大亚湾海域石斑鱼(Epinephelus spp.)寄生单殖吸虫群落结构研究*
2018-01-12曾碧健黄建荣杨廷宝
曾碧健 黄建荣 杨廷宝①
(1. 广东第二师范学院生物与食品工程学院 应用生态学实验室 广州 510303; 2. 中山大学生命科学学院 广州 510275)
一直以来, 我国关于鱼类寄生虫群落的研究主要集中在淡水鱼类, 而且多数集中在淡水鱼类肠道寄生虫, 关于海水鱼类寄生虫的相对较少(吕军仪等,2001; 吴金英等, 2001; 李文祥等, 2014; Li et al, 2015;卢明淼等, 2015)。近十年来, 我国海水鱼类外寄生虫从最初的形态分类, 逐渐开展了一系列的自然生态学及养殖生态学研究(张剑英, 1999, 2001; Luo et al,2010a, b, 2012)。
石斑鱼作为经济价值极高的海水鱼类一直受到研究者的关注, 国内外关于石斑鱼寄生虫的研究报道, 大部分集中在分类学方面(Moravec et al, 1997;Bu et al, 1999; 张剑英, 2001; Nolan et al, 2004;Justine, 2005a, b, 2007a; Justine et al, 2009), 在石斑鱼寄生虫群落多样性等方面也有报道(Justine, 2007b;Justine et al, 2010)。另外, Leong等(1988)比较了马来西亚野生和网箱养殖的点带石斑(Epinephluls malabaricus)的寄生虫区系; Cruz-Lacierda等(2000)研究了池塘养殖的斜带石斑(E. coioides)寄生石斑分性吸虫(复殖吸虫)的发生和组织病理学。Cribb等(2002)回顾了石斑鱼亚科寄生复殖吸虫的情况。Justine(2007a, b)报道了南太平洋 New Caledonia的花点石斑(E. maculatus)外寄生虫16种, 其中桡足类3种, 等足类幼虫1种, 单殖吸虫12种, 其中鳞盘虫科 10种, 指环虫科 2种。Justine等(2010)对珊瑚鱼28种科鱼类寄生虫多样性进行了7年详细的调查,检获的单殖吸虫主要为鳞盘虫科下四个属, 分别是拟合片虫属已鉴定种33种, 鳞盘虫属4种, 片盘虫属2种, Echinoplectanum属5种, 且Echinoplectanum仅在鳃棘鲈上检获。该调查发现有些石斑鱼类有显著的单殖吸虫多样性, 例如细点石斑(E.cyanopodus), 点带石斑, 花点石斑, 在 New Caledonia采样的鱼类上总计发现鉴定了 42种鳞盘虫科单殖吸虫, Justine等(2010)估计New Caledonia的石斑鱼类很可能有60种拟合片虫, 而在检查的科鱼类中未发现片盘虫, 这提示鳞盘虫科这个成员属的记录可能是偶然的/次要的。
Vidal-Martínez 等(1998)对 尾 斑 丽 体 鱼(Cichlasoma urophthalmus)寄生虫群落结构的构建进行了研究, 发现养殖宿主寄生虫群落模式可预测, 在野生宿主上感染频率最高和平均密度最大的虫种也会首先感染养殖宿主。石斑鱼是东南亚太平洋海岸最重要的鱼类。罗育发等在2008年4月和2009年1月通过季节性采样对中国南海大亚湾的野生和养殖斜带石斑鱼鳃部单殖吸虫群落结构组成和时空变化进行了调查, 结果发现: 斜带石斑鱼鳃部寄生单殖吸虫8种, 分属 3科 4个属: 梅氏新本尼登虫(Neobenedenia melleni), 驼背鲈海盘虫(Haliotrema cromileptis)、石斑鳞盘虫(Diplectanum grouperi)、贾氏拟合片虫(Pseudorhabdosynochus justinei)、南头拟合片虫(P.lantauensis)、斜带拟合片虫(P. coioidesis)和拟合片虫(P. serrani), 以及深圳拟合片虫(P. shenzhensis),这 8种单殖吸虫其感染动态随着季节的环境变化呈现季节变化(Luo et al, 2012)。因此, 对野生宿主寄生虫群落结构的研究结果可为相关种类的养殖防病提供参考。本研究对深圳大亚湾海域常见石斑鱼寄生的单殖吸虫进行了周年调查研究, 旨在分析单殖吸虫的群落结构特点。
1 材料与方法
大亚湾位于广东省东部沿岸, 地理位置介于113°29′42″—114°49′42″E、23°31′12″—24°50′00″N 之间, 被深圳大鹏半岛、惠阳南部沿海及惠东平海半岛三面环绕, 西南邻香港, 南接广阔的南海。
1.1 材料处理及临时形态标本制作
