林生风毛菊(Saussurea sylvatica)头状花序权衡机制的海拔差异
2018-01-27王一峰祁如林杨亚军毛婉嫕
王一峰,祁如林,杨亚军,杨 洋,毛婉嫕
(西北师范大学 生命科学学院,甘肃 兰州 730070)
由于植物在不同季节的不同生命过程中对有限的资源存在定量的限值性,比如其生长发育、维持代谢、储藏能量、繁殖、修复、抵抗等各种生命活动都会需要资源且会发生竞争,导致植物产生如何最优化的分配这些资源的问题.因此,植物能不能准确合理的权衡这些生活史过程中的资源分配就显得尤为重要[1],只有处理好资源分配才能更好地适应复杂多变的环境.植物在其整个生活史过程中存在两个重大的权衡:一个是繁殖和其他生命活动间的权衡,主要表现为植物对自身繁殖的代价;另一个是后代花和种子的大小与数量间的权衡[2],表现为后代繁殖的适合度.但是,当前对这两种权衡关系的研究偏向营养投资和繁殖投资、种子的大小和数量、雌性分配和雄性分配、叶大小和数量、根系质量和茎叶质量等各种生物量之间[3-8].作为植物繁殖器官-头状花序,其结构大小与数目对植物的繁育系统和交配对策两者的进一步进化有不可估量的研究价值.因此,国内外很多研究植物生态和植物进化方向的专家对此非常青睐[9-11].植物的花部结构(花大小-数量)对繁殖分配甚至对传粉昆虫的吸引都有明显的影响,花数目可以表示其繁殖适合度,增大繁殖适合度就必须使花数量所占比重增大,导致产生种子的可能性增大[12].很多植物在高海拔区因为昆虫数量不足而造成访花频率和传粉效率的降低时,都选择延长花寿命或增加其传粉者吸引结构(花展示)的构建来增大花粉的传播[13-14].但是,植物将有限的资源同时分配到花数量和大小是极其困难的,妥协的结果就是在两者之间做出利益权衡[4]使这两个性状间出现负相关关系.迄今为止,对植物花大小-数量权衡关系的研究主要集中在龙胆科、蓼科和瑞香科狼毒属以及霸王花等植物[8,15,12,16,10,17],而对于风毛菊属植物花部生物学特征的变异以及对有限资源的权衡等问题在不同海拔梯度上的报道极为罕见[4].
青藏高原特殊的地理位置和气候条件被称作全球“第三极”以及“世界屋脊”,在这个极大的生态系统中蕴藏着许多种独特而复杂的植物类群.然而,生长在青藏高原的风毛菊属植物林生风毛菊是个常见种,其分布范围广,能够适应独特极端的高寒气候条件,对高寒草甸上的物种多样性有着一定的影响力.林生风毛菊(Saussureasylvatica)种群在青藏高原高海拔区承受的繁殖压力以及生存难度非常大,其在保证存活的前提下必须对自身繁殖略做出适应性的调整,重新对有限的资源进行合理分配和权衡,最后才能改变其整个种群的生态适应机制和权衡策略.随着海拔的升高,植株选择增加每个头状花序质量的投入,减少每株头状花序数目的投入来权衡资源分配,这种权衡现象目前只在少数研究中得到证实[18].本研究以青藏高原东北部的多年生植物林生风毛菊为对象,在了解其自然种群繁殖分配生物学特征的基础上,主要探讨林生风毛菊12个居群头状花序花大小与数量和海拔的相关性以及在高海拔区变异规律,深层次的研究花大小-数目之间权衡关系的差异性及其形成机制,旨在进一步了解林生风毛菊种群对高海拔环境的适应性对策,并为此类植物的开发利用提供理论依据.
1 样地、材料及研究方法
1.1 采样地概况
本试验材料均采集于青海湖西北部、柴达木盆地东缘的天峻县及其周边,全县面积为25 700 km2,地形主要以丘陵、湖泊、沼泽、平原、冰川为主,该区属于典型的高原大陆性气候(海拔高,气候寒冷,降水时间不均匀,年平均强风天数为97 d),其经纬度为96°49′~99°42′E,36°54′~39°12′N,境内海拔2 850~5 827 m,年降雨量400~800 mm.无绝对无霜期,日照时间充足.
1.2 研究材料
本材料为中国特有多年生植物林生风毛菊.紫色直立茎,有细沟纹,根状茎全部有纤维状撕裂的残迹叶;头状花序在茎枝顶端呈伞房花序状或圆锥花序状排列,总苞呈球形,外被白色卷毛,花苞直径2~5 cm;瘦果呈四棱形且浅褐色,长4~6 mm,顶端有两层小冠毛;叶呈披针形,长7~13 cm,宽0.8~2.0 cm,两面无毛,边缘有锯齿,基部下沿成3~5 cm的叶柄,顶端钝;生活环境常为高山灌丛、草地、云杉林缘、湿润石坡等.海拔2 700~4 800 m,花果期为7~9月[19].
