青蛤凝集素CSL对酿酒酵母细胞形态的影响
2018-01-22,,,,*,,*
,,,,*, ,*
(1.东北林业大学盐碱地生物资源环境研究中心,东北油田盐碱植被恢复 与重建教育部重点实验室,黑龙江哈尔滨 150040; 2.河套学院农学系,内蒙古巴彦淖尔 015001; 3.大连海洋大学食品科学与工程学院,辽宁大连 116032)
酿酒酵母(Saccharomycescerevisiae)是人类最早利用的真核单细胞微生物,早在公元前3000年,人类就已经开始利用酵母制作食品。目前人们对酿酒酵母进行的研究,尤其是对其细胞壁结构的研究早已建立在分子水平[1]。研究酵母细胞壁,对更好地利用酵母有着重要的作用[2-6]。酵母细胞壁约占细胞干重的20%~30%,主要由D-葡萄糖和D-甘露糖两类多糖组成,酵母细胞壁会随着周围环境发生变化而发生自我调节,从而适应或抵抗不良环境,保护酵母细胞[7]。
青蛤凝集素(CSL)是一种能够凝集酵母细胞的Ca2+依赖型凝集素,其凝集活性可以被D-甘露糖和N-乙酰-D-半乳糖胺抑制[8]。CSL通过与酿酒酵母细胞壁上的甘露糖相互作用,能够提高酿酒酵母单位时间内的乙醇产量[9]。酵母细胞壁含有肽聚糖,其对维持细胞形态具有重要作用[1]。但目前为止,对于凝集素作用对酵母形态影响的研究还未有报道。本文通过对CSL与酵母相互作用时,分析酵母体积的变化,了解酵母细胞壁的变化规律,并分析CSL与酵母细胞壁肽聚糖的相互作用,有助于了解凝集素与酵母相互作用的机理。
1 材料与方法
1.1 材料与仪器
青蛤凝集素(CSL) 由实验室前期制备所得;酿酒酵母 安琪酵母股份有限公司;辣根过氧化物酶(Horse radish peroxidase,HRP)、牛血清蛋白(BSA) 北京索莱宝公司;肽聚糖(peptidoglycan)、甘露糖(D-Mannose)、N-乙酰-D-半乳糖胺(N-acetyl-D-galactosamine) 美国sigma公司;钙离子定量试剂盒 AAT Bioquest Inc;其他试剂 均为分析纯。
YPD液体培养基(Yeast Extract Peptone Dextrose Medium) 分别取葡萄糖2.0 g、酵母浸粉1.0 g、蛋白胨2.0 g溶于100 mL去离子水中,121 ℃条件下灭菌20 min。
LDZX-30KBS型立式压力蒸气灭菌器 上海申安医疗器械厂;HZQ-C型空气浴振荡器 哈尔滨市东联电子技术开发有限公司;ZDP-A2160A型曲线控制恒温培养箱 上海智城分析仪器制造有限公司;ZHJH-C1109B型超净工作台 上海智城分析仪器制造有限公司;Multiskan MK3型酶标仪 美国Thermo公司;S-4800型扫描电镜 日本Hitachi公司;Q150R ES型离子溅射镀膜仪 英国Quorum公司。
1.2 实验方法
1.2.1 CSL-HRP酶联物的制备 凝集素酶联物制备采用高碘酸钠法[10]。取10.0 mg CSL溶解于1 mL 0.01 mol/L磷酸盐缓冲溶液(pH5.5)备用。取5.0 mg HRP溶解于1 mL超纯水中,再加入0.2 mL 0.1 mol/L高碘酸钠溶液,避光搅拌20 min,在1 mmol/L醋酸钠缓冲溶液(pH4.4)中4 ℃透析12 h。透析结束后,在透析液中加入20 μL 0.2 mol/L 碳酸盐缓冲溶液(pH9.5),之后加入全部CSL备用液,避光搅拌2 h。再加入0.1 mL 4.0 mol/mL硼氢化钠溶液,4 ℃静置2 h。最后在0.2 mol/L磷酸盐缓冲溶液(pH7.4)中透析12 h,制得CSL-HRP。
