福建沿海沙地4种防护林类型的群落学特征
2018-02-10林爱玉
林爱玉
(宁德市林业局,福建 宁德 352100)
沿海防护林具有防风固沙、保持水土、涵养水源等功能,对沿海地区防灾、减灾和维护生态平衡具有重大意义.木麻黄(Casuarinaequisetifolia)因具有较强的抗逆和环境适应能力,在改善生态环境、防风固沙等方面发挥了重要作用[1-4].但因为长期多代连栽,木麻黄老林带出现了林木生理衰退、地力下降、生态系统抗逆性差等问题,致使2、3代更新极其困难,防护效能不断下降[5-7].因此,增加海岸带防护林的物种多样性,实现可持续经营,已成为沿海防护林体系建设的当务之急.研究[8]表明,在沿海沙地上通过引种竹子来提高防护林生态系统多样性是推进海岸带防护林可持续经营的有效措施.已有大量研究对非沿海地带竹林的群落特征进行了报道[9-12],对沿海沙地竹林的研究主要集中在养分[13-15]、生物量[16,17]、生理生化[18,19]、地表径流[20]、叶绿素荧光特性[21,22]等,但针对引种竹子后形成的防护林群落特征则鲜见报道.防护林群落与结构特征是评价防护林生态系统稳定性的基本指标,开展防护林群落结构特征的定量分析与评价,有助于进一步理解群落结构与功能的关系[23].基于此,本文对沿海沙地不同林带引种竹子后形成的竹林群落特征进行了研究,旨在进一步了解防护林群落结构变化、防护林营造、生态恢复以及潜在的植被防风固沙性,为沿海沙地物种多样性和生态系统稳定性、可持续性研究提供依据.
1 材料与方法
1.1 研究区概况
研究区位于福建省沿海南部漳州东山赤山国有林场,北纬23°40′,东经118°18′,属于亚热带海洋性季风气候地区.该区干湿季节明显,年均降水量为945 mm,多集中在5—9月,其中11月至翌年2月为旱季,年均蒸发量为1 056 mm,年均气温20.8 ℃,极端最高和极端最低气温分别为36.6和3.8 ℃.台风和干旱为该区主要自然灾害,其中台风多发于夏秋7—8月,年均5.1次.土壤属沿海沙土,肥力较差.天然植被零落,林下零星分布有木豆(Cajanuscajan)、鼠刺(Spinifexlittoreus)和牡荆(Verbenanegando)等[1].
1.2 防护林类型
选择沿海沙地相同方位上、具有代表性的的4种防护林,分别为木麻黄纯林、竹子与木麻黄混交林(简称竹木混交林)、绿竹纯林、各竹种混交林(混交竹林),进行样地设置.4种防护林皆为人工林,其中竹种为2001—2002年引种,皆为丛生竹;木麻黄为2001年种植.各群落基本情况详见表1.

表1 防护林群落基本情况Table 1 Basic information of sample plots
1.3 样方设置及数据调查
在4种防护林里分别设置3块20 m×20 m的固定样地,并将每个样地分成16个5 m×5 m的小样方,调查各小样方内乔木、灌木、草本的种类及数量,记录其高度、冠幅、胸径(灌木草本调查地径)等指标,并以植树高度6 m作为划分乔木层和灌木层的依据.
1.4 参数计算
1.4.1 重要值(Ⅳ) Ⅳi=RAi+RFi+RDi[24].其中,RAi为相对多度、RFi为相对频度、RDi为相对优势度.
1.4.2 多样性指数 选择Simpson多样性指数(DS);均匀度指数选择Simpson均匀度指数(E);丰富度指数选择Gleason指数(D);生态优势度选择Simpson优势度指数(C).以下是群落以及群落各层物种多样性指数的计算方法[25-28].
(1)
(2)
D=S/lnA
(3)
(4)
式中,N为群落中全部物种的个体数;Ni为种i的个体数;S为物种数;A为样地面积;Pi为第i物种所占比例.
