白云质砂石山地区翅荚香槐春季光合生理特性研究
2018-01-16苏春花伍生磊
姜 霞,苏春花,伍生磊,张 喜
(1.贵州省林业科学研究院,贵州 贵阳 550005;2.贵州民族大学化学与环境科学学院,贵州 贵阳 550025)
贵州处于东亚岩溶区域中心,是中国碳酸盐岩出露面积最大、岩溶发育最好的省区,同时也是中国土壤侵蚀严重和生态环境脆弱的地区[1]。以贵州高原为中心的中国南方喀斯特地区达5.5×105km2,是全球三大喀斯特集中连片区域中分布面积最大、发育类型最全的地区[2]。贵州省连续性白云岩地区石漠化的面积发生比例仅次于连续性石灰岩地区[3]。白云质砂石山是白云岩组成的砂质丘陵石漠化,其表面剥蚀严重,土层浅薄,为20 cm左右,自然肥力差,土壤经常处于干旱状态[4]。光合作用是树木重要的生理过程,在自然状态下,测定物种的光合作用能更直接反映该植物的光合生理过程[5]。光合作用是植物生态系统中最主要的同化过程,在维持生态系统稳定、平衡大气碳汇中扮演重要角色。它不仅受到植物遗传特性的影响,也与环境因子相关,能反馈植物对环境因子的适应[6]。喀斯特地区雨热同期,旱、雨季区分明显,天气变化频繁,常导致局部高温和干旱,因此光和水也成了限制植物生长的重要因素之一。研究表明,植物对逆境的适应性,从其生长及光合生理生态特性可以直接表现出来,能够很大程度反映出它们对当地生长环境的适应状况[7-8]。对白云质砂石地区植被恢复已成为当务之急,近年来我国对白云质砂石地区研究较少,主要集中在树种生长量[9]、灌木资源调查[10]等方面,对白云质砂石地区造林树种的光合特性还未见报道。翅荚香槐〔Cladrastis platycarpa(Maxim.)Makino〕属于豆科香槐属落叶乔木,是速生优良用材树种,适应范围广。本文以贵州白云质砂石地区适生树种翅荚香槐为研究对象,从植物光合生理特性中探寻植物的生态适应性对策,为白云质砂石地区人工育种造林、苗期管理等提供参考依据,为白云质砂石地区退化生态系统的重建和恢复提供理论指导。
1 材料与方法
1.1 实验地概况
凯里市位于贵州省东南部、苗岭山脉东段北麓,地处26°24′13″~26°48′11″N、107° 41′13″~108°12′19″E,海拔532~1 447 m、平均高度850 m,平均气温13.6~16.2℃,一月均温2.6~5.2℃、七月均温23.2~25.8℃,年日照时数1 289 h、太阳辐射量88.29 kJ/cm2,年降水量1 240.4 mm,年均相对湿度78%,无霜期288 d。属中亚热带温暖湿润季风气候区,植被是中亚热带常绿阔叶林,且属于我国中亚热带东部湿润常绿阔叶林。
