高地钩叶藤与大钩叶藤纤维特性1)
2018-01-15张菲菲李担汪佑宏季必超薛夏王鹏程刘亚梅张利萍
张菲菲 李担 汪佑宏 季必超 薛夏 王鹏程 刘亚梅 张利萍
(安徽农业大学,合肥,230036)
棕榈藤是棕榈科中具刺和鳞状果皮的攀援植物,是亚洲热带地区宝贵的植物资源,也是著名非木材林产品—原藤(仅次于木材和竹材的重要非木材林产品)的来源植物。世界棕榈藤总共有13个属600余种,每种或多或少都有利用价值,但有较高经济价值的只有10~20种[1]。由于棕榈藤具有多种作用,人们还根据不同目的种植不同的藤种。
棕榈藤资源生物利用等正成为我国林业生物产业发展的重点[2-3],棕榈藤的利用主要为原藤利用,其中天然棕榈藤占到90%,而棕榈藤人工林仅占10%。印度尼西亚是棕榈原藤的最大生产国,截止20世纪70年代,印度尼西亚提供了世界原藤的90%。我国95%的商业用藤依靠进口。自2012年1月1日,印度尼西亚出台了更严格的藤原料出口限制政策后,中国藤产业受到了很大冲击。对北京和广东藤产业的调查分析显示,许多小型藤企业已倒闭或转行,剩余的藤企业也面临着严重的原料匮乏问题[4-5]。因此,加大藤资源培育,做好非商业用藤材的性能研究,以期为其改性研究及商业化利用提供基础数据支持。
目前,我国对高地钩叶藤(PlectocomiahimalayanaGriff.)与大钩叶藤(PlectocomiaassamicaGriff.)的研究还处于起步阶段。高地钩叶藤、大钩叶藤属棕榈科(Palmae)省藤亚科(Calamoideae)钩叶藤属(Plectocomia)。高地钩叶藤,属于大径藤(藤径4~5 cm),攀援、丛生藤类。分布于云南南部、西南部、西部海拔1 450~1 800 m的箐沟、竹林、山地常绿阔叶林中,在不丹、印度、尼泊尔、老挝、泰国均有分布。其导管在横切面上近圆形、主要为单管孔、少数为复管孔,导管分子最长,达3.229 mm,导管分子宽度在0.151~0.300 mm。藤茎质地较粗糙,一般用于编织较粗糙的藤器或栅栏用[6]。高地钩叶藤纤维比量为4.9%,径向由外向内,纤维比量呈减小趋势;随着轴向高度的增加,纤维比量呈下降趋势[6]。大钩叶藤分布于云南南部(勐腊)、东南部(马关)、海拔820~1 230 m的次生林中。在印度(阿萨姆、梅加拉亚)、缅甸(北部)也有分布。其藤径质脆,很少利用。藤材导管单管孔,近圆形,导管分子长度在2 mm以上,宽度为0.151~0.300 mm[7]。棕榈藤的功能主要取决于维管束及其周围的维管束鞘[8]。藤茎纤维细胞均存在于纤维鞘中,纤维细胞长,两端尖削,壁厚腔小,其部位不同数量和大小均有差异。藤茎外侧,纤维细胞直径最小,长度大;而中部及中央部纤维细胞直径大,长度短[9]。纤维的性质对藤材的宏观性能有着很大的影响,从而影响到藤材的使用。
1 材料与方法
试验材料高地钩叶藤与大钩叶藤分别采自云南省的梁河县和盈江县。
取样:在藤材基部、距地面2 m、中部和梢部的地方各取一小截1~2 cm长的材料。
永久切片制作[10]:取得的试样经过微波软化后,利用滑走式切片机将藤材横切面切成厚度为40 μm的切片,进行染色、脱色、脱水,透明后用中性树胶固封于载玻片上制成永久性切片。