活的石斑鱼样本从市场或渔船购买, 购得的石斑鱼先暂养在充气过滤的玻璃鱼缸, 在48h内处理。为获取体表寄生单殖吸虫, 样本鱼先放入盛咸淡水的大烧杯或塑料盆浸泡约10min或淡水浸泡约5min,取其浸泡液迅速倒入海水以保持体表寄生虫的存活。经浸泡过的鱼放于解剖盘中, 用手术剪刀截断延髓,然后剪开鳃腔、将每片鳃小心完整地取出, 放入盛有海水的培养皿中。用解剖针和镊子刮下鳃上皮, 使鳃上寄生虫脱落, 然后把培养皿中的混合物倒入大烧杯; 用海水将鳃分离物小心反复冲洗, 倒去上清液,经多次洗净污物后, 在解剖镜下挑取寄生虫, 用吸管将寄生虫吸到载玻片上, 盖上盖玻片并用指甲油固定盖玻片四角, 用固定液固定虫体。石斑鱼的鉴定参照相关文献资料(依据Heemstra et al, 1993; Froese et al, 2004)和FishBase (World Wide Web electronic publication, www.fishbase.org, version)。
1.2 生态术语
生态术语: 感染率(prevalence)、平均密度(abundance)、平均感染强度(mean intensity)、物种丰富度(species richness)、内群落(infracommunity)、组分群落(component community)等依据 Margolis等(1982)和 Bush等(1997), 存在分歧部分以 Bush等(1997)为准。
2 结果
本次调查, 共检查了 13种石斑鱼(体长范围11.0—27.0cm, 平均体长 19.4cm), 其中, 青石斑、橙点石斑、云纹石斑、鲑点石斑和玳瑁石斑5种石斑鱼(11.0—24.8cm, 18.87±3.20cm)检查 5尾及以上, 其余石斑鱼及网箱养殖的橙点石斑取样数少于5尾。本次调查从大亚湾海域石斑鱼中共检获 16种单殖吸虫,隶属于3科5属。检查了13种石斑鱼(橙点石斑野生和网箱养殖的都有), 发现有 10种鱼感染单殖吸虫,占调查鱼种的四分之三。
2.1 石斑鱼宿主与单殖吸虫的关系
2.1.1 石斑鱼寄生单殖吸虫组分群落的组成及丰度本次调查, 检查大亚湾海域石斑鱼体表及鳃的单殖吸虫, 其中, 点带石斑和未定种石斑分别检查1尾和2尾鱼未检获单殖吸虫、黑边石斑和大斑石斑体表单殖吸虫未统计, 鞍带石斑检查了1尾病鱼。表1比较了不同种石斑鱼寄生单殖吸虫种类、感染率、感染强度、方均比、平均密度及寄生部位。以感染强度来划分优势种(Rohde et al, 1994)。
从宿主-寄生虫关系看石斑鱼寄生单殖吸虫群落的多样性, 如表1可知, 除鞍带石斑和拟青石斑仅感染1种单殖吸虫, 其余几种石斑鱼感染单殖吸虫3—7种, 感染单殖吸虫种类最多的是玳瑁石斑(Epinephelus quoyanus)达7种, 次之是青石斑和养殖的橙点石斑均感染6种。感染强度最大的是拟合片虫(棕点石斑)128只/尾, 次之是石斑拟合片虫(青石斑)62只/尾, 接下来是斜带拟合片虫(大斑石斑)59只/尾和杯阴拟合片虫(玳瑁石斑)49只/尾。平均密度最高的是棕点石斑上检获的拟合片虫128只/尾。

表1 深圳大亚湾海域几种石斑鱼外寄生单殖吸虫种类及感染参数Tab.1 Occurrence and infection parameters of ectoparasitic monogeneans of Epinephelus spp. in Daya Bay, South China Sea

续表
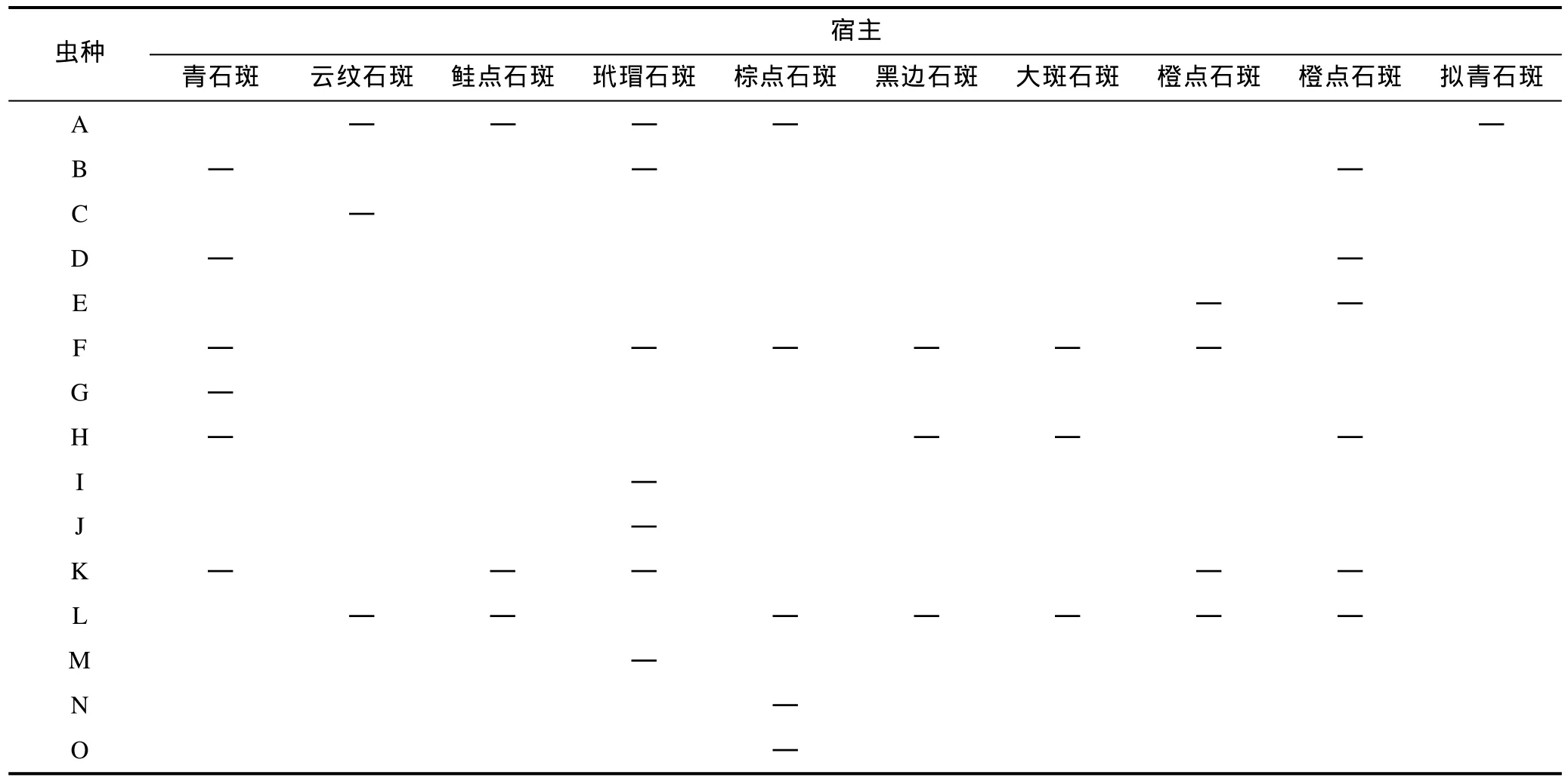
表2 深圳大亚湾海域石斑鱼寄生单殖吸虫群落组成Tab.2 Component community of ectoparasites of Epinephelus spp. in Daya Bay, South China Sea
在检查鱼数大于等于5尾的石斑鱼中, 玳瑁石斑单殖吸虫最高感染率 62.07%(18/29), 其单殖吸虫的群落为隶属于2科4个属的7种单殖吸虫: 体表寄生石斑本尼登虫和梅氏新本尼登虫, 鳃寄生石斑鳞盘虫、贾氏拟合片虫、“杯阴拟合片虫” (Species of “P.cupatus” group, Justine, 2005a)、坛状拟合片虫和南头拟合片虫。感染率由高到低依次为石斑本尼登虫(41%)、贾氏拟合片虫(38%)、南头拟合片虫(38%)和“杯阴拟合片虫”(21%)。
检查了3尾网箱养殖的橙点石斑, 都有检获单殖吸虫, 共 6个虫种, 隶属于 3个属。其中, 斜带拟合片虫的感染率最高(67%), 其次是梅氏新本尼登虫(50%); 石斑海盘虫、海盘虫未定种和南头拟合片虫的平均感染强度最高, 依次为14.0、9.0、9.0, 以上3个虫种在野生橙点石斑上都有检获。
检查的13种石斑鱼, 寄生体表的只有石斑本尼登虫和梅氏新本尼登虫两种, 其他虫种只在鳃上有检获,各单殖吸虫种群的方均比均大于1, 呈聚集分布。
2.1.2 单殖吸虫的宿主特异性 在调查的石斑鱼种类中, 检获 16种单殖吸虫, 可以两种以上宿主寄生的共 7个虫种, 占检获虫种数的 43.75%(7/16), 有: 石斑鳞盘虫、拟合片虫、石斑本尼登虫、斜带拟合片虫、南头拟合片虫、梅氏新本尼登虫和石斑海盘虫, 感染鱼种类数依次为: 6、6、5、4、4、3 和 2。