1.3 取样方法
2015年8~10月,从低地到高山灌丛共选择6个海拔(3 400,3 700,3 900,4 050,4 200,4 400 m),不同海拔梯度选取两个居群(间隔约为100 m),每个居群随机采集20~25株完整样品植株(避免在剪取和运输过程中造成植株器官损失和丢失,还要压标本留凭证).首先测量每个植株高度(从茎基部到花顶的长度),再由地面剪取其地上部分(植株的根未必能够挖全),然后将样品分株编号带回实验室.在实验室先统计每株头状花序数量,再将植株的地上部分分成繁殖器官(包括所有的头状花序以及其各个苞片)和营养器官(包括茎以及花梗、叶以及叶柄)两部分,最后将各部分放置于80 ℃的烘箱24 h,再用天平分别称量每个头状花序的质量及营养器官和繁殖器官生物量.
1.4 数据处理
两个居群所有测量数据都取其平均值;个体大小用地上部分生物量和株高两部分表示;地上部分生物量用繁殖和营养器官生物量干重之和表示;繁殖器官生物量用每株头状花序总生物量干重表示;繁殖分配为繁殖器官生物量干重占地上部分生物量干重的百分比;花大小用单个头状花序重量表示,即植株每株头状花序总生物量与头状花序花总数量的比值;花数量指植株上的头状花序总数量(已经开放过、正在开放的花以及未开放的花).
实验数据均采用Excel 2003进行统计,并用Origin 8.5绘图,最后用SPSS 20.0对林生风毛菊各种生物量与海拔及花大小和数量间的权衡关系进行线性回归和方差分析[20](显著性水平设为0.05).
2 结果与分析
