谷物中γ-氨基丁酸(GABA)富集工艺的研究进展
2018-01-22,,,,2,*
,,,,2,*
(1.上海交通大学农业与生物学院食品科学与工程系,上海 200240;2.上海交通大学陆伯勋食品安全研究中心,上海 200240)
γ-氨基丁酸(γ-aminobutyric acid,简称GABA)是一种四碳原子组成的非蛋白质氨基酸,主要由谷氨酸经谷氨酸脱羧酶(GAD)催化转化而来。其在哺乳动物中枢神经系统中作为抑制性神经递质、参与脑循环生理活动,具有降血压、抗心律失常、利尿、镇痛和缓解焦虑等功能[1]。还有研究发现,GABA与创伤后应激障碍(PTSD)[2]、精神分裂症[3]、肌纤维痛和其他中枢性疼痛综合征存在密切关联,并且GABA功能障碍与躁郁症的产生存在联系[4]。Yoto A 等[5]的人群实验研究证明,口服GABA能缓解脑力劳动后的精神压力。Hayes D J等[6]提出大脑GABA与神经精神疾病中的核心问题冲动性有关。此外,有研究指出,GABA能改善氧化应激与甲状腺激素的功能,并减轻体重,可控制肥胖[7]。可见,GABA是人类神经系统中重要的活性物质,具有多种有益的保健功能。日本厚生劳动省2001年正式将GABA列入食品级,我国卫生部也于2009年9月将GABA列入新资源食品,批准其用于食品生产和加工。
虽然GABA在人体中可由谷氨酸在专一性较强的谷氨酸脱羧酶(GAD)作用下催化而成,但仅从天然食物中摄取GABA的量不能满足人体生理需要;另外,尽管GABA在微生物、植物、昆虫和哺乳动物组织中分布广泛[8],但因其含量低、提取技术不完善,提取难度与成本相对较高等原因,加大富含GABA食品的开发具有重要意义。
目前,生物化学途径富集谷物GABA的方法主要有植物代谢法和微生物发酵法,富集所用原料来源丰富,包括麦类、稻类、粟米等,富集方法主要包括厌氧培养、溶液浸泡、恒温发芽以及用细菌、霉菌和酵母等微生物进行发酵处理等。本文将主要就谷物类原料的富集工艺、富集效果及工艺的优缺点进行阐述。
1 植物代谢法富集谷物GABA
植物代谢法是对食品原料进行浸泡、萌芽、厌氧、低温刺激等处理,通过影响其中的GAD酶活性,催化由内源性蛋白水解成的谷氨酸脱羧合成GABA[9]。植物代谢法主要有生长代谢与应激代谢,生长代谢富集法主要利用植物发芽过程中一系列物质与酶活性的变化积累GABA;应激代谢则利用低氧、低温、干旱、高H+浓度等逆境处理方法富集GABA。植物代谢法主要工艺为先浸泡后发芽,浸泡工艺中需要注意时间、温度、所用浸泡溶液的种类;发芽工艺中则需控制温度、时间和氧气条件等。
1.1麦类
大麦中含有丰富的蛋白质和GAD酶。有研究[10]以制作麦芽的大麦为原料,研究了大麦浸泡及发芽过程中GABA含量和GAD酶活性的变化,发现大麦浸泡5 h后,断水放置7 h,如此循环处理4次,在温、湿度分别为15 ℃,95%的条件下发芽,结果表明浸泡完成时GAD酶活性最低,发芽24 h时GABA含量最高,达到原料的6.5倍。因此,大麦发芽过程出现GAD活性变化,存在富集GABA的可能性。Jin W J等[11]利用大麦和小麦麸皮中的GAD,以谷氨酸为底物,磷酸吡哆醛为辅因子,进行酶促反应富集GABA,得到的最佳工艺条件:大麦麸皮加入10 μmol/L磷酸吡哆醛和10 mmol/L谷氨酸,30 ℃摇床反应12 h;小麦麸皮中加入50 μmol/L磷酸吡哆醛和10 mmol/L谷氨酸,30 ℃摇床反应36 h,GABA产量分别为10.4 mmol/L和7.3 mmol/L。此外,浸泡大麦后进行发芽培养,能继续提高GABA富集量,25 ℃下浸泡大麦24 h后,15 ℃恒温发芽36 h,GABA含量达到20.9 mg/100 g,是原麦的14.6倍[12]。