由表 2可知: 在检查的各种石斑鱼(除鞍带石斑和拟青石斑外)的寄生单殖吸虫群落组成中, 都至少有1个虫种可以寄生在其他种类的宿主上。这种在一定分类水平上, 宿主-寄生虫的共专一可以用嵌套结构来表示, 即在石斑鱼属或几个相近的宿主种类中一些共有虫种组成了它们的外寄生虫群落。
2.2 石斑鱼寄生单殖吸虫内群落物种丰富度及其频率分布
由表3可知: 在调查的5种石斑鱼寄生单殖吸虫内群落中, 在橙点石斑体表未见单殖吸虫, 鳃上的单殖吸虫由 0—3个虫种组成, 平均值为 0.42; 玳瑁石斑体表上的单殖吸虫为0—2个虫种, 平均0.48, 鳃上的单殖吸虫为 0—4个虫种, 平均 1.21, 外寄生(体表+鳃)0—5个虫种, 平均 1.69; 在鲑点石斑体表和鳃都分别发现1个单殖吸虫虫种, 外寄生1个虫种; 在青石斑体表上检测到0—1个虫种, 鳃上0—3个虫种,外寄生0—3个虫种; 在云纹石斑体表上检测到0—1个虫种, 鳃上0—1个虫种, 外寄生0—2个虫种。
由表3可知: 在 19尾橙点石斑的寄生单殖吸虫内群落中(体表无寄生虫), 由2个和3个虫种组成的内群落各1个, 由1个虫种组成的内群落有3个; 在检查的 11尾青石斑中, 其寄生单殖吸虫内群落由 2个和3个虫种组成的宿主各有1尾, 由1个虫种组成的有3尾; 检查了5尾云纹石斑, 发现由1个和2个虫种组成的宿主各有1尾; 检查了29尾鲑点石斑, 在4尾鱼上发现的内群落都是由1个虫种组成; 在检查的29尾玳瑁石斑中, 其寄生单殖吸虫内群落由1个、2个、4个、5个虫种组成的宿主, 分别有6尾、5尾、2尾和5尾。以上这些石斑鱼寄生单殖吸虫内群落中,最多可以由4个属的虫种组成: 在玳瑁石斑的1个寄生单殖吸虫内群落中, 组成内群落的来自4个属虫种是: 石斑本尼登虫、梅氏新本尼登虫、石斑鳞盘虫和驼背鲈海盘虫。
2.3 石斑鱼寄生单殖吸虫内群落物种丰富度与宿主体长的关系
将宿主体长与其感染的单殖吸虫物种数做相关分析(本文采用Spearman秩和等级相关分析, 设定显著性检验水准P=0.05)。如表4可知: 检查了这 5种石斑鱼, 发现仅玳瑁石斑寄生单殖吸虫内群落物种丰富度与宿主体长呈极显著的正相关(P<0.01)。
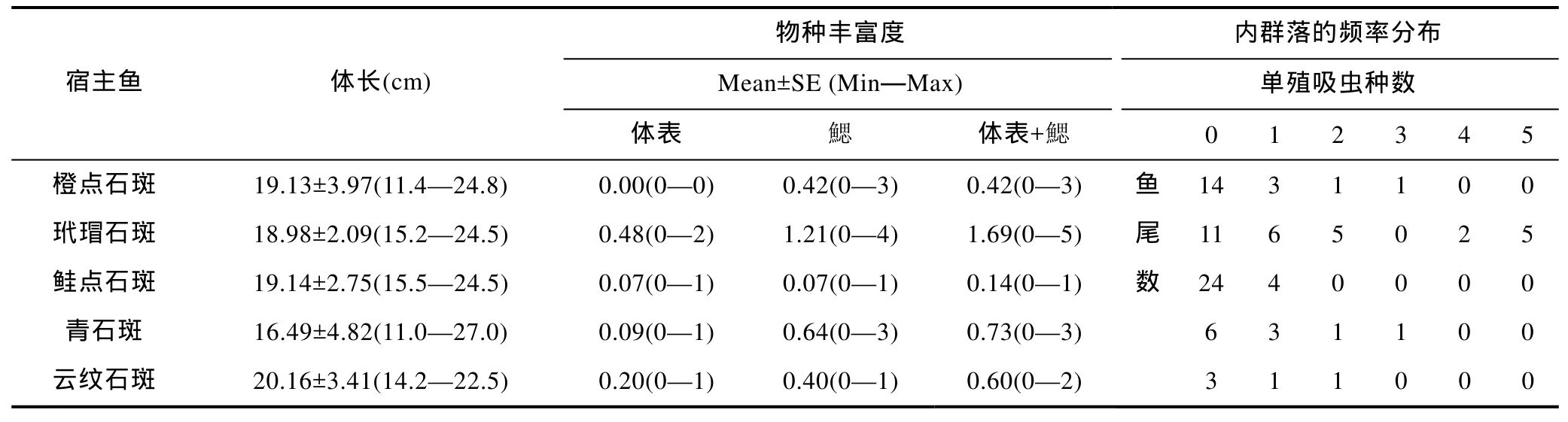
表3 深圳大亚湾海域5种石斑鱼寄生单殖吸虫内群落的物种丰富度和频率分布Tab.3 Species richness and frequency distribution of infracommunities of monogeneans of 5 species of Epinephelus spp. in Daya Bay,South China Sea

表4 深圳大亚湾海域5种石斑鱼体长与寄生单殖吸虫物种丰富度的相关分析Tab.4 Correlation analysis between species richness of monogeneans and body length of 5 fish species of Epinephelus spp. from Daya Bay, South China Sea by Spearman rank coefficients
3 讨论
3.1 寄生虫多样性
本研究共调查了13种石斑鱼, 有5种宿主检查5尾以上, 共 10种鱼检获单殖吸虫, 占调查鱼种类的四分之三; 共检获15种单殖吸虫, 隶属于3科5属。除拟青石斑(检查4尾)和鞍带石斑(检查1尾)外, 其他8种石斑鱼寄生单殖吸虫群落至少都由 3个虫种组成。Oliver(1987)认为鳞盘虫科的单殖吸虫有着严格的宿主特异性, 可以用来估算寄生虫多样性。林丽红(Lim, 1998)总结了东南亚鱼类寄生虫多样性认为每种宿主鱼的寄生虫种数大约为3种; Whittington(1998)估计每种宿主鱼有 5种; Justine等(2010)在 New Caledonia的对科鱼类调查分析后认为每种宿主鱼寄生虫可达10种。林丽红(Lim, 1998)整理总结了东南亚鱼类单殖吸虫的多样性, 认为, 这个区域有着最多的淡水鱼类大约有105科, 而南美有约60科, 非洲有 74科; 在这个地区鱼的种类多样, 单殖吸虫所知甚少。单殖吸虫在该区域的水生动物中的数量可能被低估了, 因为检查的宿主鱼极少/如此少, 她估计在马来西亚 Peninsular单殖吸虫可能的数量显示仅知8%; 在东南亚自从20世纪30年代第一次描述单殖吸虫已过了67年, 仅描述了246种; 在最近16年报道的数量明显上升, 达到每年 10个种, 而 1930—1980则是每年2个种。本次对大亚湾海域石斑鱼单殖吸虫的调查, 8种石斑鱼寄生单殖吸虫3—7种, 远远多于林丽红估算的3种。另外, 有研究表明: 寄生虫群落的物种丰富度有从温带到热带逐渐升高的趋势(Rohde, 1978; Rohde et al, 1998)。本文的宿主取样点大亚湾, 位于广东东部沿海, 属于亚热带气候, 终年气温较高。这种特殊的地理位置也可能是影响斜带石斑等宿主上外寄生单殖吸虫群落物种组成丰富度的原因之一。
