益生元对早产儿、肥胖及老年群体肠道菌群调节机制的研究现状
2018-01-08郑志昌陈映彤郭娟娟郭泽镔郑宝东
郑志昌,陈映彤,郭娟娟,郭泽镔,郑宝东,卢 旭*
(福建农林大学食品科学学院,福建 福州 350002)
益生元对早产儿、肥胖及老年群体肠道菌群调节机制的研究现状
郑志昌,陈映彤,郭娟娟,郭泽镔,郑宝东,卢 旭*
(福建农林大学食品科学学院,福建 福州 350002)
肠道微生物是人体不可或缺的一部分,微生物多样性和群落结构的稳定性与机体健康息息相关。临床研究表明,肠道微生物数量在特殊人群的疾病发展中扮演着重要角色。益生元作为人体膳食重要的组成部分,主要为植物源的非消化性低聚糖和膳食纤维,其具有选择性促进肠道内特定菌群增殖与活力的效果。本文以肠道微生物在人体中的变化过程为研究对象,主要对比3 类特殊人群(早产儿、肥胖人群及老年人)的肠道微生物定植差异以及益生元对此类人群肠道菌群的调节作用。为益生元的肠道菌群调节机理和益生元功能性食品开发提供理论依据。
益生元;肠道菌群;早产儿;肥胖;老年人
正常机体胃肠道中的微生物群落依照与机体间的关系大致可分为3大类[1]:有益菌、有害菌和潜在致病菌,三者间存在相互竞争和拮抗的关系,并构成重要的微生态系统。近现代医学研究表明,肠道菌群对人体营养物质代谢、机体发育、免疫及疾病的产生等方面都起到了极其重要的作用,因此被称为人体的“超级组织”[2]或“虚拟器官”[3]。肠道微生物能够利用广泛的食物基质和寄主分泌物产生多种代谢产物,如胆汁酸衍生物、维生素、支链脂肪酸(branched-chain fatty acids,BCFAs)和短链脂肪酸(short-chain fatty acids,SCFAs)等,对寄主生理健康可产生局部或系统性的影响。同时,微生物也是导致肠道感染、炎症性肠病、结肠癌、肥胖、糖尿病、心血管疾病、食物过敏等代谢或免疫性疾病的关键因素之一。
当机体肠道中有益菌数量占优势时,有利于人体健康;反之当肠道微生态系统中有害菌处于优势时,菌群结构平衡遭受破坏后,则引发相关疾病的概率将会大为增加。益生元可选择性地促进双歧杆菌、乳杆菌等益生菌的生长或活力,并通过益生菌形成优势菌群进而竞争性地拮抗有害菌,或通过其代谢作用产生SCFAs和细菌素等物质抑制有害菌的生长,此外益生菌内部的竞争关系还可通过负反馈回路抵消菌群多样性造成的不稳定[4]。肠道微生物数量在不同时间和空间中将会产生巨大的差异,不同人群与发育阶段对肠道菌群的结构和稳定性具有显著性影响,膳食补充益生元作为干预肠道菌群的有效措施,近年来被广泛应用于某些特殊人群的肠道调理中。本文主要概述了有关益生元的发展、人体肠道微生物的演变过程及其影响因素、3 类特殊人群肠道菌群的结构特点及益生元对肠道菌群的调节作用。
1 益生元
益生元,最早在1995年由英国的Gibson等[5]提出,其被定义为一种不被消化的食物成分,摄入后不被小肠消化吸收而直接到达大肠,进而通过选择性地促进大肠中一种或有限的几种细菌的生长或活力对寄主产生健康效应。最初的益生元物质仅限于少数几种非消化性的低聚糖,其中低聚果糖作为被广泛认可的一种益生元,已被视为功能性成分应用于食品及保健品中。
