啮齿动物对不同大小辽东栎和山桃种子的取食和搬运
2018-01-17欧瑞东闫兴富马惠娟姜英娜
欧瑞东,闫兴富,马惠娟,姜英娜
(北方民族大学生物科学与工程学院国家民委生态系统模型及应用重点实验室,宁夏 银川750021)
啮齿动物对种子的取食和搬运作为植物的一种有效扩散机制[1],将成年植株生活史过程的种子阶段和幼苗建成联系在一起,是影响植物种群适合度和动态的重要生态学过程,对植物种群更新、群落物种组成和多样性都具有重要影响[2]。食物贮藏是动物长期适应于不确定的食物供应变化的重要行为策略,植物种子通过啮齿动物的搬运和贮藏而实现的扩散不仅可降低种子被其他动物发现和取食的机会[3],有利于种子定向扩散至更有利于萌发和幼苗建立的微生境,而且大大降低同种个体间的竞争和幼苗密度依赖性死亡[3]。因此,扩散动物对种子的捕食、搬运和贮藏通过种子存活和幼苗建立而影响植物的种群更新和分布[4]。
种子特征对啮齿动物的取食和搬运行为偏好具有重要影响[57],其中种子大小是种子质量的直接反映,比其他特征更易于作为评价种子质量的指标[5],是决定种子是否及如何被动物扩散和扩散距离的最重要而持久的因素[89]。大种子通常具有更高的营养价值,具有单位时间和能量投入的更高营养回报和潜在收益[10],被啮齿动物贮藏的可能性更大,而且大而营养丰富的种子被扩散的距离更远,而小种子则更为可能被啮齿动物取食[8,11]。一些种子的厚而坚硬的种皮对动物的觅食行为策略具有调节作用[12],坚硬种皮引起的种子处理时间的延长意味着动物吃掉种子要比贮藏种子耗费的时间更长。因此,动物为了提高单位时间投入的觅食净收益,可能优先就地取食种皮薄而脆的种子,而搬运和贮藏种皮坚硬的种子[13]。
辽东栎(Quercus wutaishanica)是我国暖温带落叶林的重要优势树种之一,种子在萌发前后被动物大量取食可能是限制其种群更新的重要瓶颈因素[1415];山桃(Prunus davidiana)是在六盘山区散生于天然次生林中的主要灌木树种,局部地区也有以山桃为优势的灌丛群落,有关山桃的研究主要集中于破除种子休眠的方法[16]、叶片光合与蒸腾生理特征及气体交换参数[17]和林地水分动态及耗水性能[18]等方面,其种子成熟后是否与辽东栎种子一样被啮齿动物取食和扩散还缺乏研究。因此,为了比较啮齿动物对这两类大小和种皮特征差异较大的种子(辽东栎种子的种皮薄而脆,山桃种子的种皮厚而坚硬)取食和搬运行为,本试验采用塑料标签标记法,研究了啮齿动物对不同大小的辽东栎和山桃种子的取食和搬运行为及其对种子命运的影响,研究结果可为这2个物种的人工辅助种群更新和直播造林及苗木繁育等林业生产实践提供参考。
1 研究地区和样地概况
研究地区为六盘山国家级自然保护区(35°15′~35°41′N,106°09′~106°30′E),属温带半湿润大陆性季风气候区,冬季寒冷干燥,夏季高温多雨,年降水量676 mm,其中60%的降水集中于7、8、9月,年蒸发量1 426 mm,年均温5.8℃,土壤以灰褐土为主[19]。