纤维的分离(富兰克林离析法[11]):用蒸馏水将离析后的样品洗至没有酸味,并用玻璃棒将纤维束轻轻捣散成单根纤维,稀释成浆液备用。
利用光学显微镜和显微图像分析系统(Motic 3.0软件)观察统计处理过的切片与已分离的纤维,在切片上观察纤维在横切面的微观形态,测量纤维的直径和腔径[12-13],离析成单根的纤维用来测量其长度;每个试样测30组重复,取平均值。
2 结果与分析
2.1 高地钩叶藤与大钩叶藤纤维形态径向分布
高地钩叶藤与大钩叶藤的纤维宽度、腔径和双壁厚径向分布见表1和表2。
如表1所示,从藤皮至藤芯,高地钩叶藤纤维的长度依次为1 960.59、2 126.34、1 935.61 μm,呈先上升后下降的趋势,且纤维长度由大到小的顺序为藤中、藤皮、藤芯;大钩叶藤纤维的长度依次为2 144.58、2 048.16、1 855.32 μm,呈逐渐下降的趋势。高地钩叶藤与大钩叶藤纤维长度的平均值分别为2 007.51、2 016.02 μm,前者比后者低0.42%,经F检验(P>0.05)差异不显著(表2)。高地钩叶藤纤维的宽度呈先上升后下降的变化趋势,且由大到小的顺序为藤中、藤芯、藤皮;大钩叶藤纤维的宽度呈先下降后上升的变化趋势,且由大到小的顺序为藤芯、藤皮、藤中。高地钩叶藤与大钩叶藤纤维宽度的平均值分别为18.11、20.46 μm,前者比后者低11.49%,经F检验两种藤材纤维宽度差异不显著(P>0.05)(表2)。高地钩叶藤与大钩叶藤纤维的腔径均为逐渐上升的变化趋势;高地钩叶藤与大钩叶藤纤维腔径的平均值分别为10.62、8.84 μm,前者比后者高20.14%,经F检验两种藤材纤维腔径差异极显著(P<0.01)(表2)。高地钩叶藤纤维的双壁厚呈逐渐下降的变化趋势;大钩叶藤纤维的双壁厚变化趋势为先下降后上升,且纤维厚度由大到小的顺序为藤皮、藤芯、藤中;高地钩叶藤与大钩叶藤纤维双壁厚的平均值分别为7.49、11.61 μm,前者比后者低35.52%,经F检验两种藤材纤维双壁厚差异不显著(P>0.05)(表2)。

表1 高地钩叶藤与大钩叶藤纤维形态的径向分布
注:P.h.为高地钩叶藤(P.himalayana);P.a.为大钩叶藤(P.assamica)。
由表1可看出,高地钩叶藤纤维的长宽比变化于107.36~127.32,最小值和最大值分别在藤中与藤皮处,且藤皮和藤中纤维长宽比差距明显,藤皮至藤芯的下降幅度为20.20%。大钩叶藤纤维的长宽比变化于77.12~113.54,最小值和最大值分别在藤芯与藤皮处,且长宽比由藤皮至藤中下降了2.10%,由藤中至藤芯下降了30.60%。从藤皮到藤芯,高地钩叶藤纤维的腔径比变化于0.39~0.75;大钩叶藤纤维的腔径比变化于0.33~0.50,两者均呈逐渐增大的趋势。高地钩叶藤纤维的壁腔比变化于0.34~1.56;大钩叶藤纤维的壁腔比变化于1.01~2.07,且壁腔比由藤皮至藤中下降了42.60%,由藤中至藤芯下降了15.20%;两者壁腔比均呈逐渐下降的趋势。

表2 高地钩叶藤和大钩叶藤纤维特性径向变异及F检验