1.2.2 肽聚糖包被96孔酶标板 将3.0 mg酵母菌肽聚糖溶解于8 mL 0.1 mol/L 碳酸盐缓冲溶液(pH9.5)中,并在每孔中加入肽聚糖溶液100 μL,4 ℃过夜。用含有0.05% Tween 20的0.02 mol/L磷酸盐缓冲溶液(pH7.2)洗涤3次,每次3 min。洗涤后,在每孔中加入200 μL含有0.1% BSA的磷酸盐缓冲溶液(pH7.2),4 ℃过夜。最后再用相同方式洗涤3次,备用[10]。
1.2.3 CSL-HRP固相吸附实验 将1.2.1方法所得的CSL-HRP溶液用超纯水分别稀释至2、1、0.5、0.25、0.125 mg/mL。取各稀释液100 μL加入到1.2.2方法所得的96孔板中,37 ℃反应1 h后,以PBS洗涤3次。加入100 μL含有0.4 mg/mL邻苯二胺及0.01% H2O2的磷酸-柠檬酸缓冲液(pH5.0),37 ℃进行显色反应10 min,用2 mol/L H2SO4终止反应,在492 nm处测定吸光值[10]。
1.2.4 pH对CSL-HRP结合肽聚糖的影响 取90 μL 0.05 mol/L的HAc-NaAc(pH5.0)、NaH2PO4-Na2HPO4(pH6.0)、NaH2PO4-Na2HPO4(pH7.0)、Tris-HCl(pH8.0)、Gly-NaOH(pH9.0)溶液加入到1.2.2方法所得的96孔板中,再加入90 μL CSL-HRP,37 ℃反应1 h后,以PBS洗涤3次后,按照1.2.3显色反应和终止反应方法进行,并测吸光值[10]。
1.2.5 D-Mannose与GalNAc的浓度对CSL-HRP结合肽聚糖的影响 分别取50 μL 0.625、1.25、2.5、5、10 mg/mL的D-Mannose溶液和GalNAc溶液,加入到1.2.2方法所得的96孔板中,再加入50 μL CSL-HRP,37 ℃反应1 h后,以PBS洗涤3次后,按照1.2.3显色反应和终止反应方法进行,并测吸光值[10]。
1.2.6 酵母菌活化 将活性干酵母1.0 g加入到100 mL含2%葡萄糖的无菌水中,35 ℃条件下,活化30 min,得到酵母菌悬液。
1.2.7 酿酒酵母与CSL结合后表面积变化测定 由于对数生长期的酵母细胞新陈代谢水平以及各种代谢物的积累水平都比较稳定,所以本实验选用对数生长期的中间阶段,即酵母生长到12 h时对其进行NaCl和CSL浸泡处理,尽量缩小由取样时间带来的偏差。
将活化后的酵母溶液10 mL加入到100 mL YPD液体培养基中,在30 ℃、150 r/min条件下培养12 h(对数生长期)得到酵母菌悬液。将进入对数生长期的酿酒酵母悬液按照1∶1体积比加入到6种不同浓度的NaCl溶液中,溶液中含有的NaCl终浓度分别为0、20、40、60、80、100 mg/mL,30 ℃浸泡30 min后,置于室温15 min后,利用显微镜测量酵母细胞直径大小,并计算其表面[11]。每个浓度重复3次,每次随机选取5个细胞进行测量。