样地位于保护区的龙潭林场所属的大倒沟林区(106°21.12′E,35°23.54′N,海拔高度1 900 m)的华北落叶松(Larix principisrupprechtii)人工林,林地位于西向山坡,坡度约35°,该林地为20世纪80年代初在辽东栎次生林皆伐后栽植,呈带状分布,林带宽约20 m,冠层盖度约70%,林下有辽东栎、珍珠梅(Sorbaria sorbifolia)、华西箭竹(Fargesia nitida)、南川绣线菊(Spiraea rosthornii)、阔叶荚!(Viburnum lobophyllum)和冰草(Agropyron cristatum)等灌木和草本植物,群落物种组成和研究地区的啮齿动物种类详见相关文献[1920]。
2 材料与方法
2.1 种子的采集和标记
辽东栎种子采自六盘山国家级自然保护区内的辽东栎植株冠层下自然散落的种子,采集时将大、小种子分开,采集表面未见明显破损和昆虫取食危害的种子,自然风干1周后大、小种子质量分别为(1.6±0.3)g(n=100)和(1.0±0.2)g(n=100);山桃种子为2013年秋季购自当地农民当年采摘的种子,大、小种子质量分别为(3.1±0.5)g(n=100)和(1.3±0.2)g(n=100),经自然风干后于室内常温贮藏。种子标记参照常罡等[21]的方法(略有改进),用钻头直径为0.5 mm 的电钻在种子基部或一侧钻一小孔,分别用长约7.5 cm(直径为0.4 mm,外面包有塑料保护层)的铜线将种子与大小为2.5 cm×1.2 cm 的塑料标签(含铜线质量(0.17±0.002)g,n=100)连接,种子的标签信息包括种子投放点及种子编号等。
2.2 种子投放和野外调查
2014年4月6日,选取4条华北落叶松人工林带作为试验样带,分别投放辽东栎大种子(LQW)、辽东栎小种子(SQW)、山桃大种子(LPD)和山桃小种子(SPD);每一样带沿山坡自下而上间隔20 m重复设置4个种子投放点,每一投放点分别投放50枚种子,将标记种子的标签朝外排列成一直径约20 cm的圆环状。共计投放种子4样带×4投放点(重复)×50枚=800枚。分别在种子投放后的第2、4、7、21天和第35天调查记录被动物就地取食的种子数量,在距离投放点25 m直径范围内搜寻种子被啮齿动物搬运后(取食、丢弃于地表和贮藏)丢弃的标签,统计不同类型命运种子的数量,记录种子标签信息,并测定标签到对应种子投放点的距离(m)。
2.3 数据处理
根据野外调查记录的各种子投放点留存种子的数量,计算各调查时期不同类型种子的留存种子数占最初投放种子数的百分比(即留存率);将种子命运分为3种类型:1)就地取食(insitupredation):种子在投放点被动物取食;2)搬运后取食(predation after removal):种子被动物搬运后取食;3)搬运后贮藏(hoarding after removal):种子被动物搬运后埋藏于土壤、洞穴中或弃于地表(因贮藏于洞穴中和丢弃于地表的种子较少,与埋藏于土壤中的种子合并统计)。统计不同命运种子的百分比、种子被搬运的距离及其分布频次。将所有试验数据进行平方根换后,用单因素方差分析(oneway ANOVA)的方法分析不同命运种子的百分比在不同类型种子间的差异显著性,所有数据统计均在SPSS 13.0中完成。
3 结果与分析
3.1 种子留存率及动态