2 结果与分析
2.1 4种防护林群落科属构成
4种林带群落科属构成分析见表2,由表2可知,混交竹林科属构成最复杂,植物种类最多,计20科38属41种,以豆科Leguminosae、竹亚科Bamdusoideae、菊科Compositae、禾本科Gramineae等为主;绿竹纯林植物种类为17科28属36种,以豆科Leguminosae、竹亚科Bamdusoideae、菊科Compositae等为主;竹木混交林植物种类为12科18属28种,以竹亚科Bamdusoideae、菊科Compositae、樟科Lauraceae等为主;而木麻黄纯林植物种类最少,计7科10属10种,科属构成较为简单,主要涵盖木麻黄科Casuarinaceae、锦葵科Malvaceae、菊科Compositae等.

表2 不同群落科属构成的调查情况Table 2 Composition of family and genus in different communities
2.2 4种防护林群落种群的数量特征
4种防护林群落主要树种重要值见表3.群落内部垂直结构分化明显,木麻黄纯林、绿竹纯林大致分为乔木层、灌木层、草本层3个层次;竹木混交林、混交竹林大致可分为乔木层、灌木层、草本层、藤本层4个层次.

表3 沿海沙地不同防护林群落主要树种重要值Table 3 Importance value of main population in different communities in coastal sandy land
续表3

层次种名科名 属名 重要值木麻黄纯林竹木混交林绿竹纯林混交竹林草本层黄鹌菜Youngiajaponica菊科 黄鹌菜属 77.276.03-24.14剪刀股Ixerisdebilis菊科 苦荬菜属 47.87--10.32肖梵天花Urenalobata锦葵科 梵天花属 46.87-1.682.93藿香蓟Ageratumconyzoides菊科 藿香蓟属 -195.6246.4817.29韩信草Scutellariaindica唇形科 黄芩属 -52.67-8.22牛膝菊Galinsogaparviflora菊科 牛膝菊属 -23.2744.0757.53牛筋草Eleusineindica禾本科 穇属 -11.077.5114.43铺地黍Panicumrepens禾本科 黍属 -9.84159.3273.86小蓬草Conyzacanadensis菊科 白酒草属 -1.517.518.14狼把草Bidenstripartita菊科 鬼针草属 --9.229.40龙葵Solanumnigrum茄科 茄属 --2.48-母草Linderniacrustacea玄参科 母草属 --11.304.21三叶委陵菜Potentllafreyniana蔷薇科 委陵菜属 --7.19-笔管草Equisetumdebile木贼科 木贼属 ---10.84长尾复叶耳蕨Arachniodessimplicior鳞毛蕨科复叶耳蕨属---16.35狗尾草Setariaviridis禾本科 狗尾草属 ---2.97铁线蕨Adiantumcapillusveneris铁线蕨科铁线蕨属 ---34.80五节芒Miscanthusfloridulus禾本科 芒属 ---2.37藤本层雀梅藤Sageretiathea鼠李科 雀梅藤属 -300.00--羊角藤Morindaumbellata茜草科 巴戟天属 ---251.32
在4种防护林乔木层中,以混交竹林出现的物种数最多,共出现8种;其次是竹木混交林和绿竹纯林,物种数均为5;木麻黄纯林物种数最少,仅1种.木麻黄纯林和竹木混交林均以木麻黄的重要值最大,分别为300、226.21,说明木麻黄在此2种林带群落中处于明显的优势地位.绿竹纯林和混交竹林以绿竹和大头典竹的重要值较大,分别为265.39、110.93.重要值较大的还有竹木混交林的大头典竹(45.10)、混交竹林的绿竹(97.43)、麻竹(32.36)、福建酸竹(23.70).
灌木层中,以竹木混交林出现的物种数最多,共出现20种,其次为混交竹林、绿竹纯林、木麻黄纯林,分别有17、15、7种.其中绿竹纯林灌木层偶见种倒卵叶算盘子、山芝麻,重要值均小于2.木麻黄纯林和竹木混交林均以木麻黄的重要值最大,分别为64.04、74.38;竹木混交林中青竿竹、花孝顺竹、厚荚相思、鼓节竹的重要值均较大,分别为48.45、35.44、34.13、32.63;绿竹纯林以早竹重要值最大,为92.96,其次是花竹,为89.9.混交竹林以厚荚相思重要值最大,为90.36;其次是马樱丹,为84.96;黄甜竹的最小,为31.6.