另外,也有研究[13]指出浸麦过程采用50 mmol/L醋酸钠溶液浸泡能更好地富集GABA,该研究还发现避光氮气厌氧处理青稞12 h后,GABA含量可达到14.3 mg/100 g,较原料(3.7 mg/100 g)增加了3倍。对大麦浸泡发芽过程GABA积累的现象,可认为是植物对环境压力的应激反应,Kleinwächter M等[14]的实验证实,在低氧环境中萌发的大麦种子会富集更多的GABA,在80% CO2和20% O2环境下发芽的大麦中GABA积累甚至超过50 mg/100 g(正常组8 mg/100 g)。另外,除大麦、小麦等常见品种,荞麦也存在发芽富集GABA的可能,苦荞在发芽4 d时叶和胚中GABA含量达到最高,分别为0.2305 mg/g和0.1549 mg/g,是第一天的1.54和1.32倍,之后出现平缓下降趋势,子叶和胚中GAD活力均呈先升后降趋势[15]。
1.2糙米
糙米于水中浸泡并发芽后形成发芽糙米(Germinated brown rice,简称GBR),被认为比白米更健康,因为它不仅富含维生素、矿物质、膳食纤维和必需氨基酸等基本营养成分,而且含有更多的生物活性成分,如阿魏酸、γ-谷维素和γ-氨基丁酸等[16]。关于发芽糙米的研究始于日本,起步较早研究背景深远,对于浸泡和发芽条件的研究较为成熟,市场上也已出现发芽糙米相关产品。Karladee D等[17]测试了五个不同发芽时间(0、12、24、36和48 h)和21个品种在发芽过程中GABA的变化,结果显示GABA平均含量从0 h的3.96 mg/100 g持续增加到12 h的10.04 mg/100 g,在24 h达到最高点 17.87 mg/100 g,之后缓慢下降为5.91 mg/100 g(36 h)和1.36 mg/100 g(48 h)。为了研究糙米不同部位在发芽过程中营养成分的变化,Kim H Y等[18]将糙米分为壳体、糙米和芽,发现各部分GABA含量在糙米萌发后分别提高到3.34、26.84和6.04 mg/100 g,糙米整体GABA含量从15.34增加到31.79 mg/100 g。此外,浸泡溶液的变化也会影响糙米GABA的含量,用电解水预浸泡和超声波处理糙米可显着提高其抗菌活性,同时酸性电解水比碱性电解水更有利于GABA的积累,发芽第4天,与自来水相比,使用OPR 195.9 mV(pH8.9)和ORP-845.45 mV(pH11.95)的碱性电解水处理,GABA含量分别下降11.59%和38.10%,而使用OPR 1024.1 mV(pH3.27)和OPR 1088.1 mV(pH3.03)的酸性电解水处理,GABA含量分别提高了39.76%和3.61%[19]。也有研究[20]使用pH5.6的酸性溶液(柠檬酸与柠檬酸钠调节)并且浸泡时添加0.25 mg/L的赤霉素和1 g/L的L-谷氨酸,发现发芽糙米GABA含量显著提高。值得注意的是对发芽糙米进行干燥处理并不会影响GABA含量,Chungcharoen T等[21]的糙米发芽后GABA含量从4.2 mg/100 g增加到22.8 mg/100 g,并且当发芽样品通过热空气流化床干燥器在130或150 ℃干燥后,GABA含量并不降低。此外,有研究[22]发现有色糙米和无色糙米发芽过程中生育酚、生育三烯酚、γ-谷维素和GABA含量显示出一定差异,无色糙米发芽后,GABA含量高达62.3 mg/kg(未处理无色糙米中含量为25.6 mg/kg),是同条件有色糙米的1.59倍。
1.3米胚芽
