DRD4基因rs752306多态性与蒙古族学龄儿童攻击行为的相关性研究
2018-01-24王燕
王燕
摘要:为了解DRD4基因rs752306多态性与蒙古族学龄儿童攻击行为的相关性,选取内蒙古赤峰市天山镇蒙古族实验小学302名7~12岁蒙古族儿童为调查对象,运用Achenbach儿童行为量表(CBCL)评价了儿童攻击行为得分;采集所有儿童口腔上皮细胞并提取DNA,采用iMLDR法测定了位点基因型;利用广义线性模型(GLM)檢验了主效应及交互作用大小。研究结果发现:DRD4基因rs752306住点有三种基因型GG、GA、AA,携带A等位基因的蒙古族学龄儿童攻击行为得分高于其他两种基因型;蒙古族学龄儿童年龄和父亲职业对蒙古族儿童攻击行为有影响,差异具有统计学意义(P<0.05);蒙古族儿童年龄和rs752306多态性的主效应及两者的交互作用对蒙古族儿童攻击行为有显著影响(P<0.05)。因此,DRD4基因rs752306多态性与蒙古族儿童攻击行为问题可能存在一定相关性。
关键词:蒙古族;学龄儿童;攻击行为;基因多态性
中图分类号:R72
文献标识码:A
文章编号:1674-9944(2018)01-0220-04
1 引言
攻击行为可以用多种方式评估,研究采用苏林雁等标准化的常模评价儿童行为问题得分高低。攻击行为是儿童青少年最常见的一种社会行为,其发生率较高。学龄期儿童的攻击行为会持续影响其身心发展,导致青少年期的行为问题及成年期精神障碍甚至犯罪等[1]。双生子研究表明,儿童攻击行为的遗传度为中等偏高水平51%~72%[2,3]。
近年来,相关研究发现,多巴胺D4受体(D4 dopamine receptor,DRD4)基因多态性与不同行为存在相关性,包括暴力攻击行为[4]、帕金森病[5]、酒精中毒[6]、注意缺陷多动障碍[7]、精神分裂症[8]、双相情感障碍[9]、猎奇的人格特征[10]等。目前共发现DRD4基因有21种多态性,非编码区存在至少18种多态性,其中rs752306( MAF—0.18)位于DRD4基因第1内含子中,为非编码标记位点,但其位置与编码区非常接近。rs752306多态性干扰转录因子与至少5种以上位点的结合。Zhang等11]研究发现,DRD4基因rs752306多态性与儿童智力发育迟缓存在弱相关性。Docherty等[12]对基因间协同作用的机制进行了探究,发现rs752306多态性与DNA甲基化相关。目前,国内外对儿童攻击性行为的同类研究较少,内蒙古自治区是一个以蒙古族为主体,多民族聚居的少数民族地区,拥有多元化的文化特点。不同民族存在遗传背景、文化等差异,对个体的行为发展可能会造成一定影响。因此,本研究以蒙古族学龄儿童为研究对象,报道DRD4基因rs752306多态性与攻击行为的相关性,对于加强相关研究深入了解蒙古族学龄儿童心理行为问题的影响因素将有重要意义。
2 材料与方法
2.1 研究对象