草本层中,以混交竹林出现的物种数最多,共16种;其次是绿竹纯林、竹木混交林、木麻黄纯林,分别有12、7、3种.且1年生草本和多年生草本物种数从多到少依次为混交竹林、绿竹纯林、竹木混交林、木麻黄纯林.木麻黄纯林和竹木混交林中1年生草本植物重要值最大,黄鹌菜的重要值为77.27,藿香蓟的为195.62;绿竹纯林和混交竹林中多年生草本铺地黍的重要值最大,分别为159.32、73.86.且4种林带中,仅木麻黄纯林有1年生草本,未见多年生植物.
藤本层中,仅竹木混交林和混交竹林有层间藤本植物,其中竹木混交林只有1种雀梅藤,混交竹林由雀梅藤和羊角藤组成,2种林带均由雀梅藤占主导地位.
2.3 4种防护林群落物种多样性
从表4可知,混交竹林群落的物种数最多,达43种;绿竹纯林与竹木混交林的物种数分别为35和33种;木麻黄纯林物种数最少,仅为11种.但竹木混交林个体数最多,为7 299;绿竹纯林和混交竹林个体数分别为4 209、4 183;木麻黄纯林个体数最少,为445.
从群落物种多样性指数看,丰富度指数、辛普森指数和均匀度指数均以混交竹林的群落值最大,且其生态优势度最小,表明混交竹林群落物种多样性最显著且树种分布最均匀.其次是绿竹纯林,丰富度指数、辛普森指数和生态优势度指数都仅次于混交竹林,优于竹木混交林和木麻黄纯林,但绿竹纯林均匀度指数低于木麻黄纯林,物种分布均匀性不及木麻黄.此外,竹木混交林丰富度指数和生态优势度指数均高于木麻黄纯林,但辛普森指数和均匀度指数低于物种数较少的木麻黄纯林,呈现出木麻黄树种在沿海沙地群落中的优势性.总的来说,各群落物种多样性指数大小顺序为混交竹林>绿竹纯林>竹木混交林>木麻黄纯林.
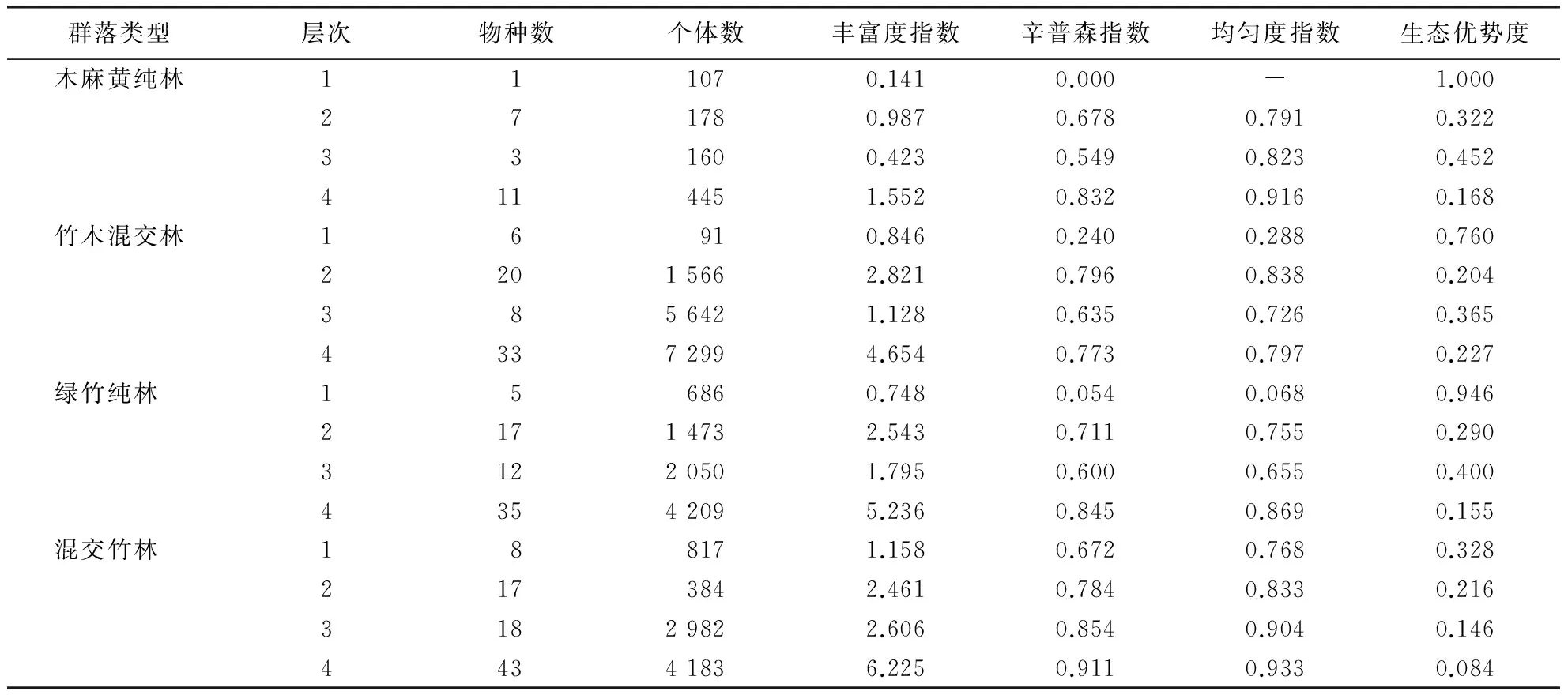
表4 物种多样性指数1)Table 4 Species diversity indices
1)层次中,1表示乔木层(高度≥6 m的乔木和灌木种),2表示灌木层(高度<6 m的乔木和灌木种),3表示草本层,4表示群落总体.