从图1可以看出,辽东栎种子在投放后被啮齿动物取食和搬运的速率较快,种子投放2 d后,辽东栎大种子和小种子的留存率分别为29.5%和22.5%,随着种子投放时间的延长,留存率快速降低,种子投放15 d后,留存率分别降至3.0%和0.5%,种子投放21 d后留存率均为0;相反,山桃种子的留存率随着投放时间的延长降低较为缓慢,种子投放2 d后留存率分别高达95.5%和85.5%,在投放15 d后仍相对较高(分别为21.0%和32.0%),试验结束时仍有5.5%和3.0%的种子留存于种子投放点。方差分析结果显示,不同大小辽东栎种子的留存率仅在投放4 d后差异显著(p<0.05),但二者在投放后所有观测时期与LPD间的差异均达显著水平(p<0.01),LQW 与SPD、SQW与LPD间除在投放35 d后差异不显著外,其他各观测时期均差异显著(p<0.01),不同大小山桃种子的留存率在投放早期(投放2 d和7 d后)差异显著(p<0.015)。
3.2 不同类型种子的命运
从图2可以看出,LQW、SQW、LPD和SPD的就地取食率分别为9.5%、39.5%、5.5%和9.0%,其中SQW的就地取食率均显著高于其他类型种子(p<0.01)。LQW 被啮齿动物搬运后的取食率最大(33.5%),SQW 次之(21.0%),其中前者显著大于后者和不同大小的山桃种子(p<0.05);SQW被动物搬运后的取食率还显著大于SPD(8.5%,p<0.01)。不同大小的山桃种子被啮齿动物搬运后的贮藏率分别为38.0%和47.0%,差异不显著,但SPD均显著大于不同大小的辽东栎种子(p<0.05),SQW被啮齿动物搬运后的贮藏率最小(15.0%),均显著低于其他3种类型的种子(p<0.05)。
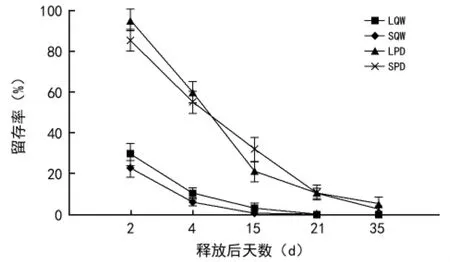
图1 不同大小的辽东栎和山桃种子在六盘山区华北落叶松人工林释放点的留存率及动态
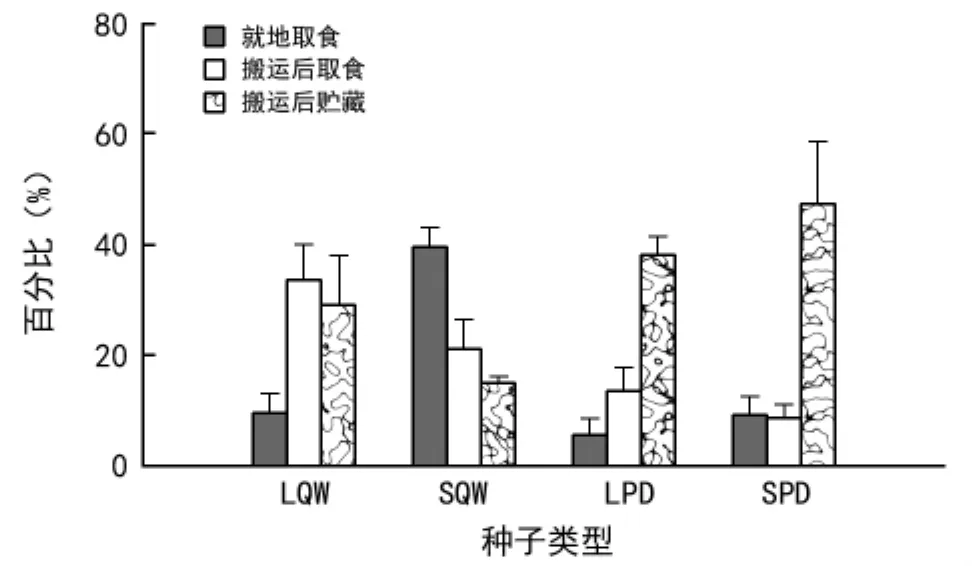
图2 不同大小的辽东栎和山桃种子在六盘山区华北落叶松人工林释放后的命运
3.3 不同类型种子被啮齿动物搬运的距离
从图3可以看出,LPD被啮齿动物搬运后取食的距离最大为4.1 m,其次为LQW(1.9 m),SQW 被啮齿动物搬运后取食的距离最小(0.8 m),除LQW与SPD间差异不显著外,其他类型种子间均差异显著(p<0.05)。种子被啮齿动物搬运后贮藏的距离也表现为LPD>LQW>SPD>SQW,分别为3.6,2.0,1.5 m 和1.4 m,各类型种子间的差异均达显著水平(p<0.05)。
3.4 种子搬运距离的分布频次
从图4可以看出,LQW被啮齿动物搬运距离的分布频次在1~2 m距离组最高(43.7%),2~5 m距离组次之(38.7%),<1 m 距离组较低(16.8%),>5 m距离组仅0.8%;超过1/2(56.3%)的SQW 被啮齿动物搬运的距离分布于<1 m距离组,其次为1~2 m距离组,2~5 m和>5 m距离组的分布频次均为0。LPD被啮齿动物搬运的距离集中分布于2~5 m距离组(83.8%),1~2 m和>5 m 距离组的分布频次相等(6.3%),<1 m距离组最低;SPD被啮齿动物搬运距离的分布频次在1~2 m距离组最高(44.8%),2~5 m距离组次之(32.4%),>5 m 距离组为0。
