初级视觉皮层在注意振荡中的作用*
2018-02-04陈艾睿王爱君王天琪唐晓雨
陈艾睿 王爱君 王天琪 唐晓雨 张 明
(1苏州大学心理学系, 心理与行为科学研究中心, 苏州 215123)
(2辽宁师范大学心理学院, 儿童青少年健康人格评定与培养协同创新中心, 大连 116029)
1 引言
一个朋友从远处走来时, 我们看到他自然地经过了沿途的每一个位置, 所以主观意识上觉得注意在时间维度上似乎也是连续的。但实际上, 这种意识的连续性并不能证明注意的连续性。注意可能每隔几十毫秒采集一次信息, 然后构建整体的知觉,同时又保证意识状态的连续性(VanRullen & Koch,2003)。近期, 诸多研究表明, 注意并非连续地而是间歇性地加工外界信息(Busch & VanRullen, 2010;Landau & Fries, 2012; Song, Meng, Chen, Zhou, & Luo,2014; VanRullen, Carlson, & Cavanagh, 2007)。注意的这种特性被称为“注意的离散性”, 指在精细的时间尺度上, 注意间歇性地采集外界信息的特性, 即“采集”、“停止”、“采集”、“停止”……如此循环往复(VanRullen et al., 2007)。该特性在行为上表现为指标(正确率或反应时)有规律地时高时低, 这种由于注意导致的行为指标有规律的变化被称为注意振荡(attention oscillation)。新近研究采用高时间分辨率的行为学研究方法, 打破了传统认知任务不能精细研究注意离散性的局限, 直接在被试的行为中(探测正确率和反应时)发现了振荡模式(Benedetto,Spinelli, & Morrone, 2016; Chen, Wang, Wang, Tang,& Zhang, 2017; Dugué, Marque, & VanRullen, 2015;Dugué, McLelland, Lajous, & VanRullen, 2015; Dugué,Roberts, & Carrasco, 2016; Fiebelkorn, Saalmann, &Kastner, 2013; Huang, Chen, & Luo, 2015; Landau &Fries, 2012; Song et al., 2014)。
虽然研究者在视觉探测(Landau & Fries,2012)、视觉搜索(Dugué, Marque, et al., 2015; Dugué,McLelland, et al., 2015; Dugué & VanRullen, 2014,2017; Dugué, Xue, & Carrasco, 2017)和车轮错觉(Macdonald, Cavanagh, & VanRullen, 2014; VanRullen et al., 2007)等现象中发现并证明了注意振荡的存在, 并采用EEG和MEG发现了神经振荡与注意振荡的密切关系(Busch & VanRullen, 2010; Buschman& Kastner, 2015; Landau, Schreyer, van Pelt, & Fries,2015; VanRullen, Zoefel, & Ilhan, 2014; Zoefel &VanRullen, 2017), 但是, 还存在两方面的问题:第一, EEG和MEG技术存在部分局限(例如, 空间分辨率低、信号失真) (陈艾睿, 唐晓雨, 王爱君, 张明,2017); 第二, 现有研究发现的脑区位置相互冲突。研究使用 EEG技术发现了注意振荡与额中央区的神经活动相关(Busch, Dubois, & VanRullen, 2009;Busch & VanRullen, 2010); 而 Landau 等(2015)通过脑磁图技术探查到对侧半球距状沟、舌回以及楔前叶等脑区的神经振荡与注意振荡密切关联, 神经振荡可能是注意振荡的神经机制(Landau et al.,2015)。诚如 VanRullen (2016a, 2016b)指出, 现有研究并不能说明注意离散性究竟源于脑内的何处位置, 与何种大脑结构相关, 即注意离散性的神经节点并不清楚(VanRullen, 2016a, 2016b)。
由视觉通路的解剖特性可知, 双眼朝前的高等哺乳动物和人的单眼视觉信息是以分离的方式传输到左右两侧视皮层的。在外侧膝状体的六层细胞中, 来自两眼的信息分别独立投射至不同的细胞层内。仅当信息传递至初级视觉皮层(Primary visual cortex, V1)时, 来自双眼的信息才能产生汇聚(Hubel & Wiesel, 1977; 寿天德, 2010)。V1的第四层细胞(IVc)是大脑皮层接收视觉信号的第一站。此处, 从双眼而来的视觉信号仍和外侧膝状体神经元一样, 彼此独立, 每一个单眼细胞仅能接受来自一只眼的信息(同眼条件), 另一眼内的信息对它没有影响(异眼条件)。相比于其他视皮层区域, 单眼细胞仅在V1存在, 这些细胞提供了“信号来自哪一只眼”的信息(Hubel & Livingstone, 1987)。除此之外,V1还包含大量对双眼的信息均有反应的神经元,称为双眼细胞, 只有来自同一感受野输入的单眼细胞信息才汇聚至同一双眼细胞。因此, 若一个现象在同、异眼中有差异, 则说明该现象的神经节点位于初级视觉皮层的 IVc层之前(单眼细胞部分); 若同、异眼没有差别, 则表明该现象的神经节点位于初级视觉皮层的 IVc层之后(双眼细胞部分)。Zhaoping (2008)基于此发现了注意突显地图(bottomup saliency map)产生于V1, 研究还发现, 相比靶子和干扰物同眼呈现, 异眼的情况下被试需要花费更多时间才能找到原来的靶子, 这一差异说明了注意突显地图在V1的单眼细胞加工阶段就已经产生了(Zhaoping, 2008)。此外, 同眼异眼呈现的方法还被广泛用于其他视知觉现象的神经节点的研究中, 如朝向适应(Gilinsky & Doherty, 1969), 空间频率适应(Blakemore & Campbell, 1969), 颜色适应(McCollough,1965), 运动后效(Anstis, Verstraten, Mather, & George,1998), 主观轮廓知觉(Paradiso, Shimojo, & Nakayama,1989), 知觉学习(Schoups & Orban, 1996)等。