各群落各层次物种多样性各指数表现出不同的规律,其中混交竹林群落各层次物种多样性表现为草本层>灌木层>乔木层,这与亚热带常绿阔叶林物种多样性的格局[29]显然不同;其余3种群落各层次物种多样性均表现为灌木层>草本层>乔木层,这与邹双全等[30]对杉木人工林群落各层物种多样性规律研究结果一致.
3 讨论
4种群落成层现象明显,主要为乔木层、灌木层、草本层,竹木混交林和混交竹林群落还有少量的层间藤本层植物,木麻黄是木麻黄纯林和竹木混交林群落的优势种,而绿竹和大头典竹分别是绿竹纯林和混交竹林群落优势种.
混交竹林群落科属构成最复杂,计20科38属41种,且其优势度最小,均匀度最高,树种分布均匀;其次,依次是绿竹纯林(17科28属36种)、竹木混交竹林(12科18属28种),而木麻黄纯林林带群落科属构成最简单(7科10属10种),但其生态优势度值最大,均匀度值最小,主要由木麻黄树种组成.
总体上,各群落物种多样性指数大小顺序为混交竹林>绿竹纯林>竹木混交林>木麻黄纯林.混交竹林群落各层次物种多样性表现为草本层>灌木层>乔木层,其余3种群落各层次物种多样性均表现为灌木层>草本层>乔木层.混交竹林群落草本层多样性指数高是由于大头典竹和绿竹等混交后林下透光度较其他群落大.
沿海沙地生态环境脆弱,许多学者针对木麻黄地力衰退、林带更新困难等问题,对木麻黄与厚荚相思、湿地松、青皮以及阔叶树混交进行研究[2,4-7],但多集中于土壤理化性质改良方面.竹子生长快,成材早,产竹笋,有较高生态、经济和社会效益[17].本研究分析了木麻黄与竹林混交,各竹混交林、绿竹纯林与木麻黄纯林的群落结构,表明沿海沙地引种竹子可以为解决当前沿海防护林体系木麻黄连带栽培引发的衰退问题提供参考.同时,考虑到沿海台风特点,可以在沿海前沿营造竹木混交林,后方适当营造混交竹林;在沿海沙地引种竹种选择中,将绿竹、大头典竹、麻竹以及青竿竹、花孝顺竹作为优良选择.
