扬麦16中程度更重;韧皮部薄壁细胞的细胞核开始发生核裂解,说明细胞结构开始解体,功能衰退。此时,扬麦16韧皮部的筛管及中间细胞大多被压缩成扁条形,在筛管和附近细胞内出现圆球形淀粉粒;而扬麦15中细胞形状与花后15 d相比未发生明显变化,细胞内径较大,淀粉粒不仅在筛分子和附近pan>细胞中积累,在距离较远的薄壁细胞中也出现大量的圆球形淀粉粒。这些现象表明韧皮部输导能力的减弱(图3-E、图3-F)。2个品种的木质部薄壁细胞中也积累了大量淀粉粒。扬麦16导管结构完整,内径较大,且呈线形排列;扬麦15导管内径较小,且有局部的重叠,这种排列会减小与周围环境的接触面积,并可能增大物质交流的阻力(图3-E)。
3 讨论与结论
库端的韧皮部卸载是同化物运输的关键环节,同化物卸载速率是库强的综合反映,在整个植物光合同化物分配中占有重要地位[16]。扬麦16和扬麦15同化物卸载速率的差异可较好地反映在灌浆速率上[3,14]。颖果背部维管束是同化物进入籽粒的最后一步,从结构决定功能的观点出发,扬麦16的韧皮部结构应当与其灌浆快的特点相适应。本研究发现,扬麦16韧皮部筛管和周围薄壁细胞较早发生形态改变,细胞壁向内或向外突出导致细胞膜表面积的增大,从而更利于细胞间的物质运输;韧皮部与合点之间的薄壁细胞较早压缩变形,缩短了同化物卸出后到合点和珠心突起的运输距离,细胞间隙较大、合点区液泡中丹宁类物质部分阻塞共质体运输途径后,运输速度更快的质外体途径所占比例相对增大。
于晓刚等认为,颖果背部维管束内的导管数和筛管数决定了维管束的运输能力,维管束越发达,导管和筛管越多,越有利于粒内同化物的运输[2]。但笔者发现,扬麦15的韧皮部筛管较多,管径也较大,细胞形状至花后25 d仍能保持得比较好,直观上比扬麦16略有优势,但灌浆速率、千粒质量[13]等表现却相反。前期研究还表明,扬麦16的维管束、合点、珠心突起传递细胞等母体养分运输组织以及糊粉层传递细胞、胚乳传递细胞等十分发达,共同促进了同化物高效运输[3]。关于维管束对灌浆的影响,人们观点并不一致。李绍长等认为,高等植物光合同化物的运输受韧皮部维管束发育状况影响较大,有时会限制产量[17]。但也有研究发现,维管束的输送功能较强,对同化物的运输分配一般没有限制[18];在同化物供应充足的条件下,维管束的阻力不影响同化物分配[19]。笔者认为,一般情况下维管束可以运输足够的灌浆物质,韧皮部卸载的可溶性糖转化成淀粉的能力对灌浆速率影响更大。因此,正常情况下维管束是否会成为灌浆的限制因素,还须要以更广泛的材料进行深入研究。
木质部位于韧皮部与胚乳腔之间,由韧皮部卸载流出的糖和水分必须先通过木质部才能进入胚乳,因此木质部尤其是导管的作用值得重视和深入研究。截至目前,谷物籽粒脱水在玉米中报道较多,但局限在田间表型性状的观察记载以及分子标记等研究,对籽粒脱水路径的形态解剖以及生理研究在国内外均未见详细报道[20-23]。通过对比扬麦16与扬麦15这2个脱水速率差异显著品种的木质部特征,有一些有意思的发现,即扬麦16在颖果发育早期和中期的导管数量和大小等指标并不优于扬麦15,而在花后25 d,扬麦16的导管结构比扬麦15完整,导管内径和细胞间隙更大,且由于周围薄壁细胞的变形挤压而呈线形排列,按照前人观点[11-12],这种特征使周围环境中多余的水分更容易进入导管而排出。为了进一步明确颖果背部木质部的功能和意义,笔者围绕小麦颖果中的水分运输动态和脱水路径开展相关研究,研究结果将有助于阐明禾谷类作物颖果以及无气孔果实中水分进出的机理问题。