取进入对数生长期的酵母悬液100 μL分别加入到100 μL不同浓度CSL溶液中,得到CSL最终浓度分别为3.37、6.73、13.45 mg/mL的酵母菌悬液,30 ℃浸泡30 min后,置于室温15 min后,利用显微镜测量酵母细胞直径大小,并计算其表面积。每个浓度重复3次,每次随机选取5个细胞进行测量。
由图2可知,鲜肉泥的红度值随腌制时间的延长而下降,因此,选取腌制3 h时红度值较高的生鲜肉泥用于制作肉脯。
酵母细胞表面积计算,假设酵母细胞为球体。
表面积S=4πR2(R为球体半径)
1.2.8 酵母细胞内钙离子浓度测定 钙离子标准曲线:取300 mmol/L钙离子标准溶液10 μL加入到990 μL稀释缓冲液中,制得3 mmol/L钙离子标准液。再取50 μL 3 mmol/L钙离子标准液加入到950 μL稀释缓冲液中,制得150 μmol/L钙离子标准液,1∶2梯度稀释分别得到浓度为75、37.5、18.75、9.375、4.68、2.34、0 μmol/L的钙离子标准液,在96孔板中分别每孔加入钙离子标准液50 μL及显色液50 μL,室温下避光反应5~10 min,于650 nm处测吸光度值。
酵母细胞裂解缓冲液:分别取EDTA 1.46 g,β-巯基乙醇35 μL,NaCl 0.585 g,甘油10 mL,PMSF(100×)0.5 mL,溶于0.1 mol/L 的Tris-HCl(pH7.5)并定容至50 mL。
苯甲基磺酰氟(PMSF)(100×):取0.1742 g PMSF溶于10 mL异戊醇,室温保存。
取按1.2.6方法所得的菌悬液以及终浓度为13.45 mg/mL CSL浸泡30 min的菌悬液,用0.05 mol/L pH7.4 PBS清洗3次,然后用PBS将菌液调整至OD600值为0.85±0.005,各取75 mL离心收集酵母细胞,悬浮于1 mL酵母细胞裂解缓冲液中,超声破碎30 min(超声功率300 W,工作1 s,间歇0.5 s)。4 ℃ 10000 r/min离心15 min,取上清液使用钙离子定量试剂盒于650 nm处测吸光度[12]。钙离子浓度标准曲线的绘制及样品中钙离子浓度的测定均平行3次实验。
1.2.9 扫描电镜观察 按1.2.6方法所得的菌悬液及终浓度为13.45 mg/mL CSL浸泡30 min的菌悬液,于4 ℃ 4000 r/min条件下离心10 min,所得菌体用0.05 mol/L pH7.4的PBS清洗3次,之后沉淀物用1 mL 2.5%戊二醛固定液,于4 ℃固定12 h。固定后的菌悬液,以0.05 mol/L pH7.4的PBS清洗3次,沉淀以0.05 mol/L pH7.4的PBS 制成菌悬溶。
取200 μL菌悬液滴加到盖玻片上,在培养皿中自然沉降2 h。依次以50%、70%、80%、90%、100%叔丁醇溶液脱水,脱水间隔10 min。100%叔丁醇脱水2次后,再加入100%叔丁醇于4 ℃固化,真空冷冻干燥。将干燥后的菌体切割成3 mm×6 mm小块,以导电双面胶粘贴于铜台上,用喷金仪进行喷金,置于扫描电镜中进行观察(10000倍)[13]。
2 结果与分析
2.1 CSL与肽聚糖的相互作用
2.1.1 固相吸附 将酵母细胞壁肽聚糖与新制备的不同浓度的CSL-HRP溶液在96孔板板上进行结合测试,结果见图1。由图1可知,CSL与肽聚糖可以结合,并随着CSL浓度的增加,CSL与肽聚糖结合的量也在增加,具有浓度依赖关系。