注:P.h.为高地钩叶藤(P.himalayana);P.a.为大钩叶藤(P.assamica);P<0.01,差异极显著;0.01≤P<0.05,差异显著;P≥0.05,差异不显著。
2.2 高地钩叶藤与大钩叶藤纤维形态轴向分布
高地钩叶藤与大钩叶藤的纤维宽度、腔径和双壁厚轴向分布见表3和表4。
如表3所示,从基部到梢部,高地钩叶藤纤维的长度依次为2 095.98、2 007.51、2 046.27、1 874.53 μm,呈现起伏状的变化趋势,即由大到小的顺序为基部、中部、距地面2 m处、梢部;大钩叶藤纤维的长度依次为1 963.94、2 016.02、1 959.01、2 123.83 μm,也呈现起伏状的变化趋势,由大到小的顺序为梢部、距地面2 m处、基部、中部,变化与前者相反。经F检验(P<0.01)差异极显著(表4)。高地钩叶藤纤维宽度变化呈先下降后上升的趋势,最小值和最大值分别位于中部和基部;大钩叶藤纤维的宽度整体上呈下降的趋势,最小值和最大值分别位于梢部与距地面2 m处。经F检验(P>0.05)两种藤材纤维宽度差异不显著(表4)。高地钩叶藤纤维腔径变化为起伏式的上升趋势,先由基部上升到距地面2 m处,再下降到中部,后又上升到梢部;大钩叶藤纤维的腔径呈逐渐上升的变化趋势。经F检验(P<0.01)差异极显著(表4)。高地钩叶藤和大钩叶藤纤维双壁厚均呈逐渐下降的变化趋势。经F检验(P>0.05),两者的纤维双壁厚差异不显著(表4)。
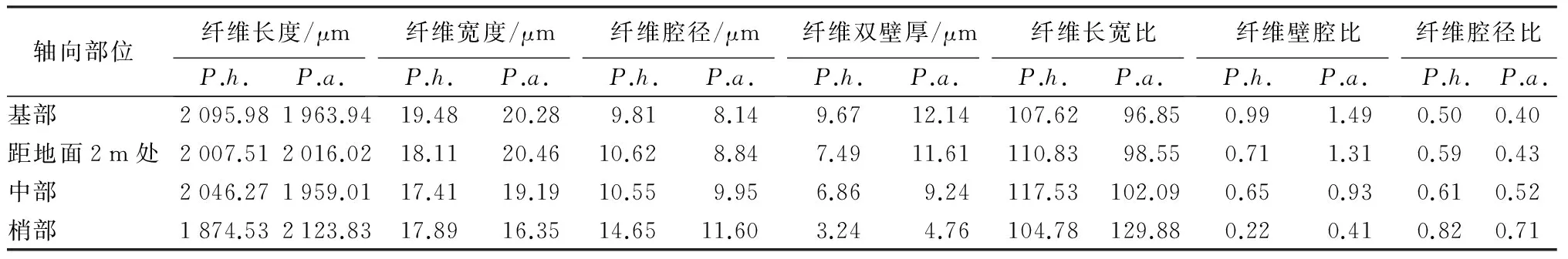
表3 高地钩叶藤与大钩叶藤纤维形态的的轴向变化
注:P.h.为高地钩叶藤(P.himalayana);P.a.为大钩叶藤(P.assamica)。
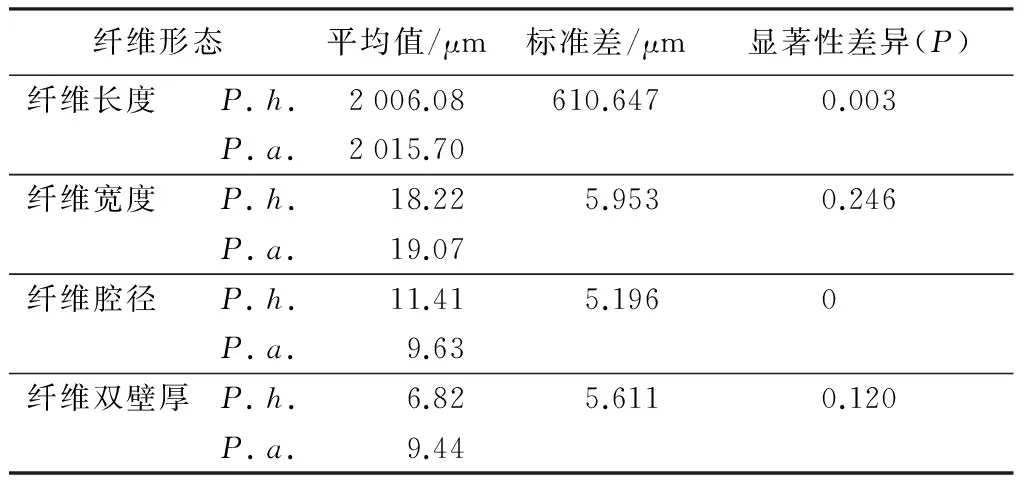
表4 高地钩叶藤和大钩叶藤纤维特性径向变异及F检验
注:P.h.为高地钩叶藤(P.himalayana);P.a.为大钩叶藤(P.assamica);P<0.01,差异极显著;0.01≤P<0.05,差异显著;P≥0.05,差异不显著。
由表3可看出,基部到梢部,高地钩叶藤纤维的长宽比变化于104.78~117.53,从基部的107.62逐渐上升到中部的117.53,后又下降到梢部104.78;大钩叶藤纤维的长宽比变化于96.85~129.88,呈逐渐上升的变化趋势。从基部到梢部,高地钩叶藤纤维的壁腔比变化于0.22~0.99;大钩叶藤纤维的壁腔比变化于0.41~1.49,两者均呈逐渐下降的变化趋势,且高地钩叶藤的壁腔比在各处均比大钩叶藤小。高地钩叶藤纤维的腔径比变化于0.50~0.82;大钩叶藤纤维的腔径比变化于0.40~0.71,两者均呈逐渐上升的趋势,且高地钩叶藤的腔径比在各处均比大钩叶藤大。
3 结束语