试验区位于凯里市东北部小河小流域下游的凯棠乡凯哨村,属清水江流域支流,区内河流切割显著,相对高差较大,为加劳-水寨岩溶槽谷中度-强度石漠化综合治理小区(Ⅲ1)。小流域白云岩分布普遍,石漠化潜在威胁较为严重。造林地海拔550~650 m,坡度10~25°,坡向西南,岩石裸露率低,土层厚度10~30 cm、平均15 cm左右,土壤石砾量高、肥力低,A层0~24 cm,pH7.4~7.8,氮含量2.42~4.16 g/kg,磷含量1.8~4.3 mg/kg,钾含量25.8~90.8 mg/kg,有机质含量4.36%~7.48%,土壤水分2.42%~6.09%[11]。灌木植物盖度为5%~20%,主要为槐蓝(Indigofera tinctora)、细叶铁子(MyrsineSP.)、地瓜藤(Ficus oligodon)等。草本盖度大于90%,主要为拟金茅(Eulaliopsis binata)、牛耳朵(Chirita edurnea)等。小河小流域是典型的白云岩丘陵石漠区,植被恢复困难。
选择树龄12年左右的翅荚香槐3株,树高2.5 m左右,地径3.0 cm左右。选取植株外围上部健康成熟的叶片,采用叶片养分速测仪(TYS-3N)测定植物叶片叶绿素和氮素含量,取平均值作为该叶片的相对叶绿素含量和氮素含量。选取叶片成熟度相近即叶绿素和氮素含量相近的叶片作为待测定叶,各测定植株叶片养分情况见表1。
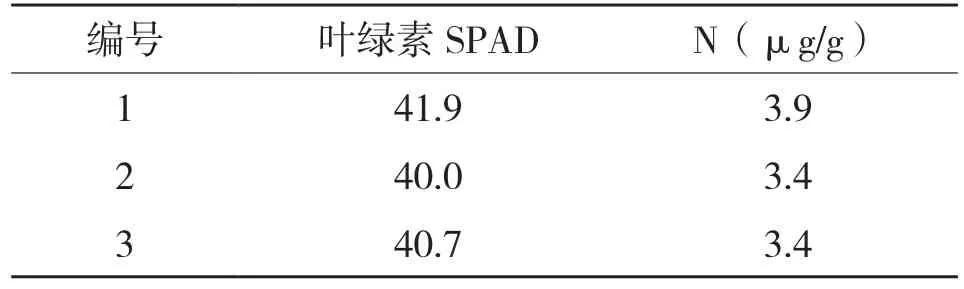
表1 翅荚香槐叶片养分情况
1.2 试验方法
光合日变化试验于2017年5月晴朗少云天气下进行。采用LI-6400xt便携式光合作用测定仪完成各项光合生理指标测定,测定时间段为8:00~19:00,每隔2 h测定光合日变化。每次测定3片样叶,每个样叶3次重复。主要测定参数包括翅荚香槐光合特性因子:净光合速率(Pn,μmol/m2·s)、蒸腾速率(Tr,μmol/ m2·s)、胞间CO2浓度(Ci,μmol/mol)、气孔导度(Gs,mol/m2·s)等;环境因子:光合有效辐射(PAR,μmol/m2·s)、空气相对湿度(RH,%)、空气温度(Ta,℃)、大气CO2浓度(Ca,μmol/m2·s)等。光响应曲线的测定以测定光合日变化的植株为对象。使用LI-6400xt—02B红蓝光源,测定时设定叶片温度25℃,相对湿度70%左右,CO2浓度380 μmol/ mol。设置光强2 000、1 500、1 000、500、250、120、60、30、15、0 μmol/m2·s测量光合生理生态因子的变化,间隔时间3 min,获得一系列数据点,由光合助手软件(Light Response Curve for Photosynthesis)拟合绘制净光合速率与光合有效辐射响应曲线[12]。根据拟合曲线计算出光补偿点、光饱和点、表观量子效率,最大净光合速度、暗呼吸速率等重要参数。
1.3 数据处理
利用LI-6400xt仪器自带软件进行指标数据采集与分析;利用SPSS18.0软件和Excel对野外观测数据进行简单相关分析、方差分析。水分利用效率(WUE)=Pn/Tr;气孔限制值(Ls)=(Ca-Ci)/Ca。
2 结果与分析
2.1 翅荚香槐光合特性日变化