诸多研究表明内源性注意和外源性注意与初级视觉皮层(V1)内的神经元激活状态有关(Briggs,Mangun, & Usrey, 2013; Chalk et al., 2010; Pooresmaeili,Poort, & Roelfsema, 2014; Pooresmaeili, Poort, Thiele,& Roelfsema, 2010; Thiele, Pooresmaeili, Delicato,Herrero, & Roelfsema, 2009; Wang, Chen, Yan, Zhaoping,& Li, 2015)。不仅内源性注意可以调控V1区域的神经元激活状态, 外源性注意同样可以增强 V1区域内线索化位置对应的神经元的放电频率(Wang et al., 2015)。Dugué等(2016)使用TMS技术干扰V1/V2, 发现非线索化条件, 即注意重定向时, TMS干扰的区域为靶子加工的脑区和干扰物加工对应的脑区时, 被试的辨别力在时间维度上呈现出周期性变化, 但在线索化条件却未发现振荡模式(Dugué et al., 2016), 虽然这与注意离散性的经典研究结果并不一致, 但研究提示了V1/V2可能也在注意离散性中起到重要作用。
基于视觉通路的解剖特性, 本研究采用高时间分辨率的线索靶子范式, 利用双眼分视技术, 考察V1区在注意离散性中的作用。实验选取平面立体镜达到同眼和异眼呈现的目的。该装置通过反射原理, 让屏幕左侧刺激进入左眼, 右侧刺激进入右眼,以此独立控制单眼呈现的刺激。当线索和靶子出现在不同的眼睛内时(异眼呈现), 二者分别驱动了V1内不同的单眼神经元; 当二者出现在同一只眼睛内时(同眼呈现), 二者驱动了相同的单眼神经元。考虑到辨别任务较探测任务更少受反应偏向的影响(Drewes, Zhu, Wutz, & Melcher, 2015; Dugué, Merriam,Heeger, & Carrasco, 2017; 钱晨灿, 刘祖详, 2016),实验1使用四择一的迫选任务(four-alternative forcedchoice, 4AFC)考察了非双眼分视下注意的振荡, 作为研究的基线条件。实验2则在实验1的基础上操控线索和靶子呈现的眼间关系, 考察同眼呈现、异眼呈现条件下的注意振荡:1)如果仅当同眼呈现时注意振荡才会发生, 说明注意离散性源于 V1的双眼细胞之前的单眼视觉通路之中; 2)如果异眼条件下和同眼条件下都出现了同样的振荡模式, 说明注意离散性源于V1的双眼细胞或之后的视觉区。
2 实验1:非双眼分视条件下的注意振荡
2.1 方法
2.1.1 被试
16名苏州大学本科生参加了实验(男 3名, 女13名), 年龄为20~25周岁(M
=21.19岁,SD
=1.60),均为右利手。所有被试视力或矫正视力正常, 无色盲或色弱。实验后给予被试相应报酬。2.1.2 实验设计
实验1作为基线条件, 考察了非双眼分视条件下的注意振荡。采用了单因素两水平(空间有效性:线索化vs.非线索化)被试内实验设计。根据线索与靶子呈现位置的异同(空间有效性), 分为线索化条件和非线索化条件。线索化条件是指靶子出现在与线索的空间位置相同的光栅之上(同侧位置), 非线索化条件是指靶子出现在与线索的空间位置相反的光栅之上(对侧位置)。
为了探测注意振荡, 实验中设置50个SOA水平, 从100到1080 ms中每间隔20 ms取一个水平(50 Hz采样频率)。实验中, 通过测量每个SOA水平下的正确率, 计算出本实验的因变量注意振荡节律。
从任务类型来看, 探测任务会更受被试主观标准的影响, 如果被试的判断标准更为冒险, 会有较多的错误反应; 而如果标准更为严谨, 会有较少的正确判断, 这种反应偏向会污染最终的实验结果。研究表明, 辨别任务相对较少受到判断标准的影响(Drewes et al., 2015)。本实验优化了注意振荡的实验方法, 将任务改为四择一的迫选任务(4AFC), 并且该任务为辨别任务, 被试需要尽量准确分辨靶子刺激的位置(左上、左下、右上、右下), 对反应速度没有要求。
2.1.3 实验仪器
实验程序采用Matlab和Psychophysics Toolbox-3编写(Brainard, 1997; Pelli, 1997), 在Dell Optiplex755计算机上运行, 显示器为22英寸ViewSonic P225f CRT, 分辨率为1024×768, 刷新频率为100 Hz, 用键盘进行按键反应。
2.1.4 实验刺激
实验场景和参数参考Landau和Fries (2012)的研究(详见图1A)。所有刺激均呈现在一个灰色背景之上, 灰色背景的亮度是 3.88 cd/m。被试需要始终盯住一个白色圆点(直径为 0.5°), 此为中央注视点。同时, 被试需要监视着左右两侧的光栅, 光栅直径为4°, 光栅距离中央注视点距离(离心率)为5°,光栅的空间频率为1.4 c/°, 对比度为100%。为了尽量减少靶刺激的突显性影响注意振荡, 光栅并非静止而是沿着某一方向运动, 每个试次中, 两个光栅运动速度均为0.7 c/s (Landau & Fries, 2012)。同时为防止视觉系统适应光栅朝向, 运动方向分别独立从 0°~360°中随机选取, 光栅朝向与运动方向垂直(Landau & Fries, 2012)。同时, 每个光栅分别被灰色十字线(线宽0.22°)平均分为4份:左上、右上、左下、右下。
线索刺激包含4个白色圆点, 圆点的直径为1°,圆点中心距离光栅边缘 1.5°。实验中, 线索呈现的时间为30 ms。靶刺激是指光栅上任一圆形区域内的对比度陡然下降, 圆形区域直径为1°。同样为了降低靶刺激的突显性, 整个圆形区域内对比度下降的幅度呈高斯分布, 此分布的标准差(SD
)为 0.5°,最大值由一个自适应阶梯法的程序(QUEST)确定(Watson & Pelli, 1983)。靶刺激的呈现时间为30 ms,靶刺激随机出现在光栅4个区域中的任意一个之中,避免与十字线重叠。2.1.5 实验流程
本实验在暗室内进行, 被试端坐在屏幕前方,距离屏幕50 cm, 头放在颚托架上。为更好地探测注意振荡, 实验首先采用 QUSET方法为每一名被试确定了一个合适的难度, 此时被试探测到靶刺激出现的概率为 50% (阈限测量) (Fiebelkorn et al.,2013; Landau & Fries, 2012); 然后考察线索化和非线索化条件被试探测视觉刺激能力的周期性变化(振荡测量)。