3.2 寄生虫宿主特异性
在检查的各种石斑鱼(除鞍带石斑外)其寄生单殖吸虫群落组成中, 都至少有1个虫种也可以寄生在另外的宿主上。即在石斑鱼属下或几个相近的宿主种类中, 一些共有虫种组成了它们的外寄生虫群落。Gutiérrez(1999a, b, 2001)调 查 阿 根 廷 的 斑 油 鲶(Pimelodus maculatus)和白油鲶(P. albicans)鳃上单殖吸虫群落, 发现共有种的比例和两个宿主的系统发生及生态习性有关联, 支持单殖吸虫和该河流(Río de la Plata)油鲶共专一性的假说; 认为这些寄生虫群落的物种聚集不大可能是由偶然机遇形成的, 提出在Río de la Plata河流的宿主寄生虫系统中, 系统发生的类似性意味着有一个单一的群落模式, 可以在同属和同域的宿主中观察到。Buchmann等(2002)总结了单殖吸虫和它们的宿主鱼之间的相互作用, 认为: 寄生虫的感受器能够检测不同鱼种之间的不同,这种洞察区分鱼种的能力可能基于宿主鱼体表的化学和物理刺激; 单殖吸虫对宿主鱼的附着是依赖于寄生虫的结构和化学因子; 对一定寄生虫易感的宿主鱼, 在感染后期该宿主鱼表现出保护应答能力的升高。宿主-寄生虫识别与作用的机制可能也是形成石斑鱼外寄生群落的作用因子之一。
本次研究发现: 这些石斑鱼外寄生单殖吸虫群落大部分是由石斑鱼属的广寄生单殖吸虫种(generalist)组成, 有7种单殖吸虫隶属于3科5属(如表 2), 这些虫种在石斑鱼属是普遍寄生的。Vidal-Martínez等(2003)研究了海水鱼类黑缘石斑(Epinephelus morio)(仅一种单殖吸虫拟合片虫 P.yacatanensis)和淡水鱼类尾斑丽体鱼寄生虫群落的时空重复性, 寄生虫在 4个不同的取样点有嵌套结构,可能是因为它们有不同的生物地理学史。Poulin(1997)把宿主特异性作为讨论嵌套结构的一个影响因子,但他在不同外寄生虫物种聚集的嵌套结构里并没有发现宿主特异性(专一性)的影响。Rohde等(1998)强调: 寄生虫群落的嵌套结构并不表明种间竞争的存在。Matějusová等(2000)研究了两种鲤科鱼类寄生三代虫的广寄生虫种和专一性虫种的嵌套结构;Simková等(2001)提到“孤岛”理论激发了更多的对群落结构问题的讨论, 嵌套结构模式的发生指示: 寄生虫群落不是一个纯粹的随机结构组织。这与本文的结果, 在不同的宿主中单殖吸虫群落共有种的普遍存在及不同时间, 同一宿主单殖吸虫群落中一些虫种的共存和隔离是一致的, 在不同石斑鱼宿主的寄生虫群落中普遍存在的石斑鳞盘虫有显著的宿主特异性(即: 目前的研究表明石斑鳞盘虫仅发现于石斑鱼上)。李文祥等(2014)研究了洄游型、淡水型和陆封型3种生态型刀鲚的寄生蠕虫群落, 为寻找区分其寄生虫标志, 发现其海水性寄生虫, 可作为区分洄游型刀鲚的寄生虫标志, 而淡水性寄生虫只在淡水型刀鲚中发现, 可作为淡水型刀鲚的寄生虫标志。石斑鱼属外寄生的石斑鳞盘虫也可以考虑为该属鱼类的寄生虫标志。
Brown(1984)提出分布-密度关系: 可以利用很多资源的广寄生虫种可以以较高的密度广泛地分布,而仅利用极少资源的专一性虫种的分布将很局限,并且密度相对的贫乏。Barger等(2001)调查了美国北卡罗莱纳洲阿帕拉契山7个小溪流的14个位点的18种鱼上寄生虫(22种)的群落, 检验了 Brown的资源-宽度假说(分布-密度关系), 发现这些寄生虫种分布(占用的位点片区)与密度(位点的平均密度)显著的正相关, 结果支持 Brown(1984)的资源-宽度假说。Brown(1984)的资源-宽度假说里, 在单一宿主鱼种群中, 因为广寄生虫种的密度难以评估和定义而不能检验。就本文的结果而言, 广寄生虫种既在相对广泛的宿主分布(可占用的资源片区), 同时也发现有相对高的感染率和平均感染强度, 大多数是寄生群落的优势种, 也支持Brown(1984)的资源-宽度假说。
3.3 寄生虫群落物种丰富度与宿主体长的关系
Lo等(1998)对法属波利尼西亚岛的三种珊瑚礁鱼类, 黑真雀鲷(Stegastes nigricans)、三带圆雀鲷(Dascyllus aruanus)和斑点九棘鲈(Cephalopholis argus)的寄生虫群落进行了调查, 黑真雀鲷和斑点九棘鲈的外寄生虫群落物种丰富度与宿主的年龄和体长正相关; 并认为外寄生虫群落似乎受生物因子影响更大(例如宿主免疫等)。本文的结果显示, 玳瑁石斑的外寄生虫群落物种丰富度与宿主的体长呈极显著的正相关, 与Lo等(1998)的结果一致。本文所调查的其它几种石斑鱼的内群落, 群落物种丰富度与宿主体长之间未见相关关系, 可能是样本量较少的缘故。