2.1.1 翅荚香槐净光合速率(Pn)与胞间CO2浓度(Ci)日变化 由图1、图2可知,翅荚香槐Pn日变化为双峰曲线,第一峰值出现在上午10:00,为10.15 μmol/m2·s;第二峰值出现在14:00,为6.29 μmol/m2·s,第一峰值高于第二峰值。从早上8:00到10:00 ,随着光合有效辐射的增强,Pn不断增大,到10:00出现第一个峰值,之后出现光抑制现象,Pn开始下降,到中午12:00时降至低谷4.53 μmol/m2·s,随后又逐渐上升,直到下午14:00出现第二个峰值,其后一直到19:00,随着光合有效辐射进一步减弱,Pn逐渐减小。
翅荚香槐Ci则成“V”字型变化,经过夜间的富集,Ci较高,8:00时即达到最大值,为327 μmol/mol,随着植物光合作用的进行,Pn升高,CO2同化加快,Ci降低,在14:00达到一天中最低值,为217 μmol/mol,随后翅荚香槐Pn开始从第2个峰值下降,加之细胞呼吸作用所释放CO2积聚在细胞间隙中,使Ci陡然上升,到19:00时达到全天第二高值,为316 μmol/ mol。总体来看,Pn与Ci变化趋势相反,当叶片Pn升高时Ci降低,而Pn降低时Ci升高。
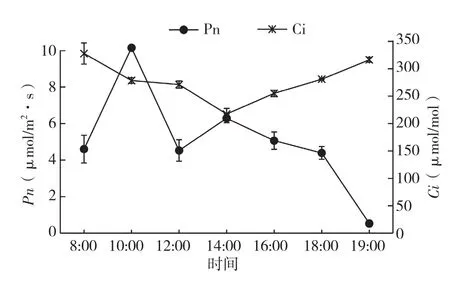
图1 净光合速率与胞间CO2浓度日变化
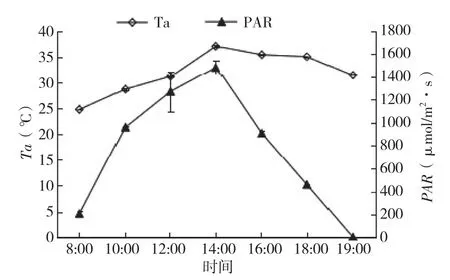
图2 大气温度与光合有效辐射日变化
2.1.2 翅荚香槐净光合速率(Pn)与气孔导度(Gs)日变化 从图3可以看出,翅荚香槐Gs的日变化与净光合速率(Pn)日变化趋势相同,呈双峰曲线。气孔导度日变化最高峰出现在10:00(0.2238 mol/m2·s),与净光合速率第一峰值出现的时间一致,说明气孔导度影响光合作用的气体交换,从而影响光合作用的进行,光合速率的变化与气孔开张程度的变化较一致。其后呈下降趋势,到下午14:00气孔导度降至一个低谷(0.0698 mol/m2·s),随后明显增加,在18:00达到第二高峰值(0.0795 mol/ m2·s)。总体来看,翅荚香槐气孔导度日变化与净光合速率日变化趋势基本一致。光合速率对气孔导度具有指示调节作用,有利于光合时,气孔导度增大,不利时气孔导度减小[13]。图3中翅荚香槐净光合速率日变化与气孔导度日变化呈相近变化趋势,可见光合速率受气孔调节的影响,气孔导度是影响翅荚香槐净光合速率的重要因子。
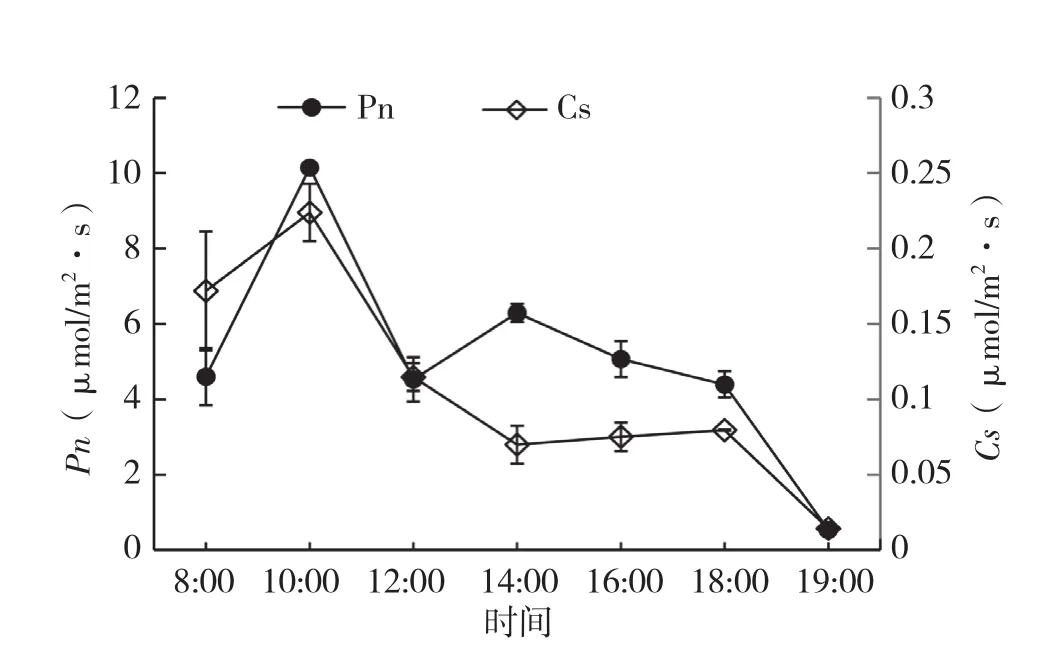
图3 净光合速率与气孔导度日变化
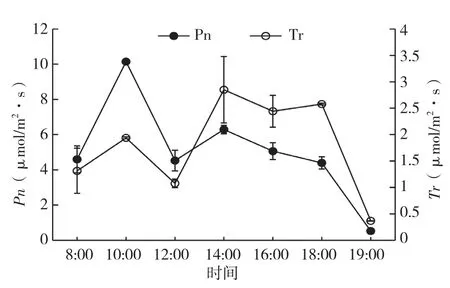
图4 净光合速率与蒸腾速率日变化