以学校为单位,随机整群抽取赤峰市天山镇蒙古族实验小学7~12岁蒙古族学龄儿童进行问卷调查及口腔上皮组织采集,有效样本量302名,儿童平均年龄为9. 22±1.52岁。按照统一标准筛选,被试儿童直系三代均为蒙古族,且所有个体均无心理疾病和明显行为异常,家中无精神疾病史。本研究均在取得学校、家长的知情同意基础上完成。
2.2 研究方法
2.2.1 问卷调查
由家长填写Achenbach儿童行为量表(ChiLd Be-havior Checklist,CBCL) [13]并计算攻击行为得分,得分越高,表明攻击行为水平越高。该量表具有较高的信度和效度。同时,家长填写家庭一般情况调查问卷,问卷中包括儿童性别、年龄、父母职业及文化程度等一系列家庭一般状况。
2.2.2 全基因组DNA提取
儿童用清水漱口后,采用一次性口腔拭子刮取口腔上皮细胞,置于含样本保存液的离心管中保存。天根试剂盒提取全基因组DNA,利用紫外分光光度计、琼脂糖凝胶电泳检测其浓度和纯度。
2.2.3
DRD4基因rs752306多态性检测
检索与DRD4基因相关的单核苷酸多态性(SingleNucleotide Polymorphisms,SNP)位点,采用的iMLDR(improved multiple ligase detection reaction)法测定位点基因型,上游引物:5- AGACGCCCACCAACTC-CTTCAT -3;下游引物:5- GTAGGGACAGGGACGGGGAAC -3。聚合酶链式反应( polymerasechain reaction,PCR)体系:lx GC -I buffer,3.o mMMg2+,0.3 mM dNTP,l U HotStarTaq polymerase,lμL样本DNA和上下游各1μL引物。PCR循环程序:95℃2 min预变性,94℃20 s变性,65℃40 s退火,72℃1.5 min延伸,72℃2 min后延伸,4℃forever。然后进行产物纯化:在20 μL PCR产物中加入5 U SAP酶和2 U Exonuclease I酶,37℃温浴th然后75℃灭活15 min。连接引物:FC; TTCCGCGTTCGGACT-GATATGGCYGCACGAGCATCCTGAC
FT: TACGGTTATTCGGGCTCCTGTGGCYGCACGAGCATCCTGAT
RP: CTGCTCCTCGGTTCCCCGTTTTTTTTT
连接反应体系:连接缓冲液lμL、高温连接酶0. 25μL、5连接引物混合液(μM)0.4μL,3连接引物混合液(2 μM)O.4 μL、纯化后多重PCR产物2μL、ddH206μL混匀。连接程序:94℃1 min变性,56℃4 min退火延伸,38次循环,4℃保存。取0.5μL稀释后的连接产物,上ABI3730XL毛细管电泳仪,收集数据分析。
2.3 统计学分析
使用SPSS 16.O统计软件进行数据处理与分析。运用KoLmogorow Smirnov检验对攻击行为得分进行正态性检验。单因素分析采用非参数检验,两个独立样本比较采用Kolmogorov - Smirnov Z检验(简称K-S检验),多个独立样本间比较采用Kruskal Wallis检验(简称K-W检验);多因素分析采用广义线性模型(GeneraLized Linear Models,GLM)检验主效应及交互作用大小。