4 结论
大亚湾海域石斑鱼寄生单殖吸虫种类丰富(3—7种)。单殖吸虫群落大部分是由石斑鱼属的广寄生单殖吸虫种(generalist)组成。石斑鳞盘虫在石斑鱼属的不同宿主鱼上普遍检出, 可考虑为该属鱼类的寄生虫标志。
卢明淼, 李文祥, 王微微等, 2015. 长江中下游三个湖泊中鳜消化道寄生蠕虫群落的组成与多样性. 水生生物学报,39(2): 301—306
吕军仪, 吴金英, 杨大伟等, 2001. 鲻鱼寄生蠕虫种群和群落组成分析. 动物学报, 47(6): 609—615
李文祥, 王桂堂, 2014. 洄游型、淡水型和陆封型刀鲚的寄生蠕虫群落结构. 动物学杂志, 49(2): 233—243
吴金英, 吕军仪, 曾 华等, 2001. 鲻鱼(Mugil cephalus)寄生蠕虫群落生态研究. 生态学报, 21(6): 1003—1008
张剑英, 1999. 鱼类寄生虫与寄生虫病. 北京: 科学出版社
张剑英, 2001. 中国海洋鱼类单殖吸虫. 北京: 中国农业出版社
Barger M A, Esch G W, 2002. Host specificity and the distribution-abundance relationship in a community of parasites infecting fishes in streams of North Carolina.Journal of Parasitology, 88(3): 446—453
Brown J H, 1984. On the relationship between abundance and distribution of species. The American Naturalist, 124(2):255—279
Bu S S H, Leong T S, Wong S Y et al, 1999. Three diplectanid monogeneans from marine finfish (Epinephelus spp.) in the Far East. Journal of Helminthology, 73(4): 301—312
Buchmann K, Lindenstrøm T, 2002. Interactions between monogenean parasites and their fish hosts. International Journal for Parasitology, 32(3): 309—319
Bush A O, Lafferty K D, Lotz J M et al, 1997. Parasitology meets ecology on its own terms: Margolis et al. Revisited. The Journal of Parasitology, 83(4): 575—583
Cribb T H, Bray R A, Wright T et al, 2002. The trematodes of groupers (Serranidae: Epinephelinae): knowledge, nature and evolution. Parasitology, 124(7): 23—42
Cruz-lacierda E R, Toleda J D, Tan-Fermin J D et al, 2000.Marine leech (Zeylanicobdella arugamensis) infestation in cultured orange-spotted grouper, Epinephelus coioides.Aquaculture, 185(3—4): 191—196
Froese R, Pauly D, 2004. FishBase, World Wide Web electronic publication. www.fishbase.org
Gutiérrez P A, Martorelli S R, 1999a. The structure of the monogenean community on the gills of Pimelodus maculatus in Río de la Plata (Argentina). Parasitology, 119(2): 177—182
Gutiérrez P A, Martorelli S R, 1999b. Niche preferences and spatial distribution of Monogenea on the gills of Pimelodus maculatus in Río de la Plata (Argentina). Parasitology,119(2): 183—188