经过20余年的研究,益生元的定义和范围不断丰富。图1为益生元定义被修改的历程,最新的版本是由Bindels等[6]于2015年在Perspectives杂志上发表,其将益生元最终定义为“通过肠道内微生物的代谢作用并可调节肠道微生物的组成或活力,从而对寄主健康产生有益的生理影响的非消化性成分”。文章在承认低聚果糖、反式低聚半乳糖、菊粉以及人乳低聚糖(human milk oligosaccharide,HMOS)等公认益生元地位的基础上,同时将抗性淀粉、果胶、阿拉伯木聚糖、全谷物、膳食纤维以及其他具有调节肠道微生物作用的非碳水化合物列为潜在益生元。此次修订恰逢益生元定义产生20周年,文章进一步扩大了益生元的范畴;文中认为,以往有关益生元的研究大都集中于选择性促进作用、生理影响及其与肠道微生物组成或代谢的关系,而有关建立肠道菌群因果作用的研究仍然有限;因此,Bindels等希望今后将益生元的研究重点从对肠道菌群组成影响的定性研究转移至对健康影响得以实现的机制上去。这对于帮助理解膳食-微生物-寄主之间相互作用关系具有重要的指导意义。
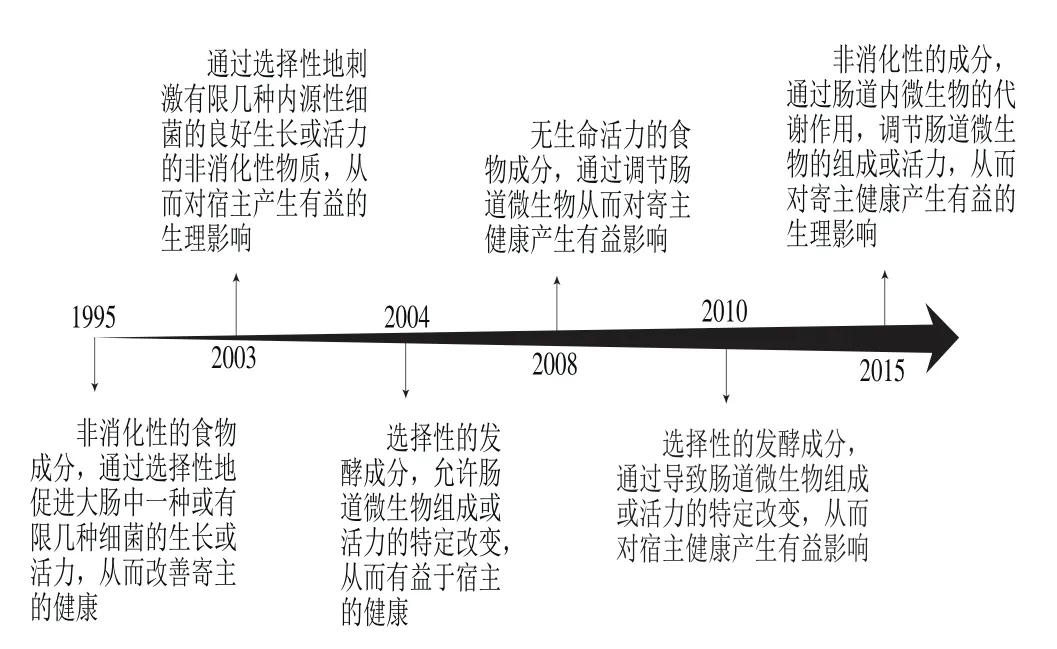
图1 益生元定义的演变史Fig. 1 Evolutionary history of prebiotics
2 机体肠道微生物的组成与变化
2.1 肠道微生物组成
据文献报道,人类肠道菌群约有100余属,超过400 个种,总质量达到1 000 g,其个体总数约为1014个[7]。每克肠道内容物约有1011~1012个细菌,占粪便干质量的1/3以上,以西式饮食为主的人员,其肠道细菌总量可达肠道内容物的50%以上。肠道微生物主要以厌氧和兼性厌氧菌为主,少数为需氧菌,大多数(70%~80%)为不可分离培养的细菌类型。微生物在肠道中的空间分布具有明显的区域性,这与食物种类、通过速度和不同区域的pH值密切相关。大肠中食物缓慢的蠕动速度和适宜的pH值环境使得微生物能够建立复杂而相对稳定的群体,因此大肠为非消化性食物成分以及内源性黏蛋白发酵的理想场所;微生物群落空间分布区域根据特定微环境的不同可进一步细分为内腔、黏液、隐窝和上表皮细胞分布层。
2.2 肠道微生物随年龄的变化历程
人类胚胎初始为无菌状态,此后由分娩到最初的几个月相继经历了各种微生物此消彼长的过程[8]。在婴儿出生前,其从母体产道获得接种物,随后婴儿肠道开始出现定植的微生物。最初以肠杆菌、肠球菌和链球菌等兼性厌氧菌为主体(第一阶段),这部分细菌在早期48 h内消耗肠道中的氧气,降低肠道的氧化还原电位,为双歧杆菌等绝对厌氧菌的生长创造有利环境(第二阶段)。婴儿出生后5 d左右,双歧杆菌开始形成优势菌群,以母乳喂养的婴儿双歧杆菌数量在哺乳期中保持绝对优势,在哺乳末期双歧杆菌相对减少,随着固体食物的摄入,哺乳期婴儿的肠道菌群开始从低丰度与低复杂性逐渐演变成熟,1周岁婴儿的肠道微生物构成逐渐趋于成年人,菌群相对稳定,不同婴儿间的菌群功能差异逐渐减小[9]。随着年龄递增,肠道菌群受生理状态、饮食模式、生活习惯等内外因素的影响,维持着以厚壁菌-拟杆菌-放线菌门为主的成年人型菌群结构[10-11]。直到中老年阶段,双歧杆菌数量显著下降,梭菌和肠杆菌数量呈逐渐递增趋势。
3 影响肠道菌群结构的环境因素
作为机体生态系统不可或缺的一部分,肠道微生态系统中的菌群结构在菌群间、菌与寄主间的相互作用维持着相对稳定的动态平衡。诸多因素可以影响肠道微生物的构成与数量,主要因素包括寄主的遗传特性、分娩方式[9,12]、出生胎龄、哺乳方式、膳食模式、年龄与身体状态以及是否接受药物治疗等内源性与环境因子。