靶刺激的明显程度事先在阈限测量阶段确定。除不呈现线索刺激之外, 实验刺激和流程与振荡测量阶段相同。程序根据被试的反应自动调节靶子位置的对比度下降的数值。下降得越多靶刺激越明显,下降得越少靶刺激越微弱。如果被试可以正确判断位置, 则在随后一次的测试中减少对比度的下降程度, 反之, 判断错误则升高对比度的下降值。通过这种方法测出被试探测到靶刺激的合适阈值, 即, 此时被试有50%的机率可以正确探测出靶刺激的位置。
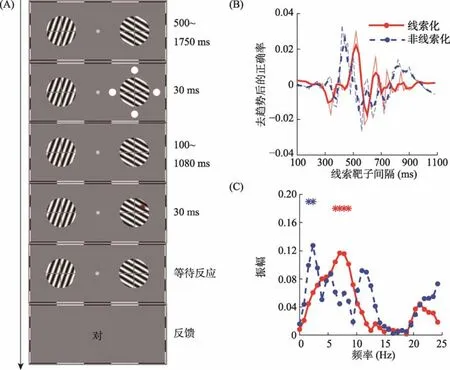
图1 实验1流程和注意振荡结果。(A)实验1流程图; (B)在补零扩展、去趋势、汉宁窗滤波之后, 通过快速傅里叶变换把行为振荡中的时域变换到频域。图中显示了线索化条件(红色实线)和非线索化条件(蓝色虚线), 被试的正确率随时间的变化趋势; (C)线索化条件和非线索化条件各频段的振幅, *表示显著的频段。彩图见电子版,下同。
振荡测量阶段的流程(详见图1A):每个试次中,空屏呈现1000至1200 ms后, 出现两个运动光栅,随后(即, 光栅呈现 500~1750 ms后)线索刺激出现在其中一个光栅的周围(30 ms), 间隔100至1080 ms之后, 靶刺激呈现30 ms后消失, 屏幕中继续呈现注视点和运动光栅等待被试按键反应。此时, 被试需要完成一个 4AFC任务:判断靶刺激的位置(即,靶刺激出现在光栅的哪个方位)。若3 s内被试仍不按键报告, 则呈现红色提示语“左下、右下、左上、右上”, 让被试猜测刚才靶刺激出现在光栅上的哪个方位, 直至被试按键, 该试次结束。为了减少被试按键难度, 我们选择键盘中小键盘部分的“1、2、4、5”四个按键分别对应光栅上的四个部分。需要注意的是, 被试可以忽略靶刺激具体出现在哪个光栅之中, 而仅仅需要判断靶刺激出现在光栅上的哪个方位。被试按键后, 通过在屏幕中心呈现“对”或“错”字样给予反馈。振荡测量阶段, 每名被试共完成1600个试次, 其中线索化条件有50种SOA水平,非线索化条件有50种不同的SOA水平, 每种条件重复16次。为了避免疲劳效应, 振荡测量阶段分为8组, 在2~3天内完成。
2.2 数据分析
采用Matlab和CircStat工具箱分析被试的正确率(Berens, 2009)。对于每名被试, 首先计算出每个SOA线索化与非线索化条件下的正确率, 接下来,将正确率依照SOA从100至1080 ms分线索化和非线索化进行排列, 由此得到被试的探测能力随时间的变化规律(ACC-SOA信号), 即行为振荡。接下来, 为了分析行为振荡的频谱特性, 对每名被试的行为振荡(即, ACC-SOA信号)进行频谱分析。具体来看, 每种条件下, 在补零扩展(Zero padded)、去趋势(Detrend)、汉宁窗滤波(Hanned)之后, 通过快速傅里叶变换(fast Fourier transform, FFT)把行为振荡中的时域变换到频域。由此, 便可探知行为振荡中的振荡信息。除了频率信息以外, 为了进一步确定振荡模式, 计算了线索化与非线索化条件下行为振荡的相位信息。在傅里叶变换中提取每名被试在每个频率(0~25 Hz)上的相位信息, 随后将线索化与非线索化条件下的相位相减, 得到二者的相位差(Phase difference), 最后计算出所有被试的相位趋势(Cross-subject coherence in the phase difference values)。由此, 确定每个频率下线索化与非线索化条件的相位关系, 并采用圆形统计(Circular statistics)中的 Rayleigh tests检查显著频段的相位信息的不一致性是否达到显著水平。
为了探测哪些频率的振荡达到显著水平, 采用以下非参数统计方法。首先, 在每个被试每种条件下, 随机打乱ACC-SOA信号中的时间信息1000次,产生1000个替代信号(Surrogate signals)。对每个替代信号, 进行上述的FFT分析, 在每个频率上得到1000个替代信号对应的振幅, 这些振幅组成了频域信息的置换分布(Permutation distribution)。采用置换检验(Permutation test)对置换分布和原始行为振荡中的振荡信息进行分析, 得到每个频率(0~25 Hz)振荡的显著性。由于该置换检验中涉及了多重比较, 采用较为严格的Bonferroni法对结果进行矫正, 以防止假阳性结果的出现。
2.3 结果
首先, 实验 1探究了非双眼分视条件下 4AFC任务中注意振荡的频率。结果发现, 线索化和非线索化条件下被试的正确率无差别, 分别为 62.04%(SD
=8.68%)和55.01% (SD
=16.00%), 配对样本t检验发现二者无显著差异,t
(15)=1.92,p
=0.074,d
=0.48, 95%CI=[−0.0077, 0.15]。更重要的是线索化位置和非线索化位置的正确率在时程上呈现此起彼伏的模式。具体见图1B, 图中红色实线代表线索化位置正确率随时间的变化, 蓝色虚线代表非线索化位置正确率随时间的变化。细线表示原始正确率, 对其进行三点平滑之后得到粗线。结果发现,当 SOA较短时线索化位置的正确率高于非线索化位置, 但是随着SOA的延长, 这种趋势并非一直保持, 而是数次发生反转, 出现“此起彼伏”的交替现象, 该现象称为行为振荡。此振荡则定性地表明注意在线索化位置与非线索化位置来回切换, 证明4AFC的范式仍旧能够探测到注意振荡。通过傅里叶分析和置换检验发现, 行为振荡在某些频段达到显著水平。具体见图1C, 线索化位置的振荡在6.25~8.59 Hz达到显著水平(p
< 0.05), 非线索化位置在1.56~2.34 Hz (p
< 0.05)达到显著水平。此结果进一步表明实验1中注意振荡出现在低频频段, 与前人结果一致(Dugué, Marque, et al.,2015; Fiebelkorn et al., 2013; Landau & Fries, 2012)。3 实验2:双眼分视条件下的注意振荡
3.1 方法
3.1.1 被试
16名苏州大学本科生参加了实验(男 5名, 女11名), 年龄为20~25周岁(M