Gutiérrez P A, 2001. Monogenean community structure on the gills of Pimelodus albicans from Río de la Plata (Argentina):a comparative approach. Parasitology, 122(4): 465—470
Heemstra P C, Randall J E, 1993. FAO Species Catalogue. Vol.16. Groupers of the world (Family Serranidae, Subfamily Epinephelinae). An annotated and illustrated catalogue of the grouper, rockcod, hind, coral grouper and lyretail species known to date. FAO Fisheries Synopsis, No.125, Volume 16.Rome: FAO.
Justine J L, 2005a. Species of Pseudorhabdosynochus Yamaguti,1958 (Monogenea: Diplectanidae) from Epinephelus fasciatus and E. merra (Perciformes: Serranidae) off New Caledonia and other parts of the Indo-Pacific Ocean, with a comparison of measurements of specimens prepared using different methods, and a description of P. caledonicus n. sp.Systematic Parasitology, 62(1): 1—37
Justine J L, 2005b. Pseudorhabdosynochus hirundineus n. sp.(Monogenea: Diplectanidae) from Variola louti (Perciformes:Serranidae) off New Caledonia. Systematic Parasitology,62(1): 39—45
Justine J L, 2007a. Parasite biodiversity in a coral reef fish:twelve species of monogeneans on the gills of the grouper Epinephelus maculatus (Perciformes: Serranidae) off New Caledonia, with a description of eight new species of Pseudorhabdosynochus (Monogenea: Diplectanidae).Systematic Parasitology, 66(2): 81—129
Justine J L, 2007b. Pseudorhabdosynochus argus n. sp.(Monogenea: Diplectanidae) from Cephalopholis argus, P.minutus n. sp. and Diplectanum nanus n. sp. from C.sonnerati and other monogeneans from Cephalopholis spp.(Perciformes: Serranidae) off Australia and New Caledonia.Systematic Parasitology, 68(3): 195—215
Justine J L, Vignon M, 2009. Monogeneans of the grouper Epinephelus tauvina (Perciformes, Serranidae) off Moorea,French Polynesia, with a description of Pseudorhabdosynochus pai n. sp. (Monogenea:Diplectanidae). Systematic Parasitology, 72(2): 113—125