米胚芽是稻米加工的副产品,一般混在米糠中,可利用比重差选法分开。米胚芽中含有丰富的GAD,王仁辰等[23]利用米胚芽粉末作为GAD酶来源,得出最佳反应条件为100 mmol/L MSG、0.2 mmol/L磷酸吡哆醛且pH5.1的反应液中,26 ℃下反应6 h,GABA产量4440.32 μg/mL。也有研究[24]在米胚芽的浸泡液中添加不同种类的蛋白酶和50 mmol/L的磷酸缓冲液,于各类蛋白酶最适pH和温度下培养6 h(胰蛋白酶45 ℃ pH8.0,复合蛋白酶45 ℃ pH8.0,碱性蛋白酶45 ℃ pH7.0),结果显示碱性蛋白酶处理最有利于GABA含量的提高。此外,为进一步优化米胚芽加工工艺,延长产品保质期,提高风味口感,有研究[25]对米胚芽进行脱脂处理,将油脂含量从21.5%降到3.9%后,可使GABA含量从251 mg/100 g上升至328 mg/100 g。
1.4粟米
粟米是最早种植的作物之一,广泛种植在亚洲和非洲的干旱和半干旱地区,含有丰富的蛋白质、纤维、矿物质和植物化学物质,具有降血脂、低血糖指数和抗氧化特性[26]。Sharma S等[27]研究了发芽对稗小米营养成分的影响,得出最佳发芽条件为浸泡时间11.78 h,发芽温度33 ℃,发芽时间36.48 h,此时GABA含量从3.38 mg/100 g增加到35.70 mg/100 g。进一步的研究[28]发现,科多粟米的富集条件为浸泡时间13.81 h,发芽温度38.75 ℃和发芽时间35.82 h,此条件下GABA含量从9.36 mg/100 g增加到47.43 mg/100 g。
注:关于富集倍数的描述,括号中为对照;明确标注者除外,其余浸泡均为水中浸泡。另外,浸泡溶液的变化同样影响粟米中GABA的富集,将狐尾粟25 ℃蒸馏水浸泡8 h后,移至含有1.2 mg/mL谷氨酸,50 μmol/L磷酸吡哆醛和2.5 mmol/L氯化钙溶液中,32 ℃避光厌氧培养,发芽后GABA含量高达42.9 mg/100 g[29]。此外,氯化钠处理被发现可提高发芽狐尾粟的GABA含量影响,狐尾粟在25 ℃浸泡8 h后25 ℃萌芽48 h,再放入不同浓度的氯化钠溶液中25 ℃培养48 h,其中100 mmol/L的氯化钠中GABA积累达到峰值23.58 mg/100 g(纯水对照15.98 mg/100 g),同时100 mmol/L氯化钠和5 mmol/L Ca2+的联合处理使得GABA富集量比单独氯化钠处理提高了93.94%[30]。
1.5玉米
玉米作为世界上最重要的粮食之一,营养丰富,食用方式广泛。Paucar-Menacho L M等[31]研究了紫玉米发芽时间和温度对GABA含量的影响,发现浸泡24 h后26 ℃下发芽63 h是最佳的发芽条件,与种子相比GABA含量增加了3.5倍。此外,玉米中也存在厌氧发芽刺激GABA积累的情况,有研究[32]发现浸泡6 h后35 ℃厌氧发芽48 h后紫玉米和白玉米GABA含量分别达到37.2 mg/100 g和54.47 mg/100 g(未发芽分别为2.68 mg/100 g和1.58 mg/100 g)。
综上所述,在植物代谢法富集谷物食品中GABA的研究中,所用原材料种类较多,不同原料GABA的富集效果差异较大,所需条件有所不同。如麦类富集倍数为原料的6倍左右,而米胚芽则较未发芽前增加80倍左右(如表1所示)。麦类作物通过浸泡可通过激活GAD酶活性富集GABA,进一步发芽可使GABA持续增加,发芽温度较低,一般为15 ℃左右,这可能与其耐寒特性有关。与麦类相似,糙米经浸泡和发芽后,GABA也得到富集。但糙米浸泡和发芽的温度一般为30到37 ℃之间,这说明不同谷物的最佳GABA富集温度不完全取决于GAD酶的最适温度,而与植物本身的特性也有一定关系。
浸泡溶液的组成对GABA的富集效果也存在显著影响,其中由于磷酸吡哆醛能显著影响GAD活性,在谷物的浸泡及其发芽过程中,其浓度可影响GABA的积累量[33]。还有研究在浸泡液中加入蛋白酶,这是由于米胚芽不是完整的植物种子,在浸泡过程中不能依靠自身的蛋白酶分解蛋白,为了提高GAD的底物浓度,适当添加蛋白酶可增加谷氨酸含量。在利用植物代谢法富集GABA的过程中,高温,缺水,物理损伤,生物胁迫等也可使谷物中GABA的含量增加,因此可根据谷物自身特性选择合适的诱导因素进行处理,以建立最佳GABA富集条件[34]。