=20.75岁,SD
=1.43),均为右利手。所有被试视力或矫正视力正常, 无色盲或色弱。实验后给予被试相应报酬。3.1.2 实验设计
实验2考察了分视条件下的注意振荡。采用了2(空间有效性:线索化 vs.非线索化)×2(眼睛有效性:同眼vs.异眼)的被试内实验设计。空间有效性的操作定义与实验1相同。根据线索与靶子呈现眼睛的异同(眼睛有效性), 分为同眼条件和异眼条件。同眼条件是指线索与靶子出现在同一只眼睛内,异眼条件是指线索与靶子出现在不同的眼睛内。
3.1.3 实验仪器
利用平面立体镜把电脑屏幕两侧图像分别反射到被试的左眼和右眼。其余设备与实验1相同。
3.1.4 实验刺激
与实验1相同。
3.1.5 实验流程
首先, 通过调试程序让每名被试调整平面立体镜成像直至两侧图像很好地在中央位置融合。然后进行与实验 1相同的阈限测量和注意振荡测量实验。图2显示了异眼条件的一种情况, 即线索出现在右眼, 而靶子出现在左眼。
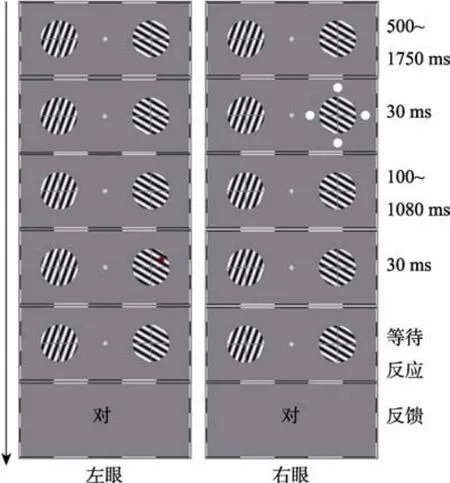
图2 实验2流程(异眼条件)
3.2 数据分析
与实验1中的数据处理与分析方法相同。
3.3 结果
实验2采用4AFC范式探究同眼和异眼下注意振荡的规律。结果发现, 线索化和非线索化条件下被试的正确率无差异, 分别为 52.53% (SD
=11.76%)和48.37% (SD
=14.13%), 配对样本t检验发现二者无显著差异,t
(15)=1.82,p
=0.09,d
=0.23, 95%CI=[−0.0072, 0.090]; 同眼线索化条件和非线索化条件下被试的正确率无差异, 分别为52.27% (SD
=12.03%)和 47.63% (SD
=14.56%), 配对样本t检验发现二者无显著差异,t
(15)=2.02,p
=0.06,d
=0.25, 95%CI=[−0.0025, 0.095]; 异眼线索化条件和非线索化条件下被试的正确率无差异, 分别为52.80% (SD
=11.67%)和49.13% (SD
=13.91%),配对样本t检验发现二者无显著差异,t
(15)=1.53,p
=0.15,d
=0.20, 95%CI=[−0.014, 0.088]。通过傅里叶分析和置换检验发现, 无论是同眼还是异眼条件下, 都出现了明显的行为振荡模式(详见图3A和C)。对行为振荡进一步分析发现, 两种条件下都发现了显著的振荡频段。具体来看, 同眼条件下, 线索化位置和非线索化位置都在12.50~13.28 Hz达到了显著水平(p
< 0.05) (详见图3B); 异眼条件下, 线索化位置的行为振荡同样在12.50~13.28 Hz达到了显著水平(p
< 0.05), 非线索位置的行为振荡则在11.72~13.28 Hz达到了显著水平(p
< 0.05) (详见图3D)。由此可知, 在线索化与非线索化位置观察到了明显的行为振荡。为了进一步确定实验2中的行为振荡, 对同眼和异眼条件下 12~13 Hz的相位进行分析, 发现两种条件下, 线索化位置与非线索化位置的相位出现了显著差异(Reyleigh test, 同眼条件p
=0.012, 异眼条件p
=0.014), 且差异在180°附近(详见图 4), 即线索化与非线索化两个位置出现了精准的反相位振荡模式。
图3 实验2注意振荡结果。(A)在补零扩展、去趋势、汉宁窗滤波之后, 通过快速傅里叶变换把行为振荡中的时域变换到频域。线索靶子出现在同眼时, 线索化和非线索化条件下, 正确率随时间变化的趋势; (B)线索靶子出现在同眼时, 线索化条件和非线索化条件各频段的振幅; (C)线索靶子出现在异眼时, 线索化和非线索化条件下, 正确率随时间变化的趋势; (D)线索靶子出现在异眼时, 线索化条件和非线索化条件各频段的振幅, *表示显著的频段。
4 讨论
本研究采用高时间分辨率的线索靶子范式, 利用平面立体镜分视, 操纵了被试双眼视像, 不仅考察了双眼分视是否会影响注意振荡(实验1 vs.实验2), 还进一步考察了线索和靶子呈现在同眼 vs.异眼对注意振荡的影响(实验 2), 以探究初级视觉皮层V1区在注意振荡中的作用。研究结果发现:1)非双眼分视条件下(实验 1), 注意振荡出现在低频(2 Hz和8 Hz), 而分视条件下, 注意振荡则出现在更高的频段(12.5 Hz)。这表明分视会对注意振荡发生的频段产生影响。2)线索靶子出现在同眼还是异眼条件下注意振荡无显著差异。这提示V1区双眼视觉通路或者更高级的视觉区可能参与注意振荡。

图4 实验2中同眼(A)和异眼(B)条件下注意振荡的相位结果。
实验1采用四择一的辨别任务测量了注意振荡,结果发现注意振荡出现在低频频段, 与以往结果一致(Dugué, Marque, et al., 2015; Fiebelkorn et al.,2013; Landau & Fries, 2012)。值得注意的是, 相较于探测任务, 本研究采用的辨别任务可以更好地避免被试反应标准(激进或保守)的影响(Drewes et al.,2015; Dugué, Merriam, et al., 2017; 钱晨灿, 刘祖详, 2016)。测得的正确率更纯净地反映了被试的注意或者知觉能力的变化, 而非决策标准。除此之外,在辨别任务中, 常见的2AFC的辨别任务的随机正确率为50%, 仅能操控50%~100%的阈限。而4AFC的随机正确率为25%, 此时实验者可以操控的阈限范围是25%~100%。此优势有利于研究不同任务难度情况下, 行为振荡模式的不同。