Justine J L, Beveridge I, Boxshall G A et al, 2010. An annotated list of parasites (Isopoda, Copepoda, Monogenea, Digenea,Cestoda and Nematoda) collected in groupers (Serranidae,Epinephelinae) in New Caledonia emphasizes parasite biodiversity in coral reef fish. Folia Parasitologica, 57(4):237—262
Leong T S, Wong S Y, 1988. A comparative study of the parasite fauna of wild and cultured grouper (Epinephelus malabaricus Bloch et Schneider) in Malaysia. Aquaculture,68(3): 203—207
Li W X, Arnott S A, Jones K M M et al, 2015. First record of paratenic hosts of the swimbladder nematode Anguillicola crassus in North America. Journal of Parasitology, 101(5): 529—535
Lim L H S, 1998. Diversity of monogeneans in Southeast Asia.International Journal for Parasitology, 28(10): 1495—1515
Lo C M, Morand S, Galzin R, 1998. Parasite diversity/host age and size relationship in three coral-reef fishes from French Polynesia. International Journal for Parasitology, 28(11):1695—1708
Luo Y F, Brown C L, Yang T B, 2010a. Seasonal dynamics of Diplectanum grouperi parasitism on wild versus cultured groupers, Epinephelus spp., and the linkage between infestation and host species phylogeny. Journal of Parasitology, 96(3): 541—546
Luo Y F, Yang T B, 2010b. Seasonal population dynamics of the monogeneans Pseudorhabdosynochus coioidesis and P.serrani on wild versus cultured groupers in Daya Bay, South China Sea. Helminthologia, 47(4): 241—250
Luo Y F, Yang T B, 2012. Seasonal patterns in the community of gill monogeneans on wild versus cultured orange-spotted grouper, Epinephelus coioides Hamilton, 1822 in Daya Bay,South China Sea. Aquaculture Research, 43(8): 1232—1242 Margolis L, Esch G W, Holmes J C et al, 1982. The use of ecological terms in parasitology (report of an ad hoc committee of the American society of Parasitologists). The Journal of Parasitology, 68(1): 131—133
Matějusová I, Morand S, Gelnar M, 2000. Nestedness in assemblages of gyrodactylids (Monogenea: Gyrodactylidea)parasitising two species of cyprinid-with reference to generalists and specialists. International Journal for Parasitology, 30(11): 1153—1158