图1 CSL固相吸附Fig.1 Binding ability of CSL to peptidoglycan
2.1.2 pH对CSL与肽聚糖结合的影响 pH对CSL与肽聚糖结合的影响结果见图2。由图2可知,当pH为6时,CSL结合肽聚糖的量最高,但随着pH的上升,CSL与肽聚糖的结合能力逐步下降。结果表明,CSL与肽聚糖的结合受pH的影响,在pH为6时结合能力最强。

图2 pH对CSL与肽聚糖结合的影响Fig.2 Effect of pH on binding ability of CSL to peptideglycan
2.1.3 GalNAc对CSL与肽聚糖结合的影响 GalNAc对CSL与肽聚糖结合的影响结果见图3。由图3可知,随着GalNAc浓度的增加,CSL结合肽聚糖的能力逐渐下降。结果表明,CSL与酵母细胞壁肽聚糖的结合受到GalNAc的抑制,而CSL凝集活性又可以被GalNAc抑制,说明CSL上有GalNAc结合位点[13]。
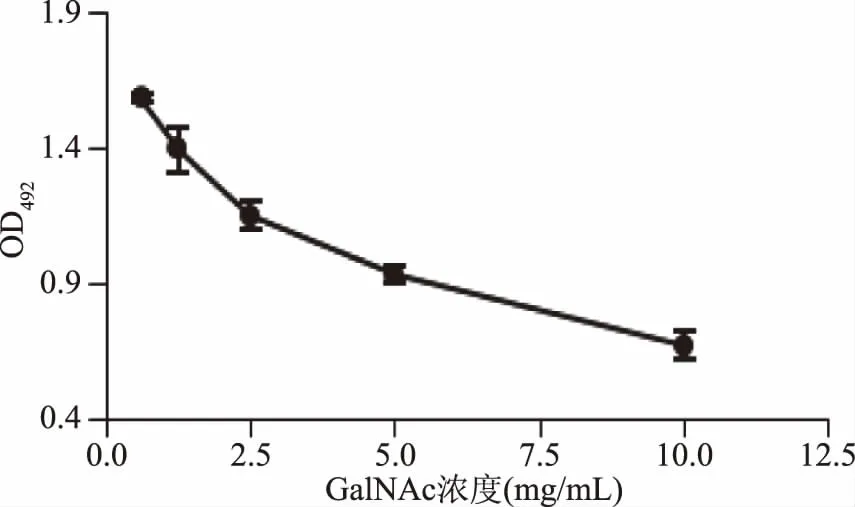
图3 GalNAc对CSL与肽聚糖结合的影响Fig.3 Effect of GalNAc on binding ability of CSL to peptideglycan
2.1.4 D-Mannose对CSL与肽聚糖结合的影响 D-Mannose对CSL与肽聚糖结合的影响结果见图4。由图4可知,随着D-Mannose浓度的增加,CSL结合肽聚糖的能力逐渐下降。结果表明,CSL与酵母细胞壁肽聚糖的结合受到D-Mannose的抑制,而CSL凝集活性又可以被D-Mannose抑制,说明CSL上有D-Mannose结合位点[13]。
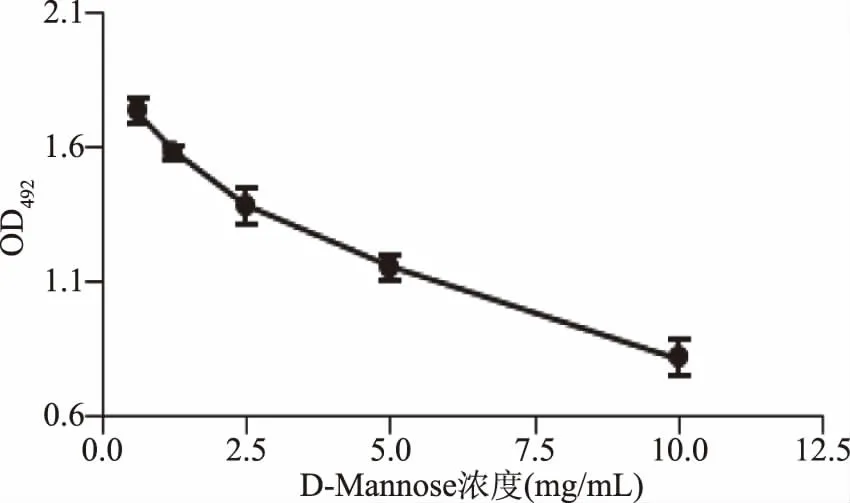
图4 D-Mannose对CSL与肽聚糖结合的影响Fig.4 Effect of D-mannose on binding ability of CSL to peptideglycan
Tong等[14]研究凝集素MCL-T时发现,该凝集素结合OMP时,受到PSM及GalNAc的抑制。一般而言,凝集素具有专一的糖结合位点,而CSL与肽聚糖的结合受到GalNAc、D-Mannose的抑制说明,肽聚糖结合位点与GalNAc、D-Mannose的结合位点分别存在于凝集素上不同的结构域中,CSL与GalNAc、D-Mannose的结合导致了其与肽聚糖的结合,导致结合减少。
2.2 酿酒酵母细胞盐胁迫条件下及青蛤凝集素CSL作用下表面积变化
酿酒酵母细胞在不同环境条件下其代谢行为发生改变,在这一应激过程中,酵母细胞可能发生了一系列的变化,包括细胞大小、表面积等的变化,因此本实验设计了青蛤凝集素与酿酒酵母共培养以观察细胞表面积变化。
将酿酒酵母细胞放大1000倍后,在100 mg/mL NaCl盐胁迫条件下表面积变化如图5所示。图5A为在正常条件下酵母细胞大小,图5B为在NaCl浓度100 mg/mL条件下酵母细胞大小。由图5可见,NaCl在浓度为100 mg/mL时会导致酿酒酵母细胞表面积变小。

图5 酿酒酵母细胞在100 mg/mL NaCl胁迫条件下细胞表面积(1000×)Fig.5 Surface area of the yeast against 100 mg/mL NaCl(1000×)注:A:对照组酿酒酵母细胞; B:100 mg/mL NaCl胁迫下的酿酒酵母细胞。