近年来, 注意离散性领域的研究结果, 极大地挑战了传统的注意聚光灯理论, 注意并非连续的而是离散的, 基于此, VanRullen等人提出“眨眼聚光灯”理论, 认为注意系统在加工外界信息时是有节律的, 当空间中存在多个物体时, 注意有规律地进行切换, 而当仅加工单个物体时, 注意也是离散加工(VanRullen, 2013, 2016a, 2016b; VanRullen et al.,2007, 2014)。本研究的结果进一步支持了“眨眼聚光灯”理论, 而且发现双眼分视与否(实验 1vs.实验2)会影响到注意振荡的频段。值得注意的是, 如果仅在一只眼中呈现刺激, 人类被试不能够区分该刺激源自于哪只眼睛, 个体意识到的是两只眼睛内图像的叠加状态(Wolfe & Franzel, 1988)。V1被认为是脑皮层中最不可能与意识相关的脑区(Crick &Koch, 1995; He, Cavanagh, & Intriligator, 1996; He& MacLeod, 2001)。所以, 双眼分视和非双眼分视条件产生的意识状态相同, 两种条件的差异不可能源于各种意识上的认知因素。Jia, Liu, Fang和Luo(2017)使用高时间分辨率的脑电技术, 在各种不同的任务中考察了注意的时空动态特征。他们发现:1)刺激出现后, 头皮表面记录到的抑制性 alpha波标识了注意指向的空间位置或者客体, 约 200 ms注意会在物体间切换一次; 2)注意时空动态特征会受到任务调控, 且观察到脑电的趋势与行为表现相关, 结果说明注意不仅会对焦点处物体进行加工,在焦点外还会监控其他物体, 这一过程是以动态系列采样的模式工作的(Jia et al., 2017)。这篇研究及本研究结果都支持了注意的离散性, 提示注意以系列的方式进行工作, 且其动态的工作模式会受到任务类型及刺激呈现眼间条件的影响而进行灵活调控。
研究采用高时间分辨率的行为学方法证实了视觉注意的离散性, 并且可以体现在正确率、反应时两个指标之上(Landau & Fries, 2012; Song et al.,2014); 电生理研究证明神经振荡可能是注意离散性的神经机制(Busch & VanRullen, 2010; Landau et al., 2015)。但是, 现有研究并不能说明注意离散性究竟源于脑内的何处位置, 与何种大脑结构相关(VanRullen, 2016a, 2016b)。本研究利用人眼视觉通路的解剖学特性, 采用双眼分视技术, 比较了线索靶子出现在同眼和异眼两种条件下的注意振荡。同眼条件, 靶子激活的是线索出现时激活的单眼视觉通路, 而异眼条件下, 靶子激活的是另外一只眼睛的单眼视觉通路。结果发现, 即使激活的单眼视觉通路发生变化, 注意振荡发生的频段并未改变, 这提示 V1区的单眼视觉通路可能并未参与注意振荡。Dugué等(2016)采用 TMS技术大范围干扰V1/V2区时发现这些区域会对非线索化条件的正确率产生周期性变化(Dugué et al., 2016), 结合本研究的结果共同提示, 注意振荡可能与 V1区的双眼视觉通路或者更高级的视觉区域有关。初级视觉皮层注意突显图理论指出, 人类的初级视觉皮层可以在视觉信息加工的早期阶段生成视觉突显图, 用以引导空间选择性注意的分布(Zhaoping, 2008)。这一理论得到了fMRI、ERP研究结果的支持, V1区的活动可以预测自下而上的注意及分布(Zhang,Zhaoping, Zhou, & Fang, 2012), 而本研究的结果发现 V1区还参与注意离散性的表达, 但额顶区是注意系统工作的重要脑区(Buschman & Kastner, 2015;Buschman & Miller, 2009, 2010), 因此未来研究需要厘清视觉区是注意离散性产生的直接原因, 抑或是额顶区通过反馈信号使得初级视觉皮层进而表现出注意离散的特性。
5 结论
(1) 四择一迫选的辨别任务可以探测到明显的注意振荡。
(2) 双眼分视和非双眼分视条件的注意振荡模式存在差异, 表明分视会对注意振荡产生影响。
(3) 同、异眼条件的注意振荡模式无差异, 表明V1区双眼视觉通路或者更高级的视觉区可能参与注意振荡。
致谢:
感谢中科院心理所董波、袁佩君在数据分析过程中提供的建议和帮助。Anstis, S., Verstraten, F.A.J., & Mather, G.(1998).The motion aftereffect.Trends in Cognitive Sciences, 2
(3), 111–117.Benedetto, A., Spinelli, D., & Morrone, M.C.(2016).Rhythmic modulation of visual contrast discrimination triggered by action.Proceedings of the Royal Society B:Biological Sciences, 283
(1831), 20160692.Berens, P.(2009).CircStat: A MATLAB toolbox for circular statistics.Journal of Statistical Software, 31
(10), 1–21, doi:10.18637/jss.v031.i10.Blakemore, C., & Campbell, F.W.(1969).On the existence of neurones in the human visual system selectively sensitive to the orientation and size of retinal images.Journal of Physiology, 203
(1), 237–260.Brainard, D.H.(1997).The psychophysics toolbox.Spatial Vision, 10
(4), 433–436.Briggs, F., Mangun, G.R., & Usrey, W.M.(2013).Attention enhances synaptic efficacy and the signal-to-noise ratio in neural circuits.Nature, 499
(7459), 476–480.Busch, N.A., Dubois, J., & VanRullen, R.(2009).The phase of ongoing EEG oscillations predicts visual perception.Journal of Neuroscience, 29
(24), 7869–7876.Busch, N.A., & VanRullen, R.(2010).Spontaneous EEG oscillations reveal periodic sampling of visual attention.Proceedings of the National Academy of Sciences of the United States of America, 107