Moravec F, Vidal-Martínez V M, Vargas-Vásquez J et al, 1997.Helminth parasites of Epinephelus morio (Pisces: Serranidae)of the Yucatan Peninsula, southeastern Mexico. Folia Parasitologica, 44(4): 255—266
Nolan M J, Cribb T H, 2004. Two new blood (Digenea:Sanguinicolidae) from Epinephelinae (Perciformes:Serranidae) of the Pacific Ocean. Parasitology International,53(4): 327—335
Oliver G, 1987. Les Diplectanidae Bychowsky, 1957 (Monogenea,Monopisthocotylea, Dactylogyridea). Systématique.Biologie. Ontogénie. Écologie. Essai de phylogenèse.Montpellier: Doctoral Dissertation of Université des Sciences et Techniques du Languedoc, 1—433
Poulin R, 1997. Parasite faunas of freshwater fish: the relationship between richness and the specificity of parasites.International Journal for Parasitology, 27(9): 1091—1098
Rohde K, 1978. Latitudinal differences in host-specificity of marine Monogenea and Digenea. Marine Biology, 47(2):125—134
Rohde K, Hayward C, Heap M et al, 1994. A tropical assemblage of ectoparasites: gill and head parasites of Lethrinus miniatus (Teleostei, Lethrinidae). International Journal for Parasitology, 24(7): 1031—1053
Rohde K, Heap M, 1998. Latitudinal differences in species and community richness and in community structure of metazoan endo- and ectoparasites of marine teleost fish.International Journal for Parasitology, 28(3): 461—474
Simková A, Gelnar M, Sasal P, 2001. Aggregation of congeneric parasites (Monogenea: Dactylogyrus) among gill microhabitats within one host species (Rutilus rutilus L.).Parasitology, 123(6): 599—607
Vidal-Martínez V M, Mendoza-Franco E F, 1998.Pseudorhabdosynochus capurroi sp. n. (Monogenea:Diplectanidae) from the gills of Mycteroperca bonaci(Pisces: Serranidae) of the Yucatan Peninsula, Mexico. Folia Parasitologica, 45(3): 221—224
Vidal-Martínez V M, Poulin R, 2003. Spatial and temporal repeatability in parasite community structure of tropical fish hosts. Parasitology, 127(4): 387—398
Whittington I D, 1998. Diversity “down under”: monogeneans in the Antipodes (Australia) with a prediction of monogenean biodiversity worldwide. International Journal for Parasitology, 28(10): 1481—1493