不同浓度NaCl环境中酵母细胞表面积大小见图6。由图6可见,NaCl浓度在小于20 mg/mL时,酿酒酵母细胞表面积无明显变化。NaCl浓度在大于40 mg/mL时,酿酒酵母细胞随着NaCl浓度的增加,酵母细胞收缩,液泡缩小,细胞的表面积缩小。

图6 在NaCl胁迫条件下酿酒酵母细胞表面积的变化Fig.6 Surface area change of yeast cell during exposure to NaCl solution
酿酒酵母细胞在CSL作用下表面积变化如图7所示。图7A、7B、7C、7D分别为CSL浓度0、3.37、6.73、13.45 mg/mL条件下的酵母细胞显微镜下照片。结果表明,CSL可以致使酿酒酵母细胞表面积发生改变。
不同浓度CSL环境中酵母细胞表面积的大小见图8。由图8可知,正常条件下,酵母细胞表面积的平均值约为89.21 μm2。在CSL的作用下,酿酒酵母细胞表面积明显缩小,并随CSL浓度增加而减小,在CSL浓度增加到3.37、6.73、13.45 mg/mL条件下,细胞表面积平均值分别为81.16、72.94及69.94 μm2,这表明CSL促使酿酒酵母细胞壁发生了改变,导致了酿酒酵母细胞的表面积随着CSL浓度的增加而变小。

图8 不同CSL浓度条件下酵母细胞表面积的变化Fig.8 Surface area change of yeast cell during exposure to different concentration of CSL solution
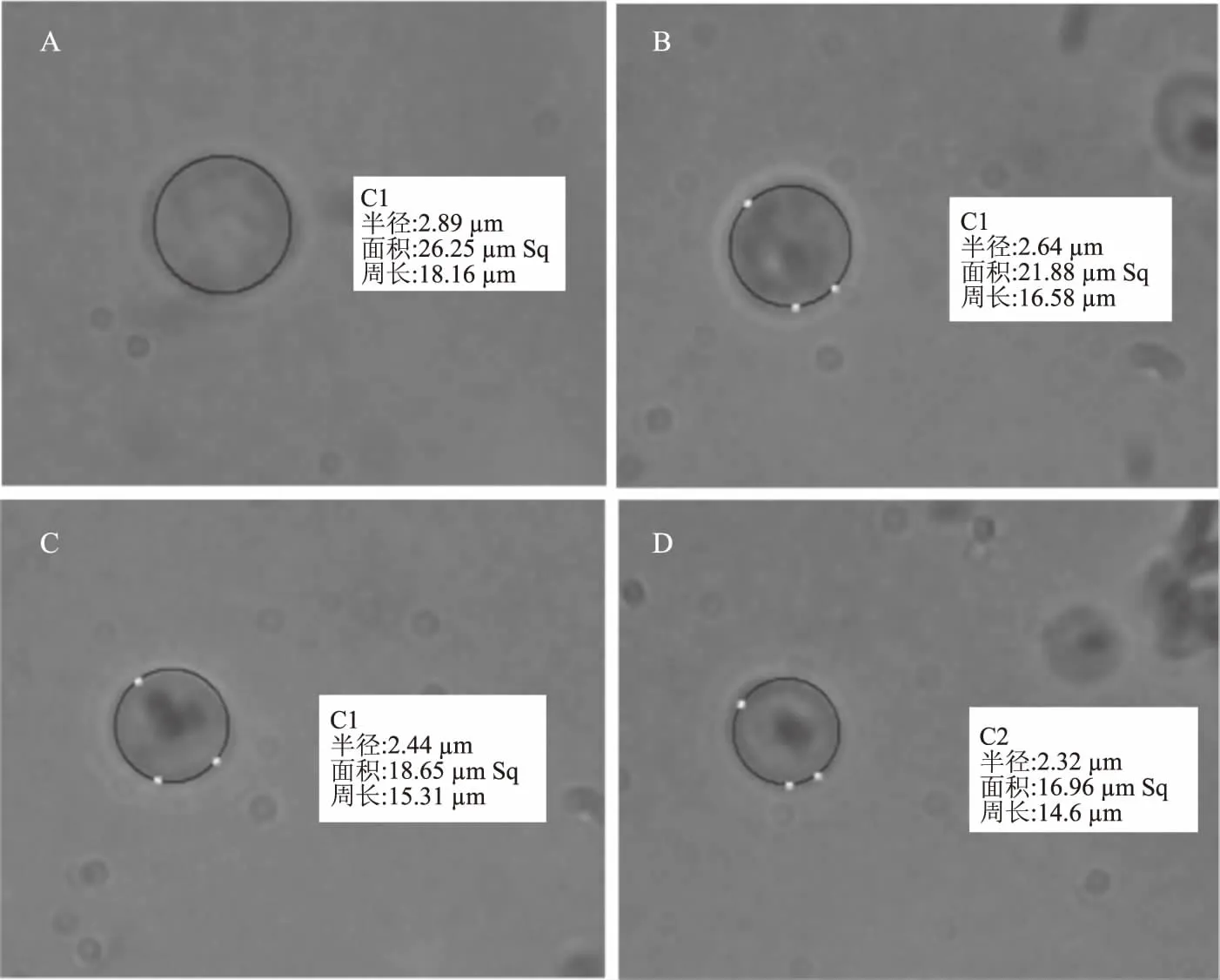
图7 CSL对酿酒酵母细胞表面积的影响(1000×)Fig.7 Effect of CSL on surface area of Saccharomyces cerevisiae(1000×)注:A:对照组酿酒酵母细胞;B:CSL为3.37 mg/mL时的酿酒酵母细胞; C:CSL为6.73 mg/mL时的酿酒酵母细胞;D:CSL为13.45 mg/mL时的酿酒酵母细胞。
NaCl浓度的增加会导致酵母细胞体积减小,说明此种酵母细胞的体积及表面积会在周围环境发生变化的条件下进行适应性改变。Tong等与Liu等已证明在酿酒酵母发酵过程中加入CSL能够凝集酿酒酵母细胞、促进其增殖和提高乙醇产量[8-9]。本实验加入CSL导致酵母细胞体积及表面积减小,说明CSL与酿酒酵母细胞壁相互作用,使酿酒酵母的细胞壁发生了变化,从而使外源信号传递到酿酒酵母胞内,使代谢途径产生了相应的响应,提高了酵母细胞产生乙醇能力。
2.3 酵母细胞内钙离子浓度
钙离子浓度标准曲线见图9。由图9可知,使用钙离子定量试剂盒绘制的钙离子浓度标准曲线浓度范围为0~75 μmol/L,其浓度和吸光度呈良好的线性关系,相关系数R2大于0.994。通过该方法测得对照组Ca2+浓度为24.0949 μmol/L,凝集素添加组钙离子浓度为24.3513 μmol/L,与对照组相比,无显著性差异(p<0.05)。
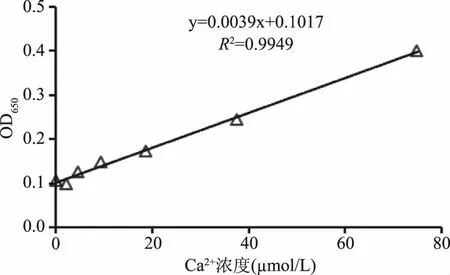