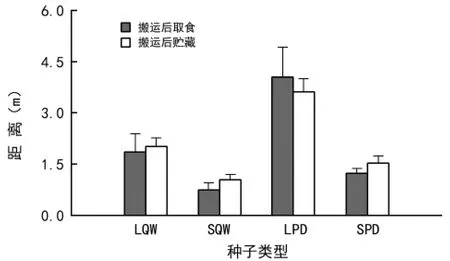
图3 不同大小的辽东栎和山桃种子在六盘山区华北落叶松人工林被啮齿动物搬运后取食和贮藏的距离
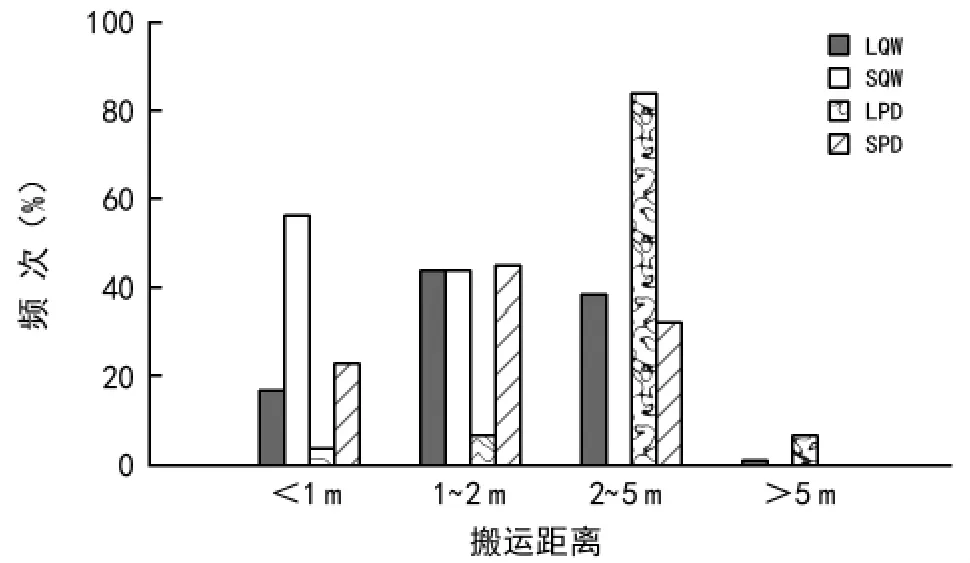
图4 不同大小的辽东栎和山桃种子在六盘山区华北落叶松人工林被啮齿动物搬运距离的分布频次
4 讨 论
啮齿动物对不同植物种子的取食和扩散行为偏好可能对种子的大小、种皮厚度、防御性成分含量及营养价值等特征做出回应[5,7,22],其中种子大小是种子质量的直接反映[5],啮齿动物对种子搬运和贮藏过程中的能量投入与种子大小呈正相关关系[8],大种子具有单位时间和能量投入的更高营养回报和潜在收益[10]。然而,本研究中,啮齿动物并没有优先取食或搬运较大的种子,尤其在种子投放的早期阶段,LQW和LPD的留存率都不同程度地高于各自的小种子。这一结果表明,尽管取食和搬运大种子获得的营养回报更高,但不同大小种子间种皮厚度的细微差异也会影响动物处理种子的时间,从而影响动物对自身被捕食风险的判断,啮齿动物可能具有区分这些细微差异的能力,尤其在春季群落冠层缺乏叶片遮蔽的条件下,动物尽量缩短觅食过程中的时间投入以降低觅食期间自身被捕食风险,表明在觅食效益和自身被捕食风险之间,动物优先选择降低被捕食的风险,从而以牺牲一定的觅食效益为代价[13]。
植物种子的机械障碍(如坚硬的种皮)可通过延长动物去除种皮的时间投入而对动物的觅食策略具有一定的调节作用[12],动物去除种皮时间的延长不仅会增大自身被捕食的风险,降低动物单位时间投入的净收益[23],而且还会减少其从事其他活动(休息或繁殖等)的时间投入。研究发现,辽东栎种子在投放后被啮齿动物取食和搬运的速率较快,种子投放2 d后,LQW和SQW 的留存率仅有29.5%和22.5%,而LPD和SPD的留存率仍分别高达95.5%和85.5%,且辽东栎种子的留存率随着种子投放时间的延长,快速降低至0,但直至试验结束不同大小的山桃种子都有少量留存于种子投放点。这些结果表明,辽东栎和山桃种子的种皮结构差异可能是影响啮齿动物觅食决策的重要因素,山桃种子的厚而坚硬的种皮引起的觅食效益的降低和动物自身被捕食风险的提高对啮齿动物优先取食和搬运辽东栎种子具有决定性作用,这是啮齿动物对觅食效益和自身被捕食风险进行权衡的结果[1314,23]。
动物在最初遇到种子时,面临立即吃掉种子还是暂时贮藏而在未来某一时间再吃掉种子的选择[12]。为了补偿因坚硬种皮引起的种子处理时间延长可能带来的风险损失,提高单位时间投入的觅食净收益,啮齿动物可能选择就地取食种皮薄而脆的种子,而将种皮坚硬的种子搬运后取食或贮藏[13];大种子通常具有更高的营养价值,啮齿动物倾向于选择就地取食小种子,而将大种子搬离资源点后取食或贮藏起来[8,13]。本研究中,SQW被啮齿动物的就地取食率和LQW被啮齿动物搬运后的取食率均显著高于其他类型的种子,而不同大小的山桃种子被搬运后的贮藏率均高于辽东栎种子,SQW被啮齿动物搬运后的贮藏率最低。这些结果表明,啮齿动物倾向于选择就地取食种皮薄而脆的辽东栎种子,而将种皮坚硬的种子搬运后取食或贮藏[13,23],这一觅食策略既可提高单位时间投入的净营养回报,又能利用种皮坚硬的山桃种子耐受长期贮藏的优势。动物在遇到种皮特征相同的种子时,可能将大小作为判断种子质量的主要依据[13],尤其在春季食物供应不确定的情况下,动物往往选择将较大的种子贮藏后再取食,这一相对保守的行为策略要优越于不贮藏食物的冒险策略[24]。因此,啮齿动物的觅食行为决策可能受种子特征的综合影响,因为任何单一的种子性状都不能决定种子的扩散模式,单个种子的收益是影响种子命运的关键因素[25]。