已有大量研究结果表明,母乳喂养与人工乳粉喂养的婴儿粪便中具有不同的微生物种群。相比于人工乳粉喂养,采用母乳喂养的婴儿,其粪便中的双歧杆菌数量和比例显著较高,而以乳粉所喂养婴儿肠道中的成人型细菌种类较多,且具有更加复杂的菌群组成。有学者认为,婴儿出生后的喂养过程为肠道菌群筛选过程[13]。在肠道微生物发展的第二阶段,婴儿的饮食组成对微生物种类起到了整合作用。
膳食模式是塑造肠道菌群的重要环境因素,膳食宏量营养素中的碳水化合物、蛋白质和脂肪的数量、种类和比例是影响肠道菌群构成的主要原因。人体结肠中的庞大菌群是绝对的厌氧菌,现代基因组学研究表明,双歧杆菌的基因组中有更多用于编码与碳水化合物代谢相关的酶系的基因片段[14],因此,双歧杆菌能优先发酵每天约40 g来源于上消化道的非消化性碳水化合物,生成乙酸和乳酸,降低大肠环境pH值,从而抑制腐败菌的繁殖。而每天来源于上消化道的非消化性蛋白质约为25 g,这些蛋白质主要被拟杆菌和梭菌发酵转变成异丁酸、异戊酸等BCFAs以及一些无机氮化合物,部分终产物可能对寄主产生毒性[15]。有学者认为增加膳食中非消化性碳水化合物的供应能够促进益生菌在增殖过程中的蛋白质合成能力与多肽利用率,从而限制蛋白质过度发酵所产生的有毒产物[16]。研究表明以植物性食物和以高蛋白食物为主食的两种膳食模式对肠道微生物组成具有深远影响,前者肠道微生物以拟杆菌门为主,而后者以厚壁菌门为主[17]。
膳食性脂肪几乎都在小肠消化吸收,但是过高脂肪摄入会导致胆汁酸等肝代谢产物和未参与肝内循环的物质更多地进入大肠;因此有学者认为,膳食脂肪对肠道菌群的影响可能是通过胆汁酸间接介导的结果[18],但也有学者提出肠道微生物组成的改变与内毒素含量增加所导致的肠道通透性提高和内脏白色脂肪组织中基质血管细胞数目的急剧增加有关[19]。动物模型研究表明,高脂膳食可显著降低粪便中SCFAs含量和双歧杆菌、罗氏菌属数量,而通过补充低聚果糖和阿拉伯木聚糖均能够恢复二者的数量[20-21]。王志凡等[22]在高脂膳食诱导肥胖大鼠模型中发现,高脂膳食组大鼠粪便鲜质量、含水率以及肠球菌、双歧杆菌、乳酸菌、类杆菌数量显著下降。Anitha等[23]研究发现,高脂膳食小鼠粪便中厚壁菌门和大肠杆菌属的数量与正常组相比更多,但拟杆菌门数量相对较少,高脂膳食组通过补充低聚果糖能够防止肠道氮神经元的损失,改善肠道蠕动时间,减少粪便中厚壁菌门和大肠杆菌属的数量。Serino等[19]对高脂膳食小鼠补充低聚半乳糖后发现,拟杆菌与厚壁菌门丰度比值急剧增加,而放线菌门几乎消失。
4 益生元对特殊人群肠道菌群的调节作用
4.1 益生元与早产儿
4.1.1 早产儿的微生态生理特点
早产儿一般泛指妊娠期小于37 周或出生体质量小于1 500 g的新生儿,其出生后常无法获得足够的母乳喂养。由于其肠道消化系统和黏膜免疫系统还未发育成熟,导致其对食物的耐受性和肠道微生物的定植尤其敏感。早产儿肠道微生物定植延迟,且存在个别菌群过度生长、多样性低以及存在与坏死性小肠结肠炎的发生有关的潜在致病菌,如梭菌、大肠杆菌和肺炎克雷伯氏菌属等细菌是造成早产儿发病和致死的重要原因。由于早产儿对多数喂养食物的不耐受性,全胃肠外营养通常作为早产儿的初始喂养策略,但是这可能导致某些肠道微生物延迟定植[1];另一方面,早产儿脆弱的免疫系统无法抵御环境微生物的入侵,从而增加了细菌过度繁殖的风险;此外抗生素的滥用进一步干预了肠道天然微生物的多样性定植。