图9 钙离子浓度标准曲线Fig.9 The standard curve of calcium ion
酵母细胞中包括盐、环境压力和营养物感应等许多方面的Ca2+信号已被充分研究,Ca2+作为真核细胞内广泛存在的细胞内信使调控许多细胞过程。酵母细胞外Ca2+一般情况下是通过一个未知的转运蛋白X(transporter X)进入到细胞质内,当外界环境中Ca2+浓度较高时还会有转运蛋白M(transporter M)参与[15]。通过研究添加CSL后,酵母细胞内Ca2+浓度变化来考察CSL信号的传递途径。
本实验通过钙离子定量试剂盒,对单位时间内乙醇产量发生变化的对照组与CSL添加组两组酵母菌内的Ca2+含量进行了测定,发现两组酵母菌内的钙离子含量未发生明显变化,说明CSL刺激酵母菌产乙醇可能与酵母细胞内钙离子通路相关性不强,CSL与酿酒酵母细胞壁的相互作用可能并未影响到transporter X。
2.4 电镜扫描结果
通过电镜扫描观察对照组与CSL添加组的酵母形态见图10,发现这两者具有较明显差异。由图10A能观察到对照组酵母细胞表面较圆润光滑,而图10B中CSL添加组酵母细胞表面有较多赘附凸起。电镜扫描结果提示,CSL使酿酒酵母的细胞壁表面形态发生了变化,这一变化与CSL促进了酵母细胞的发酵能力的提高[8-9]有一定的内在联系。

图10 酵母电镜扫描照片Fig.10 Scanning electron micrographs of yeast 注:A:对照组酿酒酵母细胞;B:CSL组酿酒酵母细胞。
3 结论
通过研究发现,CSL可与酵母细胞壁肽聚糖结合且两者的结合具有浓度依赖关系,该结合位点受甘露糖(D-Mannose)及N-乙酰-D-半乳糖胺(N-acetyl-D-galactosamine)的抑制。显微镜观察酿酒酵母细胞在盐胁迫条件和青蛤凝集素CSL作用下的细胞状态研究发现,盐胁迫和CSL分别与酿酒酵母细胞壁相互作用,均会导致酿酒酵母细胞的表面积发生改变,说明盐胁迫和CSL均会改变酿酒酵母细胞的细胞壁。对照组与CSL添加组两组酵母菌内的Ca2+含量未发生明显变化,说明CSL刺激酵母菌产乙醇可能与酵母细胞内钙离子通路相关性不强。通过电镜扫描观察对照组与CSL添加组的酵母形态,发现对照组酵母细胞表面较圆润光滑,CSL添加组酵母细胞表面有较多赘附凸起。研究结果为CSL促进酵母发酵能力的提高机理提供了基础数据。
[1]Klis FM,Boorsma A,De Groot PWJ. Cell wall construction in Saccharomyces cerevisiae[J]. Yeast,2006,23(3):185-202.
[2]钱潘攀,李红波,赵岩岩,等. 双效型酵母细胞壁提取物及其对黄曲霉毒素B1吸附特性研究[J]. 食品工业科技,2017,(15):1-12.
[3]Mokhtari S,Jafari SM,Khomeiri M,et al. The cell wall compound of Saccharomyces cerevisiae as a novel wall material for encapsulation of probiotics[J]. Food Research International,2017,96:19-26.