2.1.3 翅荚香槐净光合速率(Pn)与蒸腾速率(Tr)日变化 由图4可知,翅荚香槐Tr的日变化与净光合速率(Pn)日变化曲线都呈双峰曲线,清晨和傍晚时蒸腾速率最小,从早上8:00蒸腾速率开始升高,到10:00达到一天中的第一高峰(1.93 mmol/m2·s),随后蒸腾速率不断下降,到12:00达到低谷(1.07 mmol/ m2·s),到14:00时达到第二高峰、为全天最高值(2.85 m mol/m2·s),随后又开始下降。从图2、3、4可看出,从早上8:00开始,翅荚香槐蒸腾速率和净光合速率、气孔导度随着光合有效辐射的增大和气温的升高逐渐上升,蒸腾速率与净光合速率、气孔导度日变化趋势相近,第一峰值出现在早上10:00,随后下降,说明气孔导度是影响蒸腾速率及净光合速率的重要原因之一。
2.1.4 翅荚香槐水分利用效率(WUE)和气孔限制值(Ls)的日变化 从图5可以看出,翅荚香槐Ls的日变化呈双峰曲线,从8:00开始上升,10:00达到第一峰值(0.2546),之后下降,到12:00到达谷值(0.2195),到中午14:00达到一天的最高峰(0.4475),随后又开始下降。而胞间CO2浓度在8:00即达到最大值(327 μmol/mol),随后逐渐降低,在14:00达到一天中的最低值(217 μmol/mol),随后逐渐增加。由此可见,翅荚香槐气孔限制值的日变化与胞间CO2浓度的日变化曲线相反。
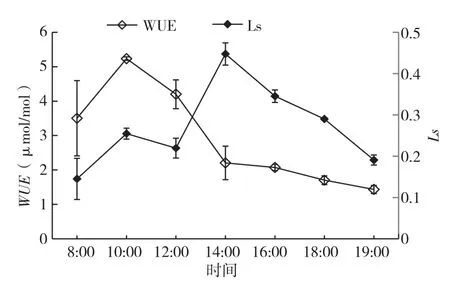
图5 水分利用效率与气孔限制值日变化
从图5可以看出,翅荚香槐水分利用效率呈单峰曲线,日出后,随着光合有效辐射增加,净光合速率增加,虽然蒸腾速率也增加,但光合速率的增幅大于蒸腾速率的增辐,使水分利用效率不断增大,10:00达到一天的峰值,为5.23 μmol/mmol,随后随着光合速率的增幅小于蒸腾速率的增幅,WUE一直下降,到19:00时达到一天的最低值、为1.43 μmol/mol。就日平均值而言,翅荚香槐水分利用效率为2.91 μmol/mol。
2.2 翅荚香槐光响应曲线
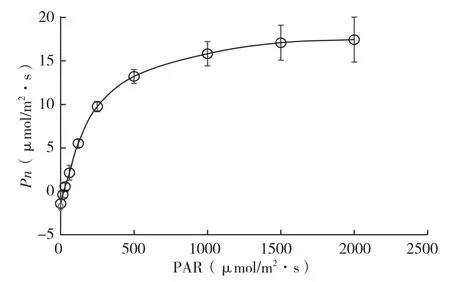
图6 翅荚香槐光响应曲线
光响应曲线反映了植物光合速率随光照强度增减的变化规律[14]。光饱和点和光补偿点是衡量植物光合能力的重要指标。由图6可知,在光合有效辐射低于500 μmol/m2·s时,翅荚香槐的净光合速率呈直线上升,即光合作用诱导期,这一阶段光合作用机构高速运转,光合碳同化酶被充分活化,气孔更加开放[15];当PAR超过500 μmol/m2·s时,净光合速率增长趋于缓和,但净光合速率仍在上升,没有光抑制现象。将3个重复叶片光合有效辐射与净光合速率的数据导入光合助手软件(Light Response Curve for Photosynthesis)直接拟合,求得平均值(表2),得到翅荚香槐的光饱和点(LsP)为984(±169.84)μmol/m2·s、最大净光合速率(Pnmax)为18.46(±2.11)μmol/m2·s、暗呼吸速率(Rd)为-1.39(±0.49)μmol/m2·s。