此外,植物代谢法富集GABA,虽工艺步骤明晰,操作简单,生产成本低廉,可选择的原料种类广泛,但其生产周期较长,且需要考虑发芽率等问题,原料利用率受限。利用微生物发酵法富集谷物中的GABA是另一个可行且高效的方法。
2 微生物发酵法富集谷物GABA
微生物发酵法富集谷物中的GABA,通常是以富含谷氨酸的谷物,如糙米、大豆、小麦等为原料,利用乳酸菌、曲霉菌或酵母等食品安全级微生物发酵富集GABA[35]。
2.1麦类
有研究以燕麦为发酵底物,利用米曲霉菌生产豆豉燕麦:燕麦浸泡8 h后磨碎,调整水分含量至35%,接种曲霉菌(106孢子/g)后再用生理盐水调整水分含量至45±5%,25 ℃静置发酵至72 h,GABA含量从57.1 mg/kg上升至435.2 mg/kg[36]。此外,米曲霉也可用于荞麦发酵富集GABA:原麦浸泡2 h后,接种米曲霉(106孢子/g),在27 ℃条件下发酵48 h,GABA含量达到最大值3.1 mg/g[37]。
2.2糙米
糙米营养丰富,食疗价值高,是近来热门的健康食材。有研究[38]以糙米粉为原料,接种高产GABA的乳酸菌进行发酵,优化后工艺条件为:糙米粉接种乳酸菌后在30 ℃发酵3 d,GABA含量比原糙米粉提高了1倍,达到25 mg/100 g;与未添加葡萄糖相比,添加少于2%浓度葡萄糖对发酵产GABA有抑制作用;添加谷氨酸钠对发酵有促进作用,浓度大于0.33%的谷氨酸钠可以使发酵糙米粉GABA含量达到500 mg/100 g.比糙米原料提高了50倍。进一步研究[39]发现,短乳杆菌P-14富集糙米GABA效果更佳,最佳富集条件为:谷氨酸钠(MSG)添加量1%,发酵温度35 ℃,发酵时间3 d,得到GABA 含量为851.24 mg/100 g,比糙米原料提高了80倍。此外,对糙米酒生产过程的研究[40]发现,酒曲发酵也能富集糙米中的GABA,在发酵温度为34~36 ℃时GABA生成量升高明显,发酵至132 h达到最高点,36 ℃发酵132 h时,GABA的含量达到最高为0.84 g/L。
2.3米糠
米糠作为稻米加工副产物,具有极高的利用价值。杨丽丽等[41]使用乳酸菌复合发酵米糠,以菌种、菌种添加量、发酵温度、发酵时间为单因素研究其对米糠发酵液中γ-氨基丁酸(GABA)含量的影响,最优条件为:在混合菌种嗜热链球菌S1添加量为1.55%,保加利亚乳杆菌L1添加量为1.45%,50 ℃下发酵米糠14 h,发酵液中GABA含量最高,为287.975 mg/100 g。还有研究[42]利用米曲霉对膨化米糠进行液态发酵,结果表明最佳发酵工艺为:料液比1∶10、接种量5%、温度28 ℃、培养时间3 d、摇床转速130 r/min,米糠发酵物中GABA含量达到234.1 mg/100 g米糠,比初始米糠发酵物中的含量提高约2.4倍。
2.4其他谷物
除了常见的麦类、糙米和米糠等谷物,近年来也有研究关注其他谷物中GABA的富集。姚世聪等[43]研究了三株不同丝状真菌(米曲霉,米根霉 3.1175 和米根霉 3.2751)发酵对小米营养成分及 GABA 含量的影响,发现米曲霉发酵的小米中GABA含量随着发酵时间的增加而显著提高(p<0.05),在48 h时达到最大值1 mg/g,之后基本维持不变。此外,有研究[44]将小米加工副产品小米糠作为培养基原料,添加L-谷氨酸钠合成底物,利用短乳杆菌发酵产GABA,在小米糠70 g/L,谷氨酸钠50 g/L,pH4.00,30 ℃,培养72 h的条件下,GABA产量可达32.037 g/L。还有研究[45]利用乳杆菌发酵米胚芽,在米胚芽基础培养基上添加8%葡萄糖、1%酵母膏、0.6%菊粉,以3%接种量在46 ℃下发酵36 h,发酵液中GABA含量可达3.11 g/L。另外,玉米同样可通过微生物发酵法富集GABA,李婷等[46]利用酶解法制备玉米低聚肽,并以其为基质经植物乳杆菌NDC75017进行发酵,以3%(v/v)的接种比例将发酵剂接种于混合基质中(质量浓度80 g/L的甜乳清粉、体积分数为 50%玉米肽 和浓度为20 mol/L的磷酸吡哆醛),30 ℃下发酵68 h后,GABA含量达到15.97 mg/100 mL。