2.1 各生物学特征及繁殖分配与海拔的相关性
不同海拔梯度林生风毛菊株高和地上生物量均存在极显著的差异(图1和图2,P<0.01),随着样地海拔高度的增加,地上生物量和株高均呈逐渐减小趋势,即林生风毛菊植株个体大小与海拔呈负相关关系.
在种群水平上,不同海拔林生风毛菊的繁殖和营养器官生物量均与海拔呈极显著负相关关系(图3,P<0.01),即两个器官生物量都随着海拔的升高而减小,但繁殖器官生物量减小的幅度比营养器官减小的幅度稍小.
不同海拔繁殖分配存在显著差异(图4,P=0.023),即林生风毛菊的繁殖分配随海拔的升高呈逐渐增大趋势,繁殖分配与海拔呈显著正相关系.
2.2 不同海拔头状花序大小与数量的变化
不同海拔梯度下林生风毛菊的每株头状花序数目存在极显著差异(图5,P<0.01),与海拔呈极显著负相关;而每个头状花序重存在显著差异(P=0.02),与海拔呈显著正相关.所以,随着海拔的升高,林生风毛菊头状花序总数量不断减少,而每个头状花序质量不断增加.
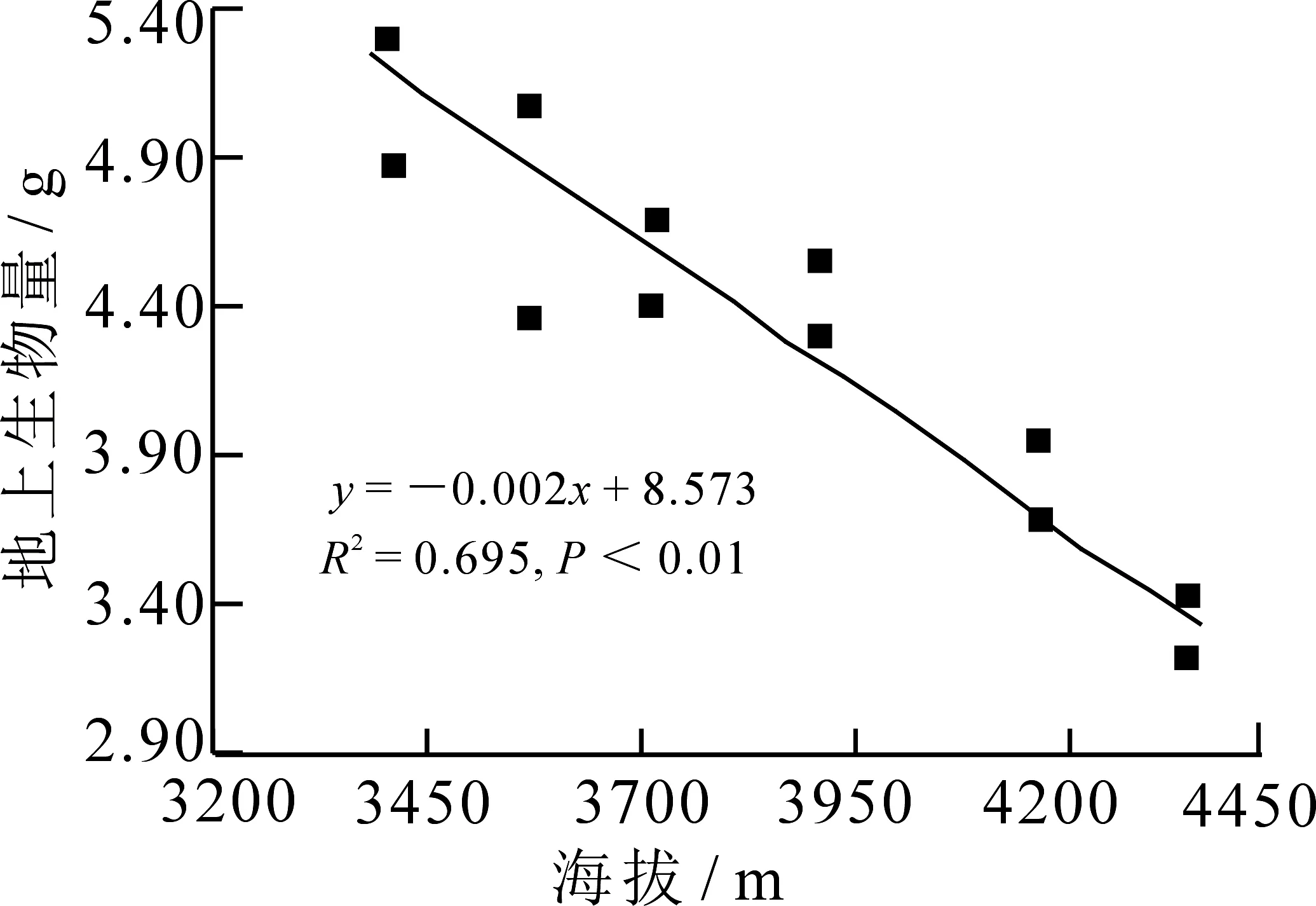
图1 地上生物量与海拔的线性回归关系
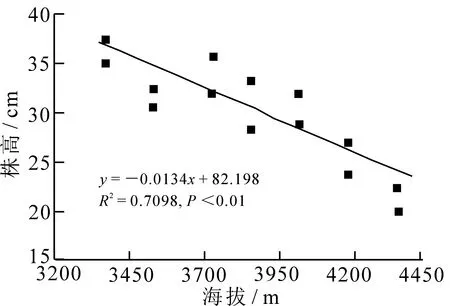
图2 株高与海拔的线性回归关系
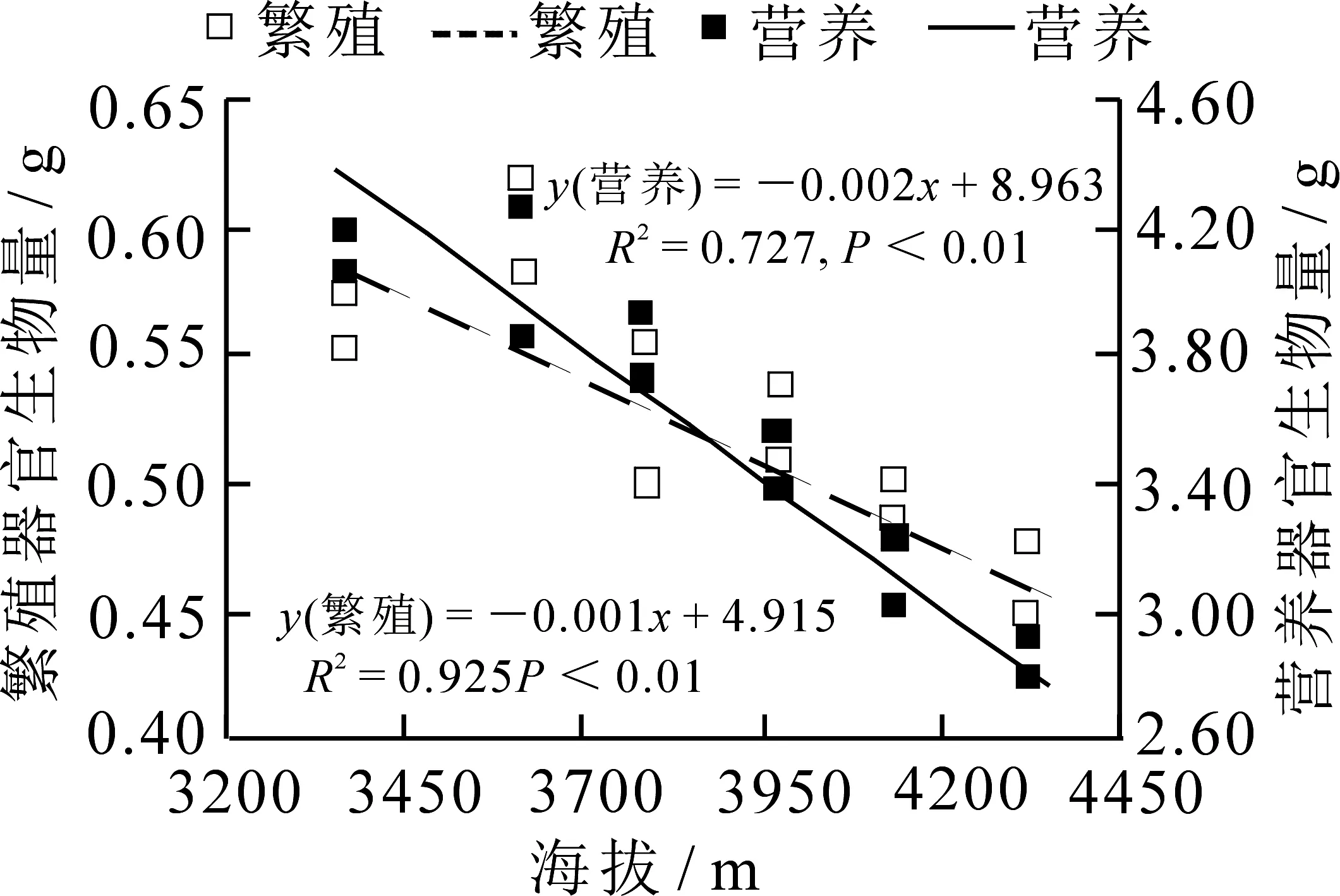
图3 繁殖器官生物量及营养器官生物量与海拔的线性回归关系
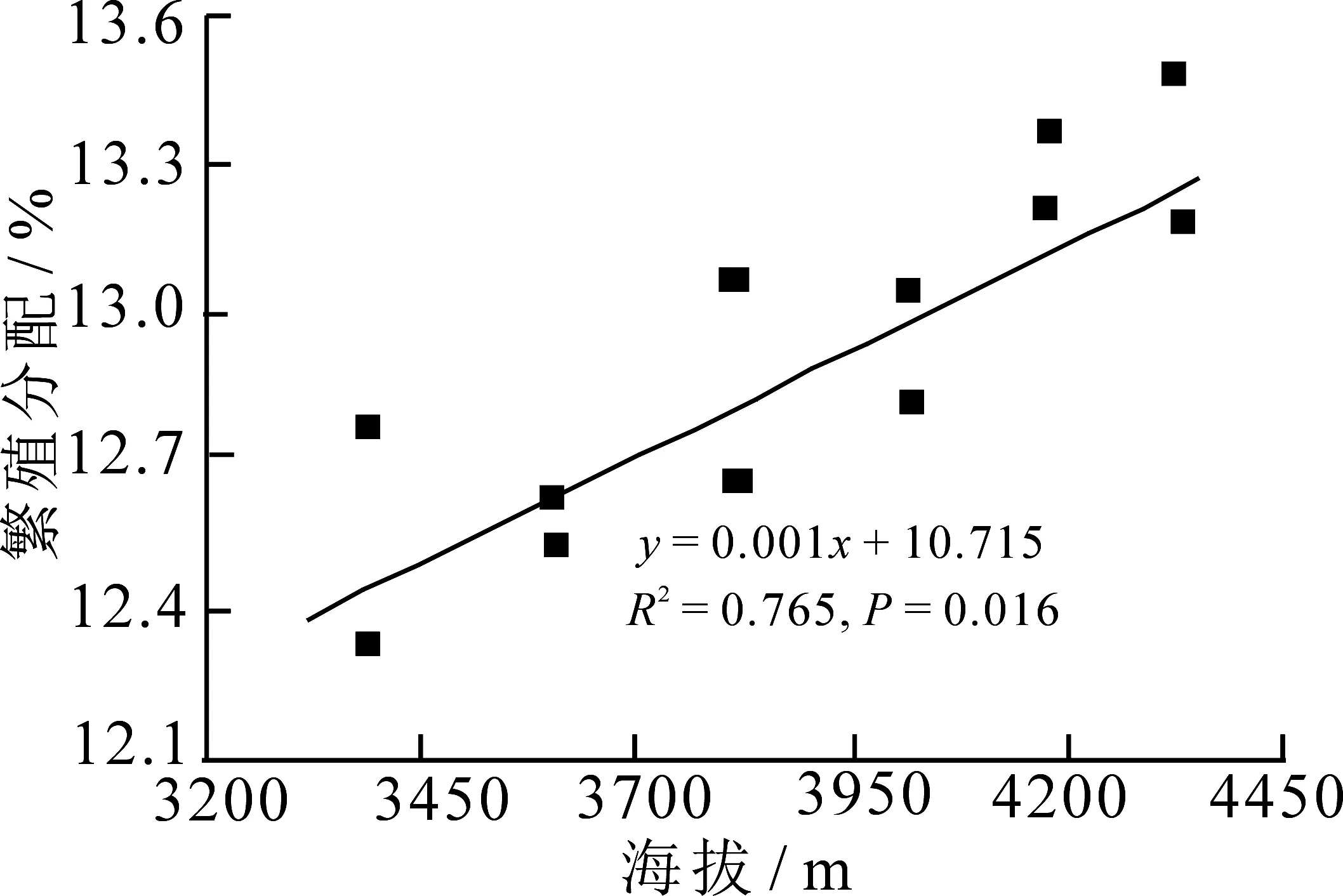
图4 繁殖分配与海拔的线性回归关系