对净光合速率在低光合有效辐射下(<250 μmol/m-2·s)的响应进行直线回归,3个重复直线方程分别为y=0.0441x- 1.2503(R2=0.9769),y=0.0443x- 0.8714(R2=0.9794),y=0.0451x- 0.3199(R2=0.9791),当y=0时,光合过程中吸收的CO2和呼吸过程中放出的CO2等量,此时的光照强度就是光补偿点。光响应曲线的初始斜率即为表观量子效率,求平均值得到表观量子效率(α)为0.0445(±0.00053),光补偿点(LCP)为18.37(±10.68)μmol/m2·s。
2.3 翅荚香槐净光合速率与其生理、环境因子的相关性分析
光合作用是一个复杂的过程,净光合速率的强弱不但与植物自身因素有关,还会受到植物生长环境中各因子的影响[16]。通过将翅荚香槐的净光合速率(Pn)与气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、大气温度(Ta)、大气CO2浓度(Ca)、大气相对湿度(RH)和光合有效辐射(PAR)等指标进行简单相关分析,其相关系数见表2。从表2可以看出,翅荚香槐净光合速率(Pn)与气孔导度(Gs)呈显著正相关,相关系数为0.791;与蒸腾速率(Tr)、大气相对湿度(RH)、光合有效辐射(PAR)呈正相关关系,但相关性不显著;胞间CO2浓度(Ci)、大气温度(Ta)和大气CO2浓度(Ca)呈负相关关系,相关性不显著。各环境因子对翅荚香槐叶片净光合速率的影响大小为Gs(0.791)>PAR(0.575)>Tr(0.552)>Ci(-0.422)>RH(0.405)>Ca(-0.211)>Ta(-0.087)。
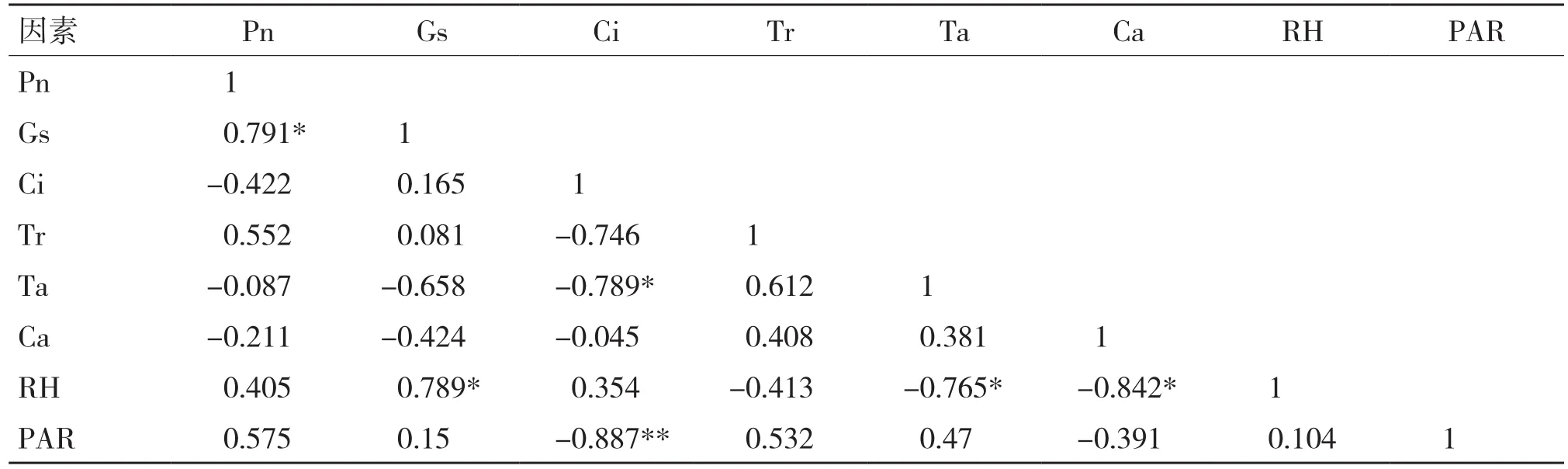
表2 翅荚香槐净光合速率与影响因子相关性分析
3 结论与讨论
植物叶片光合作用的日变化反映了不同植物对环境因子的适应性以及对各种环境因子的利用能力,因植物种类、生长状态和环境因子的不同而异[17]。研究白云质砂石山地区适生树种光合作用日变化的特性,对分析适生树种光合生产和干物质积累具有重要意义。在本研究中,翅荚香槐Pn的日变化为双峰型,在光照和温度高的中午出现明显的光合“午休”现象。这一现象与石栎和青冈[17]在春季为“双峰”曲线相类似。导致植物净光合速率日变化差异的因素主要有环境因子和物种自身生理特性,环境因子主要有光照、大气温度和相对湿度等,自身生理特性包括气孔限制因素和非气孔限制因素。判断叶片光合速率降低的主要原因是气孔因素还是非气孔因素,主要据是Ci和Ls的变化方向,Ci降低和Ls升高表明气孔因素是主要原因,而Ci增高和Ls降低则表明主要原因是非气孔因素[18]。气孔限制因素是因气孔导度下降阻止了CO2的供应;非气孔因素是因叶肉细胞光合能力下降使CO2的同化能力减弱,从而使胞间CO2含量升高。本研究结果显示,白云质砂石山地区翅荚香槐春季净光合速率日变化为双峰曲线,呈“午休”现象,第一峰值出现在上午10:00(10.15 μmol/m2·s),第二峰值出现在14:00(6.29 μmol/m2·s)。