4.1.2 HMOS对肠道微生态的重要性
HMOS是母乳中质量浓度仅次于蛋白质和脂质的第三大类活性成分,因个体差异其质量浓度约为5~15 mg/mL,HMOS包含一系列以乳糖作为还原性末端的复杂糖链,已知的HMOS约有200 种,包括中性和酸性低聚糖[23]。它们对肠道免疫系统成熟、肠道功能完整性与微生物定植至关重要。HMOS被认为是导致母乳与人工喂养的婴儿在肠道菌落组成和发展差异的本质原因。HMOS可作为双歧因子直接促进双歧杆菌等益生菌的增殖,调节肠道黏膜免疫通路,促进黏膜免疫系统成熟,间接改善细菌定植的敏感性,从而调节肠道菌群数量。He Yingying等[24]研究发现,人初乳低聚糖(human colostrum oligosaccharides,cHMOS)能减少病原体相关分子模式刺激急性期的白细胞介素(interleukin,IL)-8、IL-6、IL-1β以及单核细胞趋化蛋白-1等促炎因子的水平,同时提高机体组织修复和维持内环境稳态的有关细胞因子表达。
4.1.3 其他益生元对早产儿肠道菌群的影响
由于HMOS对于婴儿尤其是早产儿具有重要作用,因此可以通过在配方乳粉中添加益生元以减少其与人乳的营养差异,以提高两者喂养的婴儿肠道微生物的相似性。有研究表明,早产儿肠道中的双歧杆菌和乳杆菌密度低于正常产期的婴儿[25-26],因此膳食喂养促进二者在早产儿肠道中定植对于微生物在新生儿肠道中定植、肠道固有免疫和适应性免疫的发展成熟不可或缺。Ripoll等[27]发现在配方乳粉中补充5 g/L的短链低聚果糖(shortchain fructo-oligosaccharides,scFOS)时,其安全性和耐受性良好,并且发现scFOS可通过增加双歧杆菌数量调节肠道微生物组成;Westerbeek等[28]研究也发现,通过补充中性(短链低聚半乳糖、长链低聚果糖)和酸性低聚糖(海藻酸钠低聚糖)混合物能够促进早产儿产后肠道微生物定植,而广谱抗生素的使用则会产生相反的结果;短链低聚半乳糖和长链低聚果糖安全性和对肠道的耐受性也已在早产儿体内实验中得到证实[29]。Underwood等[30]发现,HMOS对早产儿肠道微生物的影响可能与特定结构的低聚糖有关,含有岩藻糖化与唾液酸化的HMOS能使早产儿粪便中的变形菌门增多而厚壁菌门数量相对减少。
4.2 益生元与肥胖人群
4.2.1 肠道微生物对肥胖寄主代谢的影响
肥胖问题如今已成为世界尤其是发达国家的主要公共性社会问题之一,肥胖的本质为流行性疾病,目前正呈现出由西方发达国家向大多数发展中国家迅速蔓延的趋势。肥胖的发生与遗传和环境因素有关,权威报告指出肠道微生物在脂肪含量增加和机体能量平衡中发挥着关键作用。肠道微生物与肥胖有关的作用机制包括以下几个方面[31]:1)增加微生物吸收膳食中能量的能力;2)调节肝脏脂肪生成;3)通过肠道饱腹感激素调节食欲,如胰高血糖素样肽(glucagon-like peptide,GLP)-1和酪酪肽;4)通过脂多糖(lipopolysaccharide,LPS)-Toll样受体激活机体自身免疫能力。