另外,还有报道将植物代谢法和微生物发酵相结合,利用微生物发酵经过植物代谢法处理的原料,以进一步改善GABA的富集效果[47]。魏一星等[48]利用植物乳杆菌(植物乳杆菌dy-1)发酵小麦胚芽,GABA含量较麦胚中的含量增加近7倍。同时,利用发芽糙米发酵富集GABA时,其含量增加约4~6倍[49]。陈璐等[50]还利用萌芽米为原料,选用食用红曲菌M1、M2,酿酒酵母Sce01和乳酸菌Lac01四种菌株进行混合发酵,也取得了一定的富集效果。
综上所述,近年来利用微生物发酵法富集GABA的研究中多用乳酸菌、曲霉菌等。表2所示为微生物发酵谷物富集GABA的主要底物、菌株、发酵温度、发酵时间以及GABA产量。微生物发酵法中菌株的特性极大程度的限制了后续发酵工艺的参数范围,如乳酸菌类菌株的发酵工艺,温度大多控制在37 ℃左右,而米曲霉发酵工艺中发酵温度则控制在25 ℃左右。同时,菌种的选择也直接影响富集效果和产品风味,其中,乳酸菌发酵产物多含有乳酸,米曲霉发酵则可能产生米酒风味。
在菌种选择方面,谷物不同选用的菌种也有所不同,一般来说,稻米多选用红曲霉,糙米多用乳酸菌,而麦类则选用米曲霉等。此外,不同原料其发酵效果也存在较大差别,这是由于当原料本身的成分构成,如谷氨酸含量、碳源和氮源等不同时,其发酵GABA的富集效果也有所不同。微生物发酵法富集GABA,生产周期较短,流水线机械化程度高,适合于工业生产。原料可选用农产品加工副产物或食用价值较低的作物,生产成本较低。发酵产物可直接开发食用产品,如混合植物酸奶饮料[51],也可收集发酵液,喷雾干燥或提取GABA,制成食品添加剂,供新型食品研发。
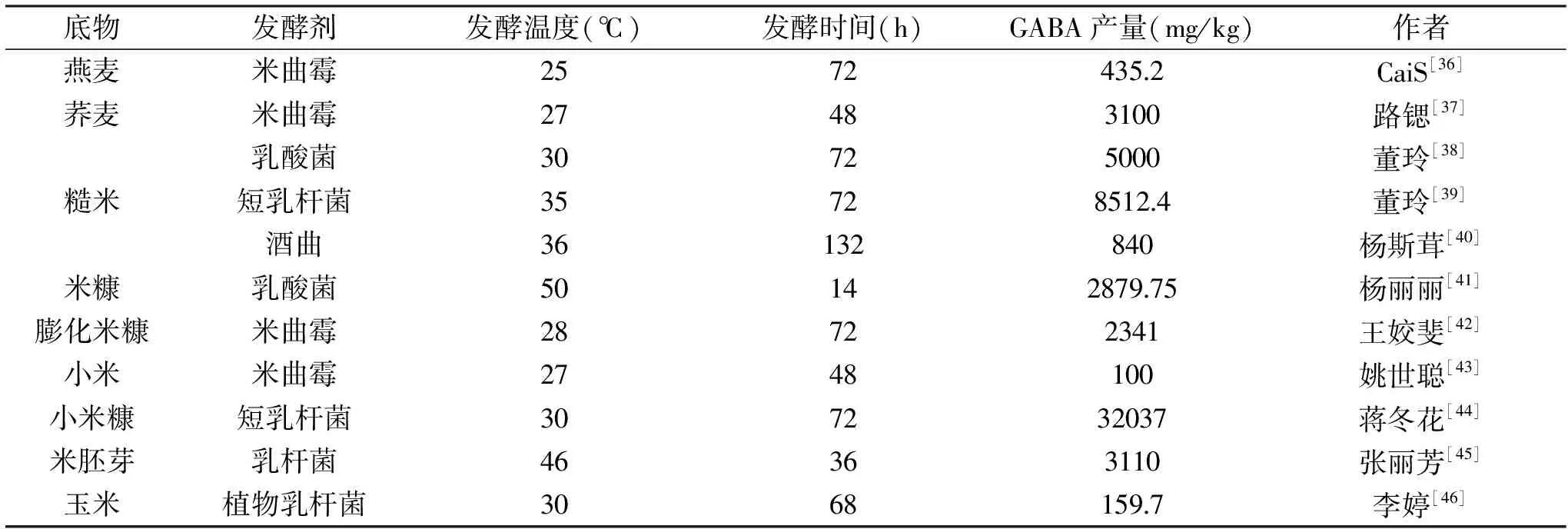
表2 微生物发酵法富集GABA的比较Table 2 Comparison of enrichment of GABA by microbial fermentation
3 前景与展望
目前,越来越多的研究发现GABA除作为抑制性神经递质外,具有较多的生理功能,如何从天然食物中摄入满足生理需求的GABA一直受到关注。食品原料,特别是糙米中的GABA富集技术的研究,在日本等国家已经取得了较多的研究结果,但是如何利用其它谷物食品及食品加工副产物进行GABA的富集生产亟待研究。另外,现有的GABA富集技术还需进一步改良,植物代谢法过度依赖原料本身的GAD,对一些GAD活性不高或含量不足的原料无法有效富集。对此,可考虑通过基因工程提高GAD酶活性以富集不同原料中的GABA。微生物发酵法中高产GABA菌种主要应用于培养基发酵,如何利用不同的食品原料进行GABA的富集,以及如何通过改变发芽条件和添加外源性因子刺激谷物发芽过程中GABA积累,从而开发出富含GABA的天然功能性食品是一个值得研究的方向。此外,关注筛选诱变高产GABA菌株并驯化其发酵特定底物,开发谷物适用的直投式发酵剂,将为谷物发酵食品的发展带来新的可能。
[1]Gao Y,Heldt S A. Gamma-Aminobutyric Acid[J]. Phobias:The Psychology of Irrational Fear:The Psychology of Irrational Fear,2015:189.