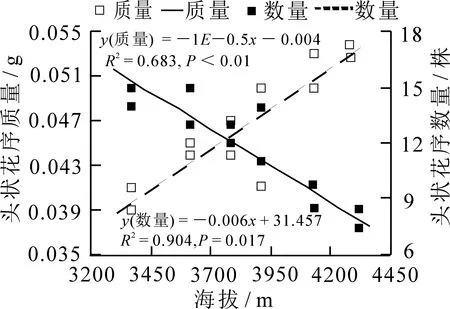
图5 每株头状花序数量及每个头状花序质量与海拔的线性回归
2.3 头状花序大小与数量之间的权衡关系
由图6显示,林生风毛菊每个头状花序质量与每株头状花序数量之间呈负相关,即随着每个头状花序质量的增加,每株头状花序总数量呈现不断减小趋势,说明花的大小和数量之间存在权衡.
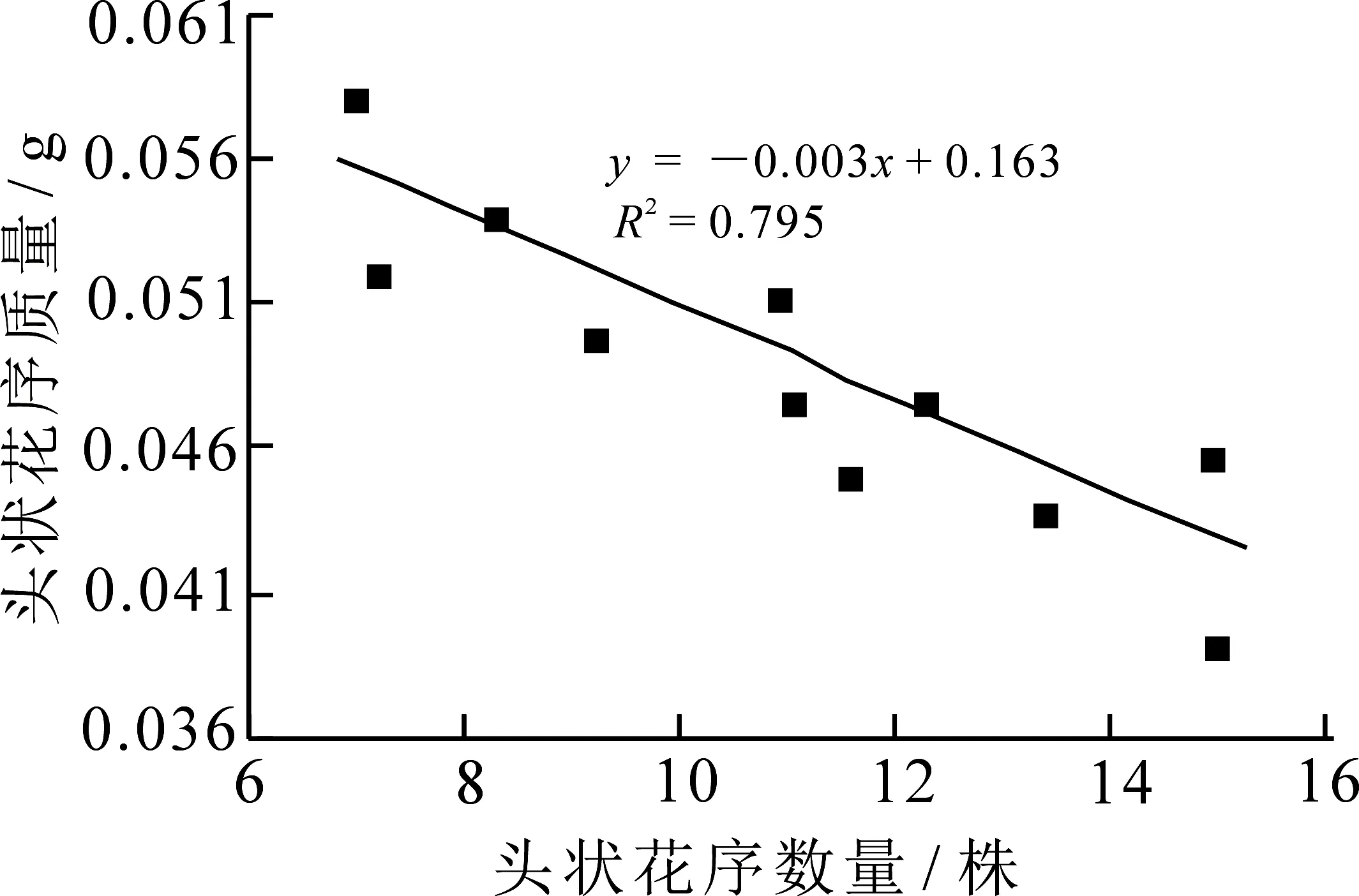
图6 每株头状花序数量与每个头状花序质量的线性回归
3 讨论
在植物个体的生长发育以及繁殖等生命活动中,高海拔区植物必须根据不同器官的不同功能把有限的资源进行最合理最优化的分配,才能维持其整个生活史的生命过程[21].高海拔区温度的不断降低和生长季节的缩短等环境胁迫导致有效的资源量减少[22],使最大光合能量的获取逐渐降低,最后在可获取的有限资源总量范围内分配给不同结构和器官组织的资源量也逐渐降低[23].对6个不同海拔居群虎耳草科岩白菜的地上部分总生物量、花数目以及营养和繁殖器官等生物量的研究得出,这些生物学特征都与海拔的升高而明显的呈现降低趋势[24-25].本研究发现,林生风毛菊在恶劣环境胁迫下也可做出适应性调整,主要体现在个体大小(地上生物量和株高)、繁殖和营养器官生物量均与海拔呈现显著负相关关系(图1-图3),均符合上述研究理论.分析以上这种负相关关系产生的原因可能是:① 为了应对青藏高原高海拔地区恶劣气候环境(低温强辐射、大风力、冰雹冰雪等)做出的繁殖对策,包括将更多的资源投资给繁殖器官,反而减少对营养器官的投入,植物形成小个体来节约储藏资源,将有限的资源量分配给花部结构.