[4]Sultana A,Miyamoto A,Hy QL,et al. Microencapsulation of flavors by spray drying usingSaccharomycescerevisiae[J]. Journal of Food Engineering,2017,199:36-41.
[5]杨婷,祝霞,李颍,等. 葡萄酒泥酵母β-葡聚糖提取工艺条件优化[J]. 食品工业科技,2015,36(18):286-289,309.
[6]王辉田,孙超,陈思,等. 酵母细胞壁多糖对肉仔鸡生长性能及免疫力的影响[J]. 中国饲料,2013(13):11-14.
[7]Klis FM,Mol P,Hellingwerf K,et al. Dynamics of cell wall structure inSaccharomycescerevisiae[J]. FEMS Microbiology Reviews,2002,26(3):239-256.
[8]Tong C,Li W,Kong L,et al. A novel yeast-binding lectin from hemolymphCyclinasinensis(Gmelin)and its effects on yeast cells[J]. Process Biochemistry,2012,47(12):2166-2171.
[9]Liu S,Li L,Tong C,et al. Quantitative proteomic analysis of the effects of a GalNAc/Man-specific lectin CSL on yeast cells by label-free LC-MS[J]. International Journal of Biological Macromolecules,2016,85:530-538.
[10]Valentina M,Irina C,Li W,et al. New GlcNAc/GalNAc-specific lectin from the ascidian Didemnum ternatanum[J]. Biochimica et Biophysica Acta General Subjects,2005,1723(1-3):82-90.
[11]Morris GJ,Winters L,Coulson GE,et al. Effect of osmotic stress on the ultrastructure and viability of the yeastSaccharomycescerevisiae[J]. Journal of General Microbiology,1986,132(7):2023-2034.
[12]胡国武,李景川,元英进. 稀土对红豆杉细胞内游离钙含量的影响[J]. 天津大学学报,2000,33(1):17-20.
[13]佟长青. 两种海洋双壳贝类凝集素及其抑菌机制的研究[D]. 哈尔滨:东北林业大学,2013.
[14]佟长青,李琦,曲敏,等. 菲律宾蛤仔凝集素抑菌机制的研究[J]. 大连海洋大学学报,2014,29(5):508-513.
[15]赵运英,蒋伶活. 酿酒酵母细胞中钙离子信号传导途径的研究进展[J]. 中国细胞生物学学报,2013,35(10):1504-1513.