[2]Meyerhoff D J,Mon A,Metzler T,et al. Cortical gamma-aminobutyric acid and glutamate in posttraumatic stress disorder and their relationships to self-reported sleep quality[J]. Sleep,2014,37(5).
[3]Maric T T,Piantadosi P T,Floresco S B. Prefrontal cortical gamma-aminobutyric acid transmission and cognitive function:drawing links to schizophrenia from preclinical research[J]. Biological psychiatry,2015,77(11):929-939.
[4]Brady R O,McCarthy J M,Prescot A P,et al. Brain gamma-aminobutyric acid(GABA)abnormalities in bipolar disorder[J]. Bipolar disorders,2013,15(4):434-439.
[5]Yoto A,Murao S,Motoki M,et al. Oral intake ofγ-aminobutyric acid affects mood and activities of central nervous system during stressed condition induced by mental tasks[J]. Amino acids,2012,43(3):1331-1337.
[6]Hayes D J,Jupp B,Sawiak S J,et al. Brainγ-aminobutyric acid:a neglected role in impulsivity[J]. European Journal of Neuroscience,2014,39(11):1921-1932.
[7]Xie Z,Xia S,Le G W. Gamma-aminobutyric acid improves oxidative stress and function of the thyroid in high-fat diet fed mice[J]. Journal of Functional Foods,2014,8:76-86.
[8]孙利净,胡满.γ-氨基丁酸及其受体生理功能与作用机制的研究进展[J]. 黑龙江畜牧兽医,2016,1:019.
[9]吕莹果,张晖,王立,等. 植物中γ-氨基丁酸的代谢和功能[J]. 中国食品添加剂,2010(1):92-99.
[10]Kihara M,Okada Y,Iimure T,et al. Accumulation and Degradation of Two Functional Constituents,GABA and. BETA. -Glucan,and Their Varietal Differences in Germinated Barley Grains[J]. Breeding Science,2007,57(2):85-89.
[11]Jin W J,Kim M J,Kim K S. Utilization of Barley or Wheat Bran to Bioconvert Glutamate toγ-Aminobutyric Acid(GABA)[J]. Journal of food science,2013,78(9):C1376-C1382.