研究[31]表明,长期经营纯竹林容易引起竹林地力衰退和生产力下降,营造竹阔混交有利于维持生态系统平衡、增加凋落物、促进养分循环、增肥改土,防止竹林地力下降[32],且纯竹林也较混交林更容易引发病虫害,不利于群落稳定发展[33].这与本文4种群落结构中,绿竹纯林植物种类数较混交竹林少,以及科属构成复杂度也相应低于混交竹林的结论相一致.木麻黄属喜酸厌碱的树种,对土壤的自肥能力较差,虽然木麻黄属生态位衰退树种,但其在沿海前沿所起的防风效能仍不可忽视[6].本研究中,木麻黄属大乔木树种,在群落中占绝对优势,郁闭度较大,林下光线弱,造成林下植被生长所需的温度、水热等气候因子不足,因此其种类数量都较少,科属构成也最简单.而竹子与木麻黄混交后,群落最顶层存在一定的层次落差,形成小空间的林窗[12]和林隙,易于其他植被的生长.4种群落中混交竹林、绿竹纯林的豆科植物均较多,且为高热值植物[5].豆科植物含有根瘤,能起到较好固氮作用,为其它植被生长提供养分[2].同时,混交竹林和绿竹纯林竹种均以丛生竹为主,为其他植被的引入提供更好的光照条件,有利于阳性植物生长[34].胡砚秋等[35]研究指出,不同光照条件下林下草本植物的多度和丰富度差异不显著,但森林冠层拦截后形成的林下散射光对林下植物分布的影响比直射光显著,林下光环境的略微变化会导致生态位的进一步分化[36].也有学者[37]指出在研究林下植物时不应简单地将光环境划分为林窗或非林窗.刘希珍等[38]研究表明毛竹林在扩展过程中,毛竹胸径和凋落物的N∶P比例会发生变化.本研究的竹子虽然都是丛生竹,但丛生竹在群落后期发展中,其地下鞭根系统和土壤理化性质对植被生长有特殊影响,进而对混交竹林和绿化纯林物种多样性产生影响.
综合分析4种防护林类型的群落学特征,有助于进一步了解在沿海沙地引种竹子对构建稳定性沿海防护林体系格局的现实意义,但各群落凋落物归还量以及土壤理化性质与群落结构特征的综合关系还需要进一步研究.
[1] 林爱玉,涂志华,上官保国,等.沿海沙地引种竹子和木麻黄固沙功能比较研究[J].福建林学院学报,2013,33(1):28-33.
[2] 游月娥.木麻黄混交林防护效能和改土效果研究[J].西北林学院学报,2005,20(4):36-38.
[3] 蒋建立.沿海沙地不同竹林土壤有机碳和土壤呼吸动态研究[D].福州:福建农林大学,2013.
[4] 施成坤.沿海前沿林带更新树种选择研究[J].海峡科学,2010(2):46-47.
[5] 张清海,叶功富,林益明.海岸沙地木麻黄人工林凋落物归还量及其热值动态研究[J].林业科学研究,2006,19(5):600-605.
[6] 陈增鸿.海岸带防护林恢复过程中凋落物的组成及养分特征[D].福州:福建农林大学,2012.
[7] 伍恩华,刘强,王敏英.海南岛北部木麻黄防护林凋落物量及养分归还动态[J].华南师范大学学报(自然科学版),2012,44(2):123-129.
[8] 荣俊冬,郝瑞,祝杰伟,等.沿海沙地竹林细根养分含量及其季节动态[J].福建林学院学报,2007,27(4):313-317.
[9] 颜玉娟,赵虎,杨倩,等.湖南阳明山毛竹林群落特征[J].华中农业大学学报,2010,29(3):375-380.
[10] 张刚华,萧江华,聂洁珠,等.不同类型毛竹林植物物种多样性研究[J].林业科学研究,2007,20(5):615-621.
[11] 孟勇,刘国华,王福升.小蓬竹群落学的特征研究[J].中南林业科技大学学报,2011,31(2):73-78.
[12] 王微,陶建平,胡凯华,等.华西箭竹对岷江冷杉林主要乔木树种幼苗结构及分布格局的影响[J].林业科学,2007,43(1):1-7.
[13] 陈羡德,陈礼光,荣俊冬,等.施肥对沿海沙地麻竹笋期叶片养分动态的影响[J].福建农林大学学报(自然科学版),2007,36(6):585-591.
[14] 郑郁善,荣俊冬,陈礼光,等.沿海沙地小叶龙竹林凋落物分解及养分归还动态[J].福建农林大学学报(自然科学版),2008,37(5):487-490.