② 根据高海拔区植物资源受限理论,有限的资源量只能满足较小植株个体器官的营养和水分传输需求[27];另外,小的植株个体可以减少其在高原极端环境中繁殖器官的暴露,从而减少牛羊等食草动物的践踏和啃食以及低温和紫外线等恶劣条件的伤害[19].③ 林生风毛菊植株多生长于灌木丛中,由于相邻的小灌木植株较多,不排除植物之间强烈的太阳光竞争使其同化的生物量减小,还有低温对植物合成新的细胞壁过程受限使其生长发育受阻[26].除了林生风毛菊本身固有的可塑性和遗传性外,影响植物个体大小的最主要的因素是各种外界环境因子(生物和非生物),所以青藏高原独特的环境条件严重影响了其生活史且增加了植物的繁殖困难度.如牛羊等食草动物的践踏和啃食、强风冰雹的损伤以及生长季节缩短和频繁的低温(霜冻),使植株只能利用土壤解冻后有限的时间和有限的资源进行生长和繁殖,而且较矮的株高所形成的温暖花部环境由于可以吸引更多的传粉者访花.因此林生风毛菊随着海拔的升高以减小个体大小以及各种器官生物量的方式来应对环境胁迫的压力,也就是减小个体大小来削弱营养器官以达到促进生殖生长的目的.
影响和调节植物各种生命特征的许多环境因素(如温度、生长季长度、资源有效性)都会随着海拔不断的升高而逐渐减小,使得不同海拔居群间的繁殖策略等都会发生适应性的改变[13].