根据最优贮藏空间模型(Optimal Cache Spacing Mode)预测,动物可能在尽量大的空间分散贮藏食物以降低贮藏点密度,从而降低贮藏食物被盗取食物的竞争者发现和盗取的风险。具有更高营养价值的大而种皮较薄的种子被动物贮藏于远离食物资源点的区域,而营养价值相对较低的小而种皮较厚的种子则被动物贮藏于食物资源附近的区域[89]。本研究发现,山桃大种子被啮齿动物搬运后取食和贮藏的距离均最大,分别为4.1 m和3.6 m,而且搬运后的取食和贮藏距离都表现为山桃大种子>辽东栎大种子>山桃小种子>辽东栎小种子。这可能是啮齿动物对不同类型种子大小、种皮结构等特征的综合反应,也是啮齿动物对觅食决策效益和觅食过程中的自身被捕食风险进行权衡的结果。一方面,对种皮厚而坚硬的山桃种子来说,坚硬种皮造成的种子处理时间延长可能使动物自身被捕食风险的提高,为了补偿因这一风险损失,动物可能采取牺牲部分营养收益的行为策略,从而选择将种皮坚硬的种子搬运至更远的距离取食或贮藏[13];另一方面,啮齿动物为了使营养回报最大化,往往通过多次搬运而将较大的种子搬运至更远的距离取食或贮藏,这也是一种降低贮藏食物被食物竞争者盗取风险的行为策略[26],尽管这一策略可能会增加觅食过程中的能量和时间投入,但这一行为不仅可降低被食物竞争者盗取的风险[27],而且有利于啮齿动物对食物贮藏点进行检查、管理,增强动物对贮藏点状态和位置信息的记忆,并对种子贮藏条件进行优化,以降低贮藏食物的损失[28]。因此,LPD和LQW被啮齿动物搬运的距离均大于其各自的小种子,而且LQW的搬运距离还大于大小与其基本一致的SPD。
[1]Hirsch BT,Kays R,Pereira VE,et al.Directed seed dispersal towards areas with low conspecific tree density by a scatterhoarding rodent[J].Ecology Letter,2012,15:1 4231 429.
[2]Meng LZ,Gao XX,Chen J,et al.Spatial and temporal effects on seed dispersal and seed predation ofMusa acuminatain southern Yunnan,China[J].Integrative Zoology,2012,7:3040.
[3]Ruxton GD,Schaefer H M.The conservation physiology of seed dispersal[J].Philosophical Transactions of the Royal Society B,2012,367:1 7081 718.