[12]Cha M N,Jun H I,Song G S,et al. The effects of germination conditions on GABA and the nutritional components of barley[J]. Korean Journal of Food Science and Technology,2012,44(1):41-47.
[13]Chung H J,Jang S H,Cho H Y,et al. Effects of steeping and anaerobic treatment on GABA(γ-aminobutyric acid)content in germinated waxy hull-less barley[J]. LWT-Food Science and Technology,2009,42(10):1712-1716.
[14]Kleinwächter M,Meyer A K,Selmar D. Malting revisited:Germination of barley(HordeumvulgareL.)is inhibited by both oxygen deficiency and high carbon dioxide concentrations[J]. Food chemistry,2012,132(1):476-481.
[15]朱云辉,郭元新. 发芽苦荞 GABA 的富集及氨基酸等含氮物的变化[J]. 食品与发酵工业,2015,41(2):85-88.
[16]Wu F,Yang N,Touré A,et al. Germinated brown rice and its role in human health[J]. Critical reviews in food science and nutrition,2013,53(5):451-463.
[17]Karladee D,Suriyong S.γ-Aminobutyric acid(GABA)content in different varieties of brown rice during germination[J]. Science Asia,2012,38(13):13-17.
[18]Kim H Y,Hwang I G,Kim T M,et al. Chemical and functional components in different parts of rough rice(OryzasativaL.)before and after germination[J]. Food Chemistry,2012,134(1):288-293.
[19]Liu R,He X,Shi J,et al. The effect of electrolyzed water on decontamination,germination andγ-aminobutyric acid accumulation of brown rice[J]. Food control,2013,33(1):1-5.
[20]Zhang Q,Xiang J,Zhang L,et al. Optimizing soaking and germination conditions to improve gamma-aminobutyric acid content in japonica and indica germinated brown rice[J]. Journal of Functional Foods,2014,10:283-291.
[21]Chungcharoen T,Prachayawarakorn S,Tungtrakul P,et al. Effects of germination process and drying temperature on gamma-aminobutyric acid(GABA)and starch digestibility of germinated brown rice[J]. Drying Technology,2014,32(6):742-753.
[22]Ng L T,Huang S H,Chen Y T,et al. Changes of tocopherols,tocotrienols,γ-oryzanol,andγ-aminobutyric acid levels in the germinated brown rice of pigmented and nonpigmented cultivars[J]. Journal of agricultural and food chemistry,2013,61(51):12604-12611.
[23]王仁辰. 利用米胚芽中麸胺酸脫羧酶与味精反应以提升γ-胺基丁酸(GABA)生成量之探讨[J]. 中兴大学食品暨应用生物科技学系所学位论文,2013:1-90.
[24]Zhang H,Yao H Y,Chen F. Accumulation ofγ-aminobutyric acid in rice germ using protease[J]. Bioscience,biotechnology,and biochemistry,2006,70(5):1160-1165.
[25]Saikusa T,Okada T,Murai H,et al. The effect of defatting with organic solvent on accumulation of 4-aminobutyric acid(GABA)in the rice germ[J]. Journal-Japanese society of food science and technology,2001,48(3):196-201.
[26]Sharma N,Niranjan K. Foxtail millet:properties,processing,health benefits and uses[J]. Food Reviews International,2017(just-accepted).
[27]Sharma S,Saxena D C,Riar C S. Analysing the effect of germination on phenolics,dietary fibres,minerals andγ-amino butyric acid contents of barnyard millet(Echinochloafrumentaceae)[J]. Food Bioscience,2016,13:60-68.
[28]Sharma S,Saxena D C,Riar C S. Using combined optimization,GC-MS and analytical technique to analyze the germination effect on phenolics,dietary fibers,minerals and GABA contents of Kodo millet(Paspalum scrobiculatum)[J]. Food Chemistry,2017.
[29]Bai Q,Chai M,Gu Z,et al. Effects of components in culture medium on glutamate decarboxylase activity andγ-aminobutyric acid accumulation in foxtail millet(SetariaitalicL.)during germination[J]. Food Chemistry,2009,116(1):152-157.
[30]Bai Q,Yang R,Zhang L,et al. Salt stress induces accumulation of gamma-aminobutyric acid in germinated foxtail millet(SetariaitalicaL.)[J]. Cereal Chem,2013,90(2):145-149.