(37), 16048–16053.Buschman, T.J., & Kastner, S.(2015).From behavior to neural dynamics: An integrated theory of attention.Neuron,88
(1), 127–144.Buschman, T.J., & Miller, E.K.(2009).Serial, covert shifts of attention during visual search are reflected by the frontal eye fields and correlated with population oscillations.Neuron, 63
(3), 386–396.Buschman, T.J., & Miller, E.K.(2010).Shifting the spotlight of attention: Evidence for discrete computations in cognition.Frontiers in Human Neuroscience, 4
, 194.Chalk, M., Herrero, J.L., Gieselmann, M.A., Delicato, L.S.,Gotthardt, S., & Thiele, A.(2010).Attention reduces stimulus-driven gamma frequency oscillations and spike field coherence in V1.Neuron, 66
(1), 114–125.Chen, A., Wang, A., Wang, T., Tang, X., & Zhang, M.(2017).Behavioral oscillations in visual attention modulated by task difficulty.Frontiers in Psychology, 8
, 1630.Chen, A.R., Tang, X.Y., Wang A.J., & Zhang, M.(2017).Experimental paradigms for discrete attention in visual domain.Advances in Psychological Science, 25
(6), 923–932.[陈艾睿, 唐晓雨, 王爱君, 张明.(2017).视觉注意离散性的实验范式.心理科学进展, 25
(6), 923–932.]Cohen, M.X.(2017).Where does EEG come from and what does it mean?Trends in Neurosciences, 40
(4), 208–218.Crick, F., & Koch, C.(1995).Are we aware of neural activity in primary visual cortex?Nature, 375
(6527), 121–123.Drewes, J., Zhu, W., Wutz, A., & Melcher, D.(2015).Dense sampling reveals behavioral oscillations in rapid visual categorization.Scientific Reports, 5
, 16290.Dugué, L., Marque, P., & VanRullen, R.(2015).Theta oscillations modulate attentional search performance periodically.Journal of Cognitive Neuroscience, 27
(5), 945–958.Dugué, L., McLelland, D., Lajous, M., & VanRullen, R.(2015).Attention searches nonuniformly in space and in time.Proceedings of the National Academy of Sciences of the United States of America, 112
(49), 15214–15219.Dugué, L., Merriam, E.P., Heeger, D.J., & Carrasco, M.(2017).Specific visual subregions of TPJ mediate reorienting of spatial attention.Cerebral Cortex
, 1–16, doi: 10.1093/cercor/bhx140.Dugué, L., Roberts, M., & Carrasco, M.(2016).Attention reorients periodically.Current Biology, 26
(12), 1595–1601.Dugué, L., & VanRullen, R.(2014).The dynamics of attentional sampling during visual search revealed by fourier analysis of periodic noise interference.Journal of Vision, 14
(2), 11.Dugué, L., & VanRullen, R.(2017).Transcranial magnetic stimulation reveals intrinsic perceptual and attentional rhythms.Frontiers in Neuroscience, 11
, 154.Dugué, L., Xue, A.M., & Carrasco, M.(2017).Distinct perceptual rhythms for feature and conjunction searches.Journal of Vision, 17