3 结果
3.1 Hardy- Weinberg平衡的吻合度检验
对DRD4基因rs752306位点进行Hardy - Weinberg平衡吻合度检验。结果显示,该位点的基因型(GG、GA、AA)期望值与观测值吻合度较好(x2=2. 733,P>O05),符合Hardy- Weinberg平衡定律。
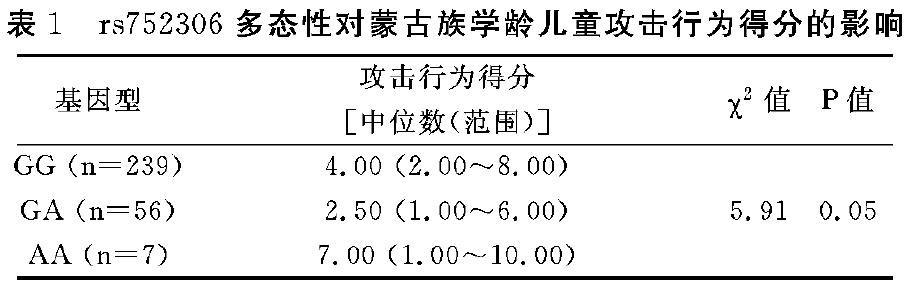
3.2 DRD4基因rs752306多态性对蒙古族学龄儿童攻击行为得分的影响
经非参数检验分析,DRD4基因rs752306多态性与蒙古族儿童攻击行为问题得分可能存在相关性( x2=5.908,P=O.05),P值接近统计检验临界值(见表1),携带A等位基因的蒙古族学龄儿童攻击行为得分高于其他两种基因型。
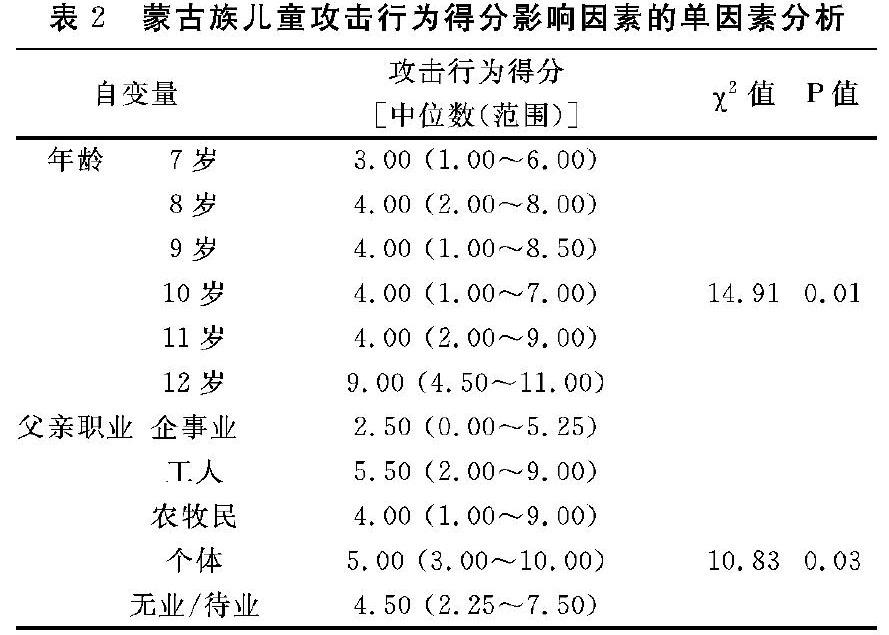
3.3 影响蒙古族学龄儿童攻击行为得分的单因素分析
以蒙古族学龄儿童攻击行为得分为因变量,以可能影响攻击行为得分的性别、年龄、父母职业、父母文化程度等因素为自变量,进行单因素分析。结果显示,年龄、父亲职业是蒙古族儿童攻击行为得分的影响因素(P<0.05),11~12岁蒙古族儿童攻击行为得分明显高于其他年龄段,父亲职业为企事业的蒙古族儿童攻击行为得分明显低于其他职业,见表2。因此,将年龄分为低年龄段(7~lO岁)和高年龄段(11~12岁)两组,父亲职业分为“企事业”和“其他”两组进行多因素分析。
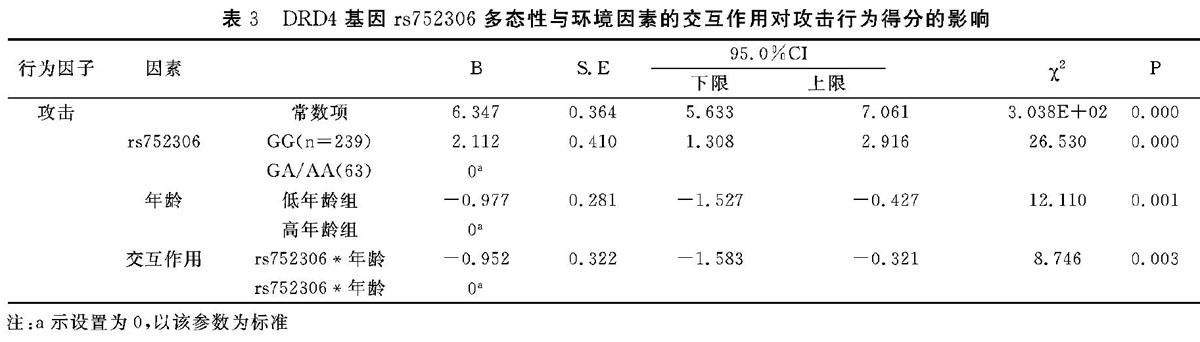
3.4 DRD4基因rs752306多态性与环境因素交互作用对蒙古族学龄儿童攻击行为的影响
将蒙古族儿童攻击行为得分作为因变量,年龄、父亲职业为自变量,进行广义线性模型(Generalized Line-ar Models,GLM)分析主效应及交互作用对攻击行为的影响。由于rs752306的AA基因型出现次数很少,因此,在进行主效应与交互作用时将AA基因型与GA基因型进行了合并。结果显示,儿童年龄和rs752306多态性的主效应及两者的交互作用对蒙古族儿童攻击行为有显著影响(P<0.05),见表3。
4 讨论
据WHO报告显示,全世界10%~20%的儿童青少年患有精神障碍,精神疾病是导致自残、自杀等的主要原因。由中国儿童中心(CNCC)最近公布的数据显示,在中国至少有3000万名17岁以下儿童青少年表现出一系列的行为问题[14]。近年来,随着城镇化进程加快,大多数来自牧区或农村的蒙古学龄儿童到城市求学,他们寄宿在学校、亲戚家或者由爷爷奶奶等陪读,父母没有更多时间照顾儿童的身心问题,导致父母与儿童关系越来越紧张,儿童出现的心理行为问题可能得不到及时指导和治疗。攻击行为是一种最常见的心理行为问题,严重会导致暴力、犯罪等行为的发生[15]。Ettekal等[1]研究发现,儿童早期的攻击行为会随年龄不断增加,持续到儿童后期且更具有敌意。与本研究结果相似,11~12岁儿童的攻击行为得分明显高于其他年龄段。因此,儿童心理行为问题应早预防、早发现及早治疗,对其健康成长非常重要。此外,父亲职业是企事业的蒙古族儿童攻击行为得分低于其他父亲职业的儿童(P