从翅荚香槐净光合速率(Pn)与胞间CO2浓度(Ci)和气孔限制值(Ls)日变化的关系可以看出:胞间CO2浓度(Ci)日变化呈“V”字型,即早上和傍晚高,中午低,其变化趋势与净光合速率(Pn)日变化趋势相反,即翅荚香槐净光合速率(Pn)降低时其胞间CO2浓度(Ci)升高;而翅荚香槐气孔限制值(Ls)的日变化与净光合速率(Pn)日变化曲线相同,即当净光合速率(Pn)降低时其气孔限制值(Ls)也降低,说明在白云质砂石山地区翅荚香槐光合“午休”的主要原因是由于非气孔因素造成。同时翅荚香槐光合“午休”也受环境因子的影响,可能是由于试验地为白云质砂石山,其土层较薄,土壤保水性较差,在高温天气下蒸腾作用旺盛,使得翅荚香槐植株缺水,导致气孔部分关闭。本研究中翅荚香槐气孔导度在10:00后明显下降,并一直维持在低值,可能是由外界温度逐渐升高,气孔受高温影响而逐渐闭合造成的。
光补偿点LCP和光饱和点LsP是植物利用光强能力的重要指标[19]。LCP较低、LsP较高的植物对光环境的适应性强,而LCP高、LsP低的植物对光强的适应性较弱。一般LCP在0~10 μmol/m2·s为阴生植物,在10 μmol/m2·s以上为阳生植物[20]。翅荚香槐光补偿点(LCP)为18.37 μmol/m2·s,属于阳生植物;翅荚香槐光补偿点较低,说明其在较低的光强下就开始了有机质的积累,植物的耐荫性强。光饱和点(LsP)较高在984 μmol/m2·s,说明翅荚香槐对强光有较强的适应能力。一般光饱和点较高、光补偿点较低的植物对光环境的适应性较强,高生产,低消耗,生物量积累效率高[21],本研究表明翅荚香槐为高光饱和点、低光补偿点植物,既喜光又耐荫,对环境的适应范围较广。翅荚香槐暗呼吸速率(-1.39 μmol/m2·s)较低,说明其对林下等弱光环境的耐受能力较高;表观量子效率也可以反映物种对弱光的利用能力以及对荫蔽环境的适应能力[22],一般植物的表观量子效率值为0.03~0.06[23],翅荚香槐的表观量子效率(0.0445)较高,说明其对弱光的利用能力以及对荫蔽环境的适应能力较强。
水分利用率的强弱与植物抗旱能力相关,翅荚香槐水分利用效率呈单峰曲线,上午10:00达到一天的峰值(5.23 μmol/mmol),到19:00时达到一天的最低值(1.43 μmol/mmol)。翅荚香槐水分利用效率日平均值较高、为2.91 μmol/ mmol,比喀斯特地区香樟(2.72 μmol/mmol)、法国冬青(2.65 μmol/mmol)[24]高,说明翅荚香槐在白云质砂石山地区更耐干旱。白云质砂石山地区由于其特殊的地质环境,植被恢复困难,在树种的选择时考虑低蒸腾速率、高光合速率和高水分利用效率地树种更利于物质累积,可提高该地区生态效益。
[1]白占国,万国江.贵州碳酸盐岩区域的侵蚀速率及环境效应研究[J].土壤侵蚀与水土保持学报,1998,4(1):1-7,46.