(3), 22.Fiebelkorn, I.C., Saalmann, Y.B., & Kastner, S.(2013).Rhythmic sampling within and between objects despite sustained attention at a cued location.Current Biology, 23
(24), 2553–2558.Gilinsky, A.S., & Doherty, R.S.(1969).Interocular transfer of orientational effects.Science, 164
(3878), 454–455.He, S., Cavanagh, P., & Intriligator, J.(1996).Attentional resolution and the locus of visual awareness.Nature,383
(6598), 334–337.He, S., & MacLeod, D.I.A.(2001).Orientation-selective adaptation and tilt after-effect from invisible patterns.Nature, 411
(6836), 473–476.Huang, Y., Chen, L., & Luo, H.(2015).Behavioral oscillation in priming: Competing perceptual predictions conveyed in alternating theta-band rhythms.Journal of Neuroscience,35
(6), 2830–2837.Hubel, D.H., & Livingstone, M.S.(1987).Segregation of form, color, and stereopsis in primate area 18.Journal of Neuroscience, 7
(11), 3378–3415.Hubel, D.H., & Wiesel, T.N.(1977).Ferrier lecture:Functional architecture of macaque monkey visual cortex.Proceedings of the Royal Society B: Biological Sciences,198
(1130), 1–59.Jia, J., Liu, L., Fang, F., & Luo, H.(2017).Sequential sampling of visual objects during sustained attention.PLoS biology
,15
(6), e2001903.Landau, A.N., & Fries, P.(2012).Attention samples stimuli rhythmically.Current Biology, 22
(11), 1000–1004.Landau, A.N., Schreyer, M.H., van Pelt, S., & Fries, P.(2015).Distributed attention is implemented through theta-rhythmic gamma modulation.Current Biology, 25
(17), 2332–2337.Macdonald, J.S.P., Cavanagh, P., & VanRullen, R.(2014).Attentional sampling of multiple wagon wheels.Attention,Perception, & Psychophysics, 76
(1), 64–72.McCollough, C.(1965).Color adaptation of edge-detectors in the human visual system.Science, 149
(3688), 1115–1116.Paradiso, M.A., Shimojo, S., & Nakayama, K.(1989).Subjective contours, tilt aftereffects, and visual cortical organization.Vision Research, 29
(9), 1205–1213.Pelli, D.G.(1997).The VideoToolbox software for visual psychophysics: Transforming numbers into movies.Spatial Vision, 10
(4), 437–442.Pooresmaeili, A., Poort, J., & Roelfsema, P.R.(2014).Simultaneous selection by object-based attention in visual and frontal cortex.Proceedings of the National Academy of Sciences of the United States of America, 111
(17), 6467–6472.Pooresmaeili, A., Poort, J., Thiele, A., & Roelfsema, P.R.(2010).Separable codes for attention and luminance contrast in the primary visual cortex.Journal of Neuroscience,30
(38), 12701–12711.Qian C.C., & Liu Z.X.(2016).High speed LED backlight tachistoscope reveals fast processing of topological invariants.Progress in Biochemistry and Biophysics, 43
(2), 157–166.[钱晨灿, 刘祖祥.(2016).高速LED背光速视器揭示拓扑性质的快速加工.生物化学与生物物理进展, 43