从藤皮到藤芯,高地钩叶藤和大钩叶藤纤维的长度变化分别为先增后降和逐渐下降;纤维宽度的变化分别为先增后降和先降后增;纤维的腔径变化两者均为逐渐上升;纤维双壁厚的变化分别为逐渐上升和先降后增。其中高地钩叶藤与大钩叶藤纤维的长度、直径、腔径、双壁厚分别为2007.51 μm和2016.02 μm、18.11 μm和20.46 μm、10.62 μm和8.84 μm、7.49 μm和11.61 μm。高地钩叶藤腔径比大钩叶藤大20.14%,经F检验在0.01的水平上差异极显著;纤维的长度、宽度和双壁厚差异性不显著。两者纤维的长宽比变化分别为先降后增和逐渐下降;纤维的壁腔比变化均为逐渐下降,而纤维的腔径比变化两者均为逐渐上升。
自基部向上,高地钩叶藤和大钩叶藤纤维长度的变化分别为起伏式降-增-降和起伏式增-降-增;纤维宽度和双壁厚的变化两者整体上均呈逐渐下降趋势;纤维腔径的变化两者整体上均呈逐渐上升趋势;经F检验发现,高地钩叶藤与大钩叶藤的纤维长度和腔径轴向变化在0.01的水平上差异极显著;纤维的宽度和双壁厚差异性不显著。高地钩叶藤和大钩叶藤纤维长宽比的变化分别为先增后降和逐渐上升,前者最大值在中部,为117.53,后者最大值在梢部,为129.88,除了梢部,高地钩叶藤的纤维长宽比均比大钩叶藤大;纤维壁腔比的变化两者均为逐渐下降,且前者的壁腔比在各处均比后者小;纤维腔径比的变化两者均为逐渐上升,且前者的腔径比在各处均比后者大。
[1] 江泽慧,萧江华,许煌灿.世界竹藤[M].沈阳:沈阳科学技术出版社,2002:509-612.
[2] 江泽慧.我国林业生物产业发展战略[J].林业经济,2011,33(7):3-5,18.
[3] 江泽慧.中国林产品发展的现状与前景[J].木材工业,2006,20(2):1-4.
[4] 尚莉莉.钩叶藤材的基本性能及增强增韧改性研究[D].北京:中国林业科学研究院,2014.
[5] 尚莉莉,刘杏娥,吕黄飞,等.大钩叶藤与玛瑙省藤材的主要物理力学性质对比[J].东北林业大学学报,2014,42(12):123-125,129.
[6] 刘杏娥,吴明山,汪佑宏,等.高地钩叶藤与钩叶藤组织比量的变异研究[J].安徽农业大学学报,2015,42(1):27-30.
[7] 江泽慧,王慷林.中国棕榈藤[M].北京:科学出版社,2013:93-97.
[8] LIU X E, TIAN G L, SHANG L L, et al. Compression properties of vascular boundles and parenchyma of rattan (PlectocomiaassamicaGriff.)[J]. Holzforschung,2014,68(8):927-932.
[9] 腰希申,许煌灿,曾炳山.棕榈藤的电镜观察Ⅰ藤基部分的电镜观察[J].林业科学,1998,34(3):106-111.
[10] 汪佑宏,徐斌,武恒,等.棕榈藤材解剖特征的取样方法[J].东北林业大学学报,2014,42(10):90-94.
[11] 汪佑宏,徐鑫,王瑞,等.黄藤发育过程中主要解剖特征的变异[J].东北林业大学学报,2010,38(12):35-37,48.
[12] 李坚,陆文达,刘一星,等.体视显微术在木材组织学中的应用[J].东北林业大学学报,1986,14(3):92-98.
[13] 黄广华,陈瑞英,陈居静.红麻秆细胞和纤维形态的研究[J].新余学院学报,2015,20(1):8-12.