本研究发现,林生风毛菊的繁殖分配随海拔的升高而呈增加趋势(图4),与最优分配模型的理论相反[24,21],但与张林静等[25]的研究结论完全一致;也有对比性研究生长在低海拔区(600 m)的20个物种以及生长在高海拔(2 700 m)的30个物种间的关系,最后发现高海拔区植物的繁殖分配与低海拔地区的植物相比高出许多,得出有性繁殖对高海拔区植物具有更为重要的作用[28].还有研究对钝裂银莲花[29]、狼毒[6]、川西风毛菊[7]等植物不同居群繁殖分配随海拔的相关性变化结果也都支持两者呈正比关系.引起林生风毛菊这种繁殖策略的原因可能是:① 本材料为虫媒传粉植物,高海拔地区的低温、薄弱的含氧量和短暂的生长季等气候条件加剧了传粉昆虫在传播花粉过程中的困难,再加上突发性暴风雨对花粉的冲刷作用,对昆虫的访花频率和传粉效率产生极其严重的影响,导致花粉数量的大大减小.② 海拔越高生态环境越恶劣,为确保林生风毛菊种群生活史的完整和后代的繁衍问题,其选择将更多的资源投入到繁殖结构(头状花序)以增大其繁殖分配[6].③ 许多草本植物的繁殖分配不仅与各种生物因素(水、温、气、盐、矿)有关,而且还受各种非生物因素(啃食者、竞争者、传粉媒介)的影响.因此,得出繁殖分配随着海拔升高而呈正相关关系结论[25,23],植物提高繁殖分配是为了将更多资源投入到花部结构,产生更大“少而优”的花使花展示更加完美鲜艳,而且昆虫的访花频率是植物获得繁殖成功的关键性因素,所以必须增加对花部特征(花展示)和昆虫吸引结构的构建以达到“广告投资”的目的,才能提高花粉的传播效率以及潜在的交配多样性[30],保证其有性生殖才体现出增大花大小,减小花数量的权衡关系.