[31]Paucar-Menacho L M,Martínez-Villaluenga C,Dueas M,et al. Optimization of germination time and temperature to maximize the content of bioactive compounds and the antioxidant activity of purple corn(ZeamaysL.)by response surface methodology[J]. LWT-Food Science and Technology,2017,76:236-244.
[32]Polthum P,Ahromrit A. GABA content and Antioxidant activity of Thai waxy corn seeds germinated by hypoxia method[J]. Songklanakarin Journal of Science & Technology,2014,36(3).
[33]王锦茹,赵红梅,胡艳宇,等. Saccharomyces cerevisiae 谷氨酸脱羧酶的诱导纯化及其活性的研究[J]. 黑龙江畜牧兽医,2016(6):33-36.
[34]Kleinwächter M,Müller C,Methner F J,et al. Biochemical heterogeneity of malt is caused by both biological variation and differences in processing:I. Individual grain analyses of biochemical parameters in differently steeped barley(HordeumvulgareL.)malts[J]. Food chemistry,2014,147:25-33.
[35]金红星,田楠,成文玉,等. 微生物发酵合成γ-氨基丁酸的研究进展[J]. 安徽农业科学,2012,40(11):6385-6386.
[36]Cai S,Gao F,Zhang X,et al. Evaluation ofγ-aminobutyric acid,phytate and antioxidant activity of tempeh-like fermented oats(AvenasativaL.)prepared with different filamentous fungi[J]. Journal of Food Science and Technology,2014,51(10):2544-2551.
[37]路锶,田北,赵丹,等.真菌固态发酵对荞麦营养功能成分的影响[J].食品科技,2012(8):148-151.
[38]董玲,姚英政,朱宇. 乳酸菌发酵糙米产γ-氨基丁酸的研究[J]. 食品与发酵科技,2014,50(5):9-11.
[39]董玲,姚英政,朱宇. 短乳杆菌发酵提高糙米中γ-氨基丁酸含量的研究[J]. 粮食与饲料工业,2016(3):19-22.
[40]杨斯茸,魏淑珍,李苏红,等. 糙米酒发酵特征参数的研究[J]. 粮油食品科技,2013,21(2):62-64.
[41]杨丽丽,赵城彬,吴非. 乳酸菌发酵米糠产γ-氨基丁酸最适条件的研究[J]. 食品工业科技,2012,33(16):217-220.
[42]王姣斐. 压差式膨化前处理米糠及其发酵生产γ-氨基丁酸的研究[D]. 天津:天津科技大学,2014.
[43]姚世聪,赵丹,路锶. 不同发酵剂对小米营养成分及γ-氨基丁酸含量的影响[J]. 食品工业科技,2012,33(19):165-167.
[44]蒋冬花,高爱同,毕珂,等. 乳酸菌发酵小米糠生产γ-氨基丁酸的配方和条件优化[J]. 浙江师范大学学报:自然科学版,2013,36(1):6-10.
[45]张丽芳,翟玮玮,曹阳. 一株保加利亚乳杆菌富集米胚芽γ-氨基丁酸发酵条件优化研究[J]. 食品科技,2013,38(8):14-19.
[46]李婷,任欢,满朝新,等. 植物乳杆菌发酵玉米低聚肽乳饮料的研制[J]. 中国乳品工业,2015,43(9):51-53.
[47]康彬彬.发芽糙米的制备及其乳酸发酵工艺的研究[D].福州:福建农业大学,2006.
[48]魏一星,董英,周兴华,等. 乳酸菌发酵改善小麦胚芽的营养品质与贮藏特性的研究[J]. 现代食品科技,2014,30(4):147-153.
[49]Tamaruay K,Intaket R,Kaewkumsan P. The effect of fermentation of germinated brown rice and KKU URL0381 cultivar black rice on GABA content and chemical composition of by-product and fermented rice juice[J]. Food and Applied Bioscience Journal,2015,3(1):21-29.
[50]陈璐,祁勇刚,王常苏,等. 多菌混合发酵红曲生产 Monacolin K 与 GABA 的研究[J]. 中国酿造,2012,31(11):124-128.
[51]Coda R,Lanera A,Trani A,et al. Yogurt-like beverages made of a mixture of cereals,soy and grape must:microbiology,texture,nutritional and sensory properties[J]. International Journal of Food Microbiology,2012,155(3):120-127.