(2),157–166.]Schoups, A.A., & Orban, G.A.(1996).Interocular transfer in perceptual learning of a pop-out discrimination task.Proceedings of the National Academy of Sciences of the United States of America, 93
(14), 7358–7362.Shou T.D.(2010).Brain mechanisms of visual information processing
(2nd ed.).Hefei: China University of Science and Technology Press.[寿天德.(2010).视觉信息处理的脑机制
(第 2版).合肥:中国科学技术大学出版社.]Song, K., Meng, M., Chen, L., Zhou, K., & Luo, H.(2014).Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band.Journal of Neuroscience, 34
(14),4837–4844.Thiele, E., Pooresmaeili, A., Delicato, L.S., Herrero, J.L., &Roelfsema, P.R.(2009).Additive effects of attention and stimulus contrast in primary visual cortex.Cerebral Cortex,19
(12), 2970–2981.VanRullen, R.(2013).Visual attention: A rhythmic process?Current Biology, 23
(24), R1110–R1112.VanRullen, R.(2016a).Perceptual cycles.Trends in Cognitive Sciences, 20
(10), 723–735.VanRullen, R.(2016b).Perceptual rhythms.In J.Serences(Ed.),Stevens handbook of experimental psychology
.New York: Wiley.VanRullen, R., Carlson, T., & Cavanagh, P.(2007).The blinking spotlight of attention.Proceedings of the National Academy of Sciences of the United States of America,104
(49), 19204–19209.VanRullen, R., & Koch, C.(2003).Is perception discrete or continuous ?Trends in Cognitive Sciences, 7
(5), 207–213.VanRullen, R., Zoefel, B., & Ilhan, B.(2014).On the cyclic nature of perception in vision versus audition.Philosophical transactions of the Royal Society B: Biological sciences,369
(1641), 20130214.Wang, F., Chen, M., Yan, Y., Zhaoping, L., & Li, W.(2015).Modulation of neuronal responses by exogenous attention in macaque primary visual cortex.Journal of Neuroscience,35
(39), 13419–13429.Watson, A.B., & Pelli, D.G.(1983).Quest: A bayesian adaptive psychometric method.Perception & Psychophysics,33
(2), 113–120.Wolfe, J.M., & Franzel, S.L.(1988).Binocularity and visual search.Attention, Perception, & Psychophysics, 44
(1), 81–93.Zhang, X., Zhaoping, L., Zhou, T., & Fang, F.(2012).Neural activities in V1 create a bottom-up saliency map.Neuron,73
(1), 183–192.Zhang X., Yuan P.J., Wang Y., & Jiang Y.(2016).Neural entrainment and perception.Progress in Biochemistry and Biophysics, 43
(4), 308–315.[张雪, 袁佩君, 王莹, 蒋毅.(2016).知觉相关的神经振荡-外界节律同步化现象.生物化学与生物物理进展, 43
(4),308–315.]Zhaoping, L.(2008).Attention capture by eye of origin singletons even without awareness--A hallmark of a bottom-up saliency map in the primary visual cortex.Journal of Vision,8
(5), 1.Zoefel, B., & VanRullen, R.(2017).Oscillatory mechanisms of stimulus processing and selection in the visual and auditory systems: State-of-the-art, speculations and suggestions.Frontiers in Neuroscience, 11
, 296.