[2]熊康宁,池永宽.中国南方喀斯特生态系统面临的问题及对策[J].生态经济,2015,31(1):23-30.
[3]李瑞玲,王世杰,周德全,等.贵州岩溶地区岩性与土地石漠化的相关分析[J].地理学报,2003,58(2):314-320.
[4]吴智涛,姚银花,龙建华,等.黔东南州白云质砂石山10个树种造林试验初析[J].贵州林业科技,2006,34(4):30-34.
[5]林夏珍,卢婷.遮光对窄头橐吾形态及光合特性的影响[J].浙江林学院学报,2008,25(5):614-618.
[6]姜霞,张喜,谢涛.檫木苗期光合特性及水分利用效率研究[J].贵州林业科技,2016,44(3):19-23.
[7]冯大兰,黄小辉,刘芸,等.4种木本植物在石漠化地区的生长状况及光合特性[J].北京林业大学学报,2015,37(5):62-69.
[8]池永宽,熊康宁,王元素,等.喀斯特石漠化地区肾蕨的光合特性[J].江苏农业科学,2015,43(4):341-344.
[9]吴智涛,姚银花,龙建华,等.黔东南州白云质砂石山10个树种造林试验初析[J].贵州林业科技,2006,34(4):30-34.
[10]杨春玉,杨汉远,袁茂琴,等.喀斯特区白云质砂石山天然次生灌木林植物多样性研究[J].陕西林业科技,2012(5):91-94.
[11]伍铭凯,杨汉远,王定江,等.凯里地区白云质砂石山主要植被类型调查[J].贵州林业科技,2009,37(2):28-31.
[12]张习敏,申刚,陈玲,等.复羽叶栾树光合作用日变化及光响应特征[J].广东农业科学,2015(5):109-114.
[13]姜霞,丁海兵,张喜.苦楝叶片的光合特性研究[J].中国林副特产,2010(5):1-3.
[14]张斌,朱宁华,曹基武.不同生境对赤皮青冈光合特性的影响[J].华南农业大学学报,2017,38(3):70-78.
[15]黄国伟,彭婵,陈慧玲,等.不同种源红椿苗期生长及光合特性比较[J].西北林学院学报,2017,32(2):123-129.
[16]黄桂华,梁坤南,周再知,等.柚木无性系光合作用的日季节变化及其主要影响因素[J].中南林业科技大学学报,2016,36(4):11-16.
[17]管铭,金则新,李月灵,等.千岛湖次生林优势种植物光合生理生态特性[J].生态学报,2015,35(7):2057-2066.
[18]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[19]袁亚琦,王少杰,付宇晨,等.北京地区引种的6种彩叶树种光合生理特性[J].北京农学院学报,2017,32(2):92-97.
[20]Gul,Pallardy S G,Tu K,et al.Reliable estimation of biochemicalPARameters from C3 leaf photosynthesis- inter cellular carbon dioxide response curves[J].Plant,Cell&Environment ,2010,33(11):1852-1874.
[21]金爱武,郑炳松,陶金星,等.雷竹光合速率日变化及其影响因子[J].浙江林学院学报,2000,17(3):271-275.
[22]刘飞虎,梁雪妮,刘小莉.4种野生报春花光合作用特性的比较[J].园艺学报,2004,31(4):482-486.
[23]姚毅,刘艳红,胡耀升,等.珙桐一年生苗光合特性的研究[J].生态科学,2012,31(4):384-389.
[24]姜霞,张喜,丁海兵.黔中10个树种苗期水分利用效率及光合特性研究[J].西部林业科学,2013,42(5):75-81.