高原环境中的植物在能够保证生存的前提下其繁殖比生长占有更重要的地位,权衡是研究植物生活史过程中的核心问题,资源条件的有限使许多生命特征(如生长、繁殖和存活)之间存在着各种利益的争夺和权衡[31].植物的花大小与数量两者间的权衡机制对植物有性繁殖的成功起着决定性作用,其也是对外界恶劣环境做出的适应性变化的一种表现[32].本研究得出林生风毛菊头状花序的数量和质量随着海拔的不断升高存在着权衡关系,其增加每个头状花序的质量大小,而减少对每株植物的头状花序的总数目(图5-6).以下研究都能支持本研究结果:汪洋[4]、陈婷[33]和张茜[16]等分别对风毛菊、大耳叶风毛菊和狼毒等植物种群花大小与数量的研究发现,不管低海拔种群还是高海拔种群都表现出明显的负相关权衡关系,但是越恶劣的胁迫环境条件越是加剧两者之间的权衡进程,高海拔种群比低海拔种群产生的头状花序和管状小花更大更重,但是两者平均数量呈现降低趋势.林生风毛菊做出这种权衡的主要原因可能有3种:① 在高海拔区的生长繁殖过程中为了提高繁殖分配和传粉的成功率,选择把更多的资源投入到花部特征(花展示)的构建[34],包括花柱、花柱分支和花丝等的伸长.② 如果同一个体同时开放较多的花会引起同株异花授粉和花粉折损[35],所以选择产生大而少的花来延长花寿命并且加大花粉的输入和输出,提高传粉几率.③ 高海拔地区的植物会分配更多的生物量给花部结构而不是茎叶,这种分配有利于植物增加花展示和报酬[28],为果期产生少而优的种子以及种子的萌发做准备.本研究发现,在青藏高原的高海拔环境中,不同海拔林生风毛菊头状花序的数量和质量之间存在权衡关系,植株这种选择产生少而优的头状花序的资源分配模式可能是其提高繁殖成功率的一种生存策略,说明植物在不断增加对花内部性器官和花展示的保护和辅助结构资源量的投入,以此来确保繁殖部分结构的完整和繁殖器官及性器官的成熟,以及吸引更多的昆虫来提高访花率,以此达到有性生殖和繁殖成功的目的.林生风毛菊分配到花大小与数目的资源间存在着“此消彼长”的权衡,而且花大小-数量之间的权衡关系具有环境依赖性,使其能够对外界各种环境的变化做出响应,这种繁殖对策在植物生活史上能提高其适合度,并为植物权衡理论提供了新的证据.
[1] REGON M,HARPER J L,TOWNSEND C R.Ecology:Individuals,PopulationsandCommunities[M].London:Black Well,1990.
[2] SILVERTOWN J W,LOVETT D J.Introduction to plant population biology[J].BlackwellScientificPublications,1993,79(23):14l.
[3] 王一峰,岳永成.青藏高原东缘不同海拔波缘风毛菊资源分配及花部特征对种子数目和质量的影响[J].植物生态学报,2014,38(4):366.
[4] 汪洋,杜国祯,郭淑青,等.风毛菊花序、种子大小和数量之间的权衡:资源条件的影响[J].植物生态学报,2009,33(4):681.
[5] 赵志刚.青藏高原高寒草甸常见毛茛科植物繁殖对策研究[D].兰州:兰州大学,2006.
[6] 张茜,赵成章,马小丽,等.高寒草地狼毒种群繁殖分配对海拔的响应[J].生态学杂志,2013,32(2):247.
[7] 王一峰,靳洁,曹家豪,等.川西风毛菊花期资源分配随海拔的变化[J].植物生态学报,2015,39(9):901.