DRD4基因位于人类第11号染色体短臂15区5带(11P15.5),具有5个外显子,4个内含子。DRD4基因mRNA主要集中分布于大脑皮层的边缘系统,提示在认知与情绪方面具有重要作用[17]。目前,DRD4基因第3外显子48bp多态性及编码区SNP位点如rs1800955、rs936461是注意缺陷多动障碍、攻击行为、认知能力及人格特质等研究的热点[18~20],而位于非编码区SNP位点相关研究较为罕见,非编码区序列有与RNA聚合酶的结合位点,它的调控作用不容忽视。研究表明rs752306多态性具有干扰转录因子与位点结合的功能[12]。
本研究中rs752306多態性基因型出现频率(GG:79.14%,GA:18. 54%,AA:2.31%)与Lee等[21]对韩国人群研究结果基本一致。另外,经非参数检验发现,rs752306多态性与攻击行为可能存在相关性,携带A型等位基因的蒙古族学龄儿童攻击行为得分最高,更容易发生攻击行为。因此,对于携带有A型等位基因的儿童,教养时更应注意适度关注儿童的发展,给与其多方面的爱护,从而减少儿童心理行为问题的发生。本研究还发现,rs752306多态性与年龄的交互作用对攻击行为得分的影响具有统计学意义(P<0.05),提示携带A等位基因的高年龄段蒙古族儿童可能更容易发生攻击行为,G等位基因的保护作用只有在低年龄段时才能体现。因此,携带A等位基因蒙古族儿童攻击行为在低年龄段就应该需要得到较多关注。
以往的研究中忽略DRD4基因非编码区多态性的作用及功能,本研究以蒙古族学龄儿童为对象探讨了rs752306多态性与攻击行为的相关性问题,其研究结果可为相关研究提供一定线索。今后可考虑对更多蒙古族族群儿童心理行为问题进行研究,同时考虑分析多个基因对蒙古族儿童心理行为问题的协同作用,并结合蒙古族儿童家系、双生子及表观遗传学的角度开展更深入的研究。
参考文献:
[l]Ettekal I,Ladd G W. Developmental continuity and change inphysical, verbal, and relational aggression and peer victirruzationfrom childhood to adolescence[J]. Developmental Psychology,2017, 53(9) :1709~1721.
[2] Miles D R, Carey G.Genetic and environmental architecture ofhuman aggression[J]. Journal of PersonaLity and Social Psychology, 1997, 72(1):207~217.