[4]梁振玲,马建章,戎可.动物分散贮食行为对植物种群更新的影响[J].生态学报,2016,36(4):1 1621 169.
[5]Wang B,Chen J,Corlett RT.Factors influencing repeated seed movements by scatterhoarding rodents in an alpine forest[J].Scientific Reprts,2014,4:4 786.
[6]Xiao ZS,Gao X,Zhang ZB.Sensitivity to seed germination schedule by scatterhoarding Pére David’s rock squirrels during mast and nonmast years[J].Ethology,2013,119:472479.
[7]Yi XF,Zhang MM,Bartlow AW,et al.Incorporating cache management behavior into seed dispersal:The effect of pericarp removal on acorn germination[J].PLoS ONE,2014,9(3):92 544.
[8]Wang B,Wang G,Chen J.Scatterhoarding rodents use different foraging strategies for seeds from different plant species[J].Plant Ecology,2012,213:1 3291 336.
[9]Wang B,Chen J.Seed size,more than nutrient or tannin content,affects seed caching behavior of a common genus of Old World rodents[J].Ecology,2009,90:3 0233 032.
[10]曹林,肖治术,张知彬,等.亚热带森区啮齿动物对樱桃种子捕食和搬运的作用格局[J].动物学杂志,2006,41(4):2732.
[11]Wang B,Chen J.Effects of fat and protein levels on foraging preferences of tannin in scatterhoarding rodents[J].PLoS ONE,2012,7(7):40 640.
[12]Vander Wall SB.How plants manipulate the scatterhoarding behavior of seeddispersing animals[J].Philosophical Transactions of the Royal Society B,2010,365:989997.
[13]周立彪,闫兴富,王建礼,等.啮齿动物对不同大小和种皮特征种子的取食和搬运[J].应用生态学报,2013,24(8):2 3252 332.
[14]路纪琪,张知彬.灌丛高度对啮齿动物贮藏和扩散辽东栎坚果的影响[J].动物学报,2005,51(2):195204.
[15]闫兴富,杜茜,石淳,等.六盘山区辽东栎的实生苗更新及其影响因子[J].植物生态学报,2011,35(9):914925.
[16]赵晓光.打破山桃种子休眠方法的研究[J].种子,2005,24(5):62,65.
[17]张淑勇,周泽福,张光灿,等.半干旱黄土丘陵区天然次生灌木山桃(Prunus davidiana)与山杏(Prunus sibiricaL.)叶片气体交换参数日动态差异[J].生态学报,2009,29(1):499507.
[18]程积民,万惠娥,王静.黄土丘陵区山桃灌木林地土壤水分过耗与调控恢复[J].土壤学报,2003,40(5):691696.
[19]闫兴富,余杨春,周立彪,等.啮齿动物对六盘山区辽东栎、野李和华山松种子的取食和搬运[J].生物多样性,2012,20(4):427436.
[20]闫兴富,杨晋,思彬彬,等.凋落物和土壤覆盖对动物取食和搬运辽东栎种子的影响[J].生态学杂志,2011,30(11):2 5472 553.
[21]常罡,肖治术,张知彬.种子大小对小泡巨鼠贮藏行为的影响[J].兽类学报,2008,28(1):3741.
[22]Xiao ZS,Gao X,Zhang ZB.The combined effects of seed perishability and seed size on hoarding decisions by Pére David’s rock squirrels[J].Behavioral Ecology and Sociobiology,2013,67:1 0671 075.
[23]仝磊,路纪琪.黄胸鼠对假海桐和截头石栎种子的贮藏和取食[J].兽类学报,2010,30(3):270277.
[24]Jorge MLSP,Brown JS,van der MM.Handling time and the evolution of caching behavior[J].Behavioral Ecology,2012,23:410417.
[25]于飞,牛可坤,焦广强,等.小型啮齿动物对小兴安岭5种林木种子扩散的影响[J].东北林业大学学报,2011,39(1):1113.
[26]程瑾瑞,张知彬,肖治术.小泡巨鼠在同种竞争者存在下对其贮藏种子的分析[J].兽类学报,2005,25(2):143149.
[27]肖治术,张知彬.啮齿动物的贮藏行为与植物种子的扩散[J].兽类学报,2004,24(1):6170.
[28]Hirsch BT,Kays R,Jansen PA.Evidence for cache surveillance by a scatterhoarding rodent[J].Animal Behaviour,2013,85:1 5111 516.