2013年,Ridaura等[32]将具有一胖一瘦体型的若干对双胞胎的粪便微生物移植到悉生小鼠体内,结果发现接受肥胖型(Ob组)粪便的小鼠体质量和脂肪指数显著高于接受非肥胖型(Ln组)粪便的小鼠,研究认为导致这种差异的原因可能与Ln组小鼠中肠道微生物的SCFAs发酵作用更强有关,此外Ln组小鼠肠道微生物还可通过下调法尼醇X受体信号进而增加胆汁酸类物质转化,增加脂肪和胰岛素抵抗性;而Ob组小鼠中支链氨基酸代谢量有所增加。最近,Suárez-Zamorano等[33]通过给予经抗生素剔除肠道菌群或无菌的小鼠高脂肪膳食后发现,没有肠道菌群的动物表现出更好的葡萄糖耐受性和胰岛素敏感性,而且还会表现出低脂肪以及低体质量指数的情况,研究人员认为,这种代谢结果的改善是由于微生物剔除小鼠机体中嗜酸粒细胞浸润所介导的2型细胞因子信号以及M2巨噬细胞极化作用增强。
Greenblum等[34]揭示了肥胖人群肠道微生物中的厚壁菌门菌群具有磷酸转移酶系统的作用,通过这一系统细菌可将碳水化合物转移至细胞内贮存;肠道碱性磷酸酶(intestinal alkaline phosphatase,IAP)是与肥胖有关的另一种重要酶类,与膳食脂肪的降解有关,IAP的表达可能受肠道微生物控制[35]。在肥胖状态下,肠道微生物还可控制寄主轻微炎症的发生概率[36]。
4.2.2 肥胖人群的肠道菌群差异
近年来的研究结论虽然还存在一定的争议,但大多数研究结果认为肥胖人群与正常人群的肠道微生物的组成差异主要体现在两个细菌门的水平上。Schwiertz等[37]发现,肥胖和超重个体的拟杆菌与厚壁菌门数量比值朝着有利于拟杆菌门的方向改变,粪便总脂肪酸含量显著升高,暗示SCFAs代谢可能与肥胖密切相关。Koleva等[31]总结前人的研究成果后发现,与婴儿体质量状态有关的肠道细菌是拟杆菌和乳杆菌属。Turnbaugh等[38]通过动物模型发现,肥胖与两种细菌门的丰度改变有关;肥胖小鼠相较于正常小鼠肠道中的厚壁菌门丰度增加而拟杆菌门相对减少,这与Hildebrandt等[39]的结论相符。人体实验结果表明,肥胖个体的肠道菌落组成多样性较低,且拟杆菌门数量显著低于非肥胖个体[40-41]。但也有研究人员发现粪便菌群中的拟杆菌与厚壁菌门丰度比例与肥胖没有相关性[42]。Kasai等[43]采用末端限制性酶切片段长度多态性和新一代测序方法,以日本肥胖和非肥胖人群为实验对象,比较二者在肠道微生物组成上的差异,结果表明肥胖个体的细菌多样性显著高于后者,这虽然与Turnbaugh等[40]的研究结果相矛盾,但研究同时发现肥胖个体的拟杆菌门的数量减少,而拟杆菌与厚壁菌门丰度比值降低,结果证实了二者在肠道微生物组成上存在差异,且特定菌种与肥胖具有显著相关性。
4.2.3 益生元对肥胖人群肠道菌群的调节作用
益生元通常为低聚糖、短链多糖或抗性淀粉、膳食纤维等复杂多糖,具有低热量或无热量、耐酸耐消化酶水解、不被小肠吸收等特点,因此可成为蔗糖替代品,是肥胖人群控制体质量的理想选择。膳食补充非消化性碳水化合物能够增加饱腹感和降低食欲[44]、减少能量摄入量和动物体脂肪量[45],起到控制肥胖的作用。近年来的研究表明,肥胖与肠道微生物构成息息相关,尤其与拟杆菌门和厚壁菌门二者间适合的比例有关[46-47],通过膳食性补充益生元改善肥胖人群营养代谢和肠道微生物数量已逐渐成为热点