[3]Hudziak J J,Beijsterveldt C E,Bartels M. Individual differencesin aggression: genetic analyses by age, gender, and informant in 3-, 7- , and lO-year-old[J]. Behavior Genetic, 2003, 33(5):575~589.
[4]封宇.重庆地区暴力罪犯的DRD4基因外显子ⅢVNT R多态性、心理和生活环境的关联性分析[D].大连:大连海事大学,2011.
[5]Eisenegger C,Knoch D, Ebstein R P,et al.Dopamine receptorD4 polymorphism predicts the effect of L-DOPA on gambling behavior[J]. Biological Psychiatry, 2009, 67(8):702~706.
[6]Du Y, Yang M, Yeh H W, et al.The association of exon 3 VNTR polymorphism of the dopamine receptor D4 (DRD4) gene withalcoholism in Mexican Americans[J]. Psychiatry Research, 2010,177(3):358~360.
[7]Faraone S V, Mick E.Molecular genetics of attention deficit hy-peractivity disorder[J]. The Pediatric Clinics of North America,2010, 33(1):159~180.
[8]Lai J H, Zhu Y S,HuoZ H, et al,Association study of polymorphisms in the promoter region of DRD4 with schizophrenia, de-pression, and heroin addiction[J]. Brain Research, 2010 (1359):227~232.
[9] Nemoda Z,Lyons-Ruth K, Szekely A, et al.Association7between dopanminergic polymorphisms and borderline personalitytraits among at-risk young adults and psychiatric inpatients[J].Behavior and Brain Functions, 2010, 6(4):1~11.
[lO]Tsuchimine S,Yasui-Furukori N, Kaneda A, et al.Minor ge-netic variants of the dopamine D4 receptor (DRD4) polymor-phism are associated with novelty seeking in healthy Japanesesubjects[J]. Progress ln Neuro-psychopharmacol and BiologicalPsychiatry, 2009, 33(7):1232~1235.
[ll]Zhang K, Zhang M J,He H, et al.A family-based associationstudy of dopamine receptor D4 and mental retardat/on in Qinbaregion of China[J]. Neuroscience Letters, 2012, 516(1):1~4.
[12]DochertyS J,Daviso SP,HaworthCMA, et al.A genetic association study of DNA methylation levels in the DRD4, gene re-gion finds associations with nearby SNPs[Jl. Behavioral& BrainFunction.s, 2012(8):1~13.
[13]汪向东,王希林,马弘.心理卫生评定量表手册[M].北京:中国心理卫生杂志社,1999:49~52.
[14]仝嵐.CBCL在吉林省儿童行为问题中的应用研究[D].长春:吉林大学,2009.
[15]马中锐,蒙华庆,胡华,等.反社会行为始于青少年期和儿童期的暴力犯罪青少年童年受虐史、攻击行为的差异比较[J].第三军医大学学报,2012, 34(24):2511~2514.
[16]曲红明,吕伶,李芹.等.南京市学龄前儿童行为问题及其家庭环境影响因素[J].中国学校卫生,2008, 29 (9):810~812.
[17]Meador-Woodruff J H. Update on dopamine receptors[J]. Annals of Clinical Psychiatry, 1994,6 (2):79~90.
[18]Andrea S,Camperio C.The Dopamine D4 Receptor (DRD4) Ex-on 3 VNTR Contributes to Adapt/ve Personality Differences in anItalian Small Island Populat/on[J]. European Journal of Personality, 2013(27):593~604.
[19]季卫东,陈思路.DRD4 exonⅢ基因多态性与抽动秽语综合征共病抑郁的关系[J],上海交通大学学报(医学版),2014, 34(4):426~429.
[20]池万余.DDC基因和DRD4基因多态性与人类认知能力的关系研究[D].西安:西北大学,2011.
[21]Lee K Y, JooE J,Ji Y I,et al.Associations between DRDs andschizophrenia in a Korean population: multi- stage associationanalyses[J]. Experimental& Molecular Medicine, 2011, 43(1): 44~52.