[8] WORLEY A C,BAKER A M,THOMPHON J D,et al.Floral display in narcissus:variation in flower size and number at the species,population,and individual levels[J].InternationalJounalofPlantScience,2000,161(18):69.
[9] ZHAO Z G,WANG Y K.Selection by pollinators on floral traits in generalizedTrolliusranunculoides(Ranunculaceae)along altitudinal gradients[J].PLOSONE,2015,10(6):118.
[10] CARUSO C M .The quantitative genetics of floral trait variation in Lobelia:potential constraints on adaptive evolution[J].Evolution,2004,58(17):732.
[11] 路宁娜,曹素珍,王一峰.川西风毛菊(Saussureadzeurensis)花展示的变异:检验花及花序水平大小-数量的权衡[J].生态学杂志,2016,35(4):843.
[12] 陈学林,梁艳,齐威,等.一年生龙胆属植物的繁殖分配及其花大小、数量的衡关系研究[J].草业学报,2009,18(5):58.
[13] BINGHAM R A,ORTHNER A R.Efficient pollination of alpine plants[J].Science,1998,391(6):238.
[14] KUDO G,M0LAU U.Variations in reproductive traits at inflorescence and flower levels of an arctic legume,AstragalusalpinusL:comparisons between a subalpine and an alpine population[J].PlantSpeciesBiology,1999,14(3):181.
[15] 操国兴,李燕,刘欣.水蓼花大小在花序和个体上的变化及其与数量权衡关系研究[J].广西植物,2008,28(3):307.
[16] 张 茜,赵成章,董小刚,等.高寒退化草地狼毒种群不同海拔花大小-数量的权衡关系[J].植物生态学报,2014,38(5):452.
[17] SARGENT R D,GOODWILLIE C,KALISZ S,et al.Phylogenetic evidence for a flower size and number trade-off[J].AmericanJournalofBotany,2007,94(7):2059.
[18] TOMIMATSU H,OHARA M.Evolution of hierarchical floral resource allocation associated with mating system in an animal-pollinated hermaphroditic herb,trillium cam-schatcense(Trilliaceae)[J].AmericanJournalofBotany,2006,93(1):134.
[19] 石铸,靳淑英.中国植物志[M].北京:科学出版社,1999.
[20] GUO H,MAZER S J,DU G Z.Geographic variation in primary sex allocation per flower within and among 12 species ofPedicularis(Orobanchaceae):proportional male investment increases with elevation[J].AmericanJournalofBotany,2010,97(8):1334.
[21] 梁艳.青藏高原东缘龙胆属植物繁殖分配对策研究[D].兰州:西北师范大学,2009.
[22] 苏梅,齐威,阳敏,等.青藏高原东部大通翠雀花的花特征和繁殖分配的海拔差异[J].兰州大学学报(自然科学版),2009,45(2):61.
[23] MENDEZ M,TRAVESET A.Sexual allocation in single-flowered hermaphroditic individuals in relation to plant and flower size[J].Oecologia,2003,137(1):69.
[24] 王赟,胡莉娟,段元文,等.岩白菜(虎耳草科)不同海拔居群的繁殖分配[J].云南植物研究,2010,32(3):270.
[25] 张林静,石云霞,潘晓玲.草本植物繁殖分配与海拔高度的相关分析[J].西北大学学报(自然科学版),2007,37(1):77.
[26] KORNER C H.FunctionalPlantEcologyofHighMountainEcosystems[M].Beijing:Science Press,2008.
[27] MCCULLOH K A,SPERRY J S.Patterns in hydraulic architecture and their implications for transport efficiency[J].TreePhysiology,2005,25(5):257.
[28] FABBRO T,KORNER C.Altitudinal differences in flower traits and reproductive allocation[J].FunctionalEcologyofPlants,2004,199(1):70.
[29] 李冰,刘左军,赵志刚,等.海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响[J].草业学报,2013,22(1):10.
[30] HARDER L D,BARRETT S C H.PollenDispersalandMatingPatternsinAnim-pollinatedPlants[M].New York:Chapman &Hall,1996.
[31] STEARNS S C.TheEvolutionofLifeHistories[M].New York:New York University Press,1992.
[32] 王永健,方兴,钟章成.不同生境对蝴蝶花花部与果实特征的影响[J].生态学报,2010,30(17):4628.
[33] 陈亭,王一峰.青藏高原东缘大耳叶风毛菊繁殖特征对海拔的响应[J].生态学杂志,2014,33(12):3216.
[34] 王一峰,刘启茜,裴泽宇,等.青藏高原3种风毛菊属植物的繁殖分配与海拔高度的相关性[J].植物生态学报,2012,36(1):39.
[35] HARDER L D,BARRETT S C H.Mating cost of large floral displays in hermaphrodite plants[J].Nature,1995,373(14):512.