[15] 张梅,郑郁善.滨海沙地吊丝单竹林凋落物分解及养分动态研究[J].西南林学院学报,2008,28(3):4-7.
[16] 郑郁善,黄宇,郝瑞,等.沿海沙地竹林细根分解过程中的能量变化[J].福建林学院学报,2009,29(1):1-5.
[17] 陈礼光,郑郁善,姚庆端,等.沿海沙地新造绿竹林生物量结构[J].福建林学院学报,2002,22(3):249-252.
[18] 郑郁善,郑容妹,姚庆瑞.盐分胁迫对沿海沙地绿竹生理生化的效应[J].福建农林大学学报(自然科学版),2003,32(4):478-481.
[19] 涂志华,潘瑞,卢仁锋,等.滨海沙地沿海沙地8个竹种根际土壤的酶活性[J].福建农林大学学报(自然科学版),2012,41(3):243-247.
[20] 郑郁善,陈卓梅,邱尔发,等.不同经营措施笋用麻竹人工林的地表径流研究[J].生态学报,2003,23(3):2 387-2 395.
[21] 涂志华,卢仁锋,潘瑞,等.沿海沙地10个竹种叶绿素荧光特征比较[J].福建林学院学报,2012,32(2):131-135.
[22] 涂志华,潘瑞,洪雪沿,等.滨海沙地簕竹属10个竹种叶绿素荧光特性研究[J].竹子研究汇刊,2012,31(1):52-62.
[23] 李贵祥,孟广涛,方向京,等.滇中高原桤木人工林群落特征及生物量分析[J]. 浙江林学院学报,2006,23(4):362-366.
[24] 蓝良就,黄炎和,李德成,等.退化红壤恢复过程中灌木层主要种群的生态位[J].福建农林大学学报(自然科学版),2012,41(2):176-182.
[25] 朱学灵,刘晓静,崔向慧,等.宝天曼自然保护区栎类林群落不同火烧演替序列物种多样性特征[J].林业科学,2012,48(8):31-39.
[26] 黄承才,张骏,江波,等.浙江省杉木生态公益林凋落物及其与植物多样性的关系[J].林业科学,2006,42(6):7-13.
[27] 董建文,黄启堂,陈世品,等.杉木林迹地近自然恢复苦竹群落的研究[J].中国生态农业学报,2008,16(5):1 218-1 224.
[28] 陈辉.33年生格氏栲人工林与天然林群落特征比较[J].福建林学院学报,2009,29(1):97-102.
[29] 贺金生,陈伟烈,江明喜,等.长江三峡地区退化生态系统植物群落物种多样性特征[J].生态学报,1998,18(4):399-407.
[30] 邹双全,何宗明,陈光水,等.老杉木人工林与松阔混交林群落多样性比较[J].福建林学院学报,2002,22(2):124-128.
[31] 曹永慧,萧江华,陈双林,等.竹阔混交林中阔叶树对毛竹生长的影响及竞争关系[J].浙江林学院学报,2006,23(1):35-40.
[32] 赵永艳,苏继申.竹阔混交林土壤性状与分析[J].南京林业大学学报(自然科学版),2007,31(1):81-84.
[33] 张艳璇,张智强,斋藤裕,等.混交林和纯竹林与毛竹害螨爆发成灾关系研究[J].应用生态学报,2004,15(7):1 161-1 165.
[34] 陈志伟,伊贤贵,王贤荣,等.黄山微毛樱群落主要种群生态位特征[J].南京林业大学学报(自然科学版),2014,38(S),39-48.
[35] 胡砚秋,柯娴氡,徐明锋,等.亚热带森林群落草本植物对林下光环境的响应[J].中南林业科技大学学报,2016,36(8):72-77.
[36] SVENING J. Small canopy gaps influence plant distributions in the rain forest understory[J]. Biotropica, 2000,32(2):252-261.
[37] SCANGA S E. Population dynamics in canopy gaps: nonlinear response to variable light regimes by an understory plant[J].Plant Ecol, 2014,215(8):927-935.
[38] 刘希珍,范少辉,刘广路,等.毛竹林扩展过程中主要群落结构指标的变化特征[J].生态学杂志,2016,35(12):3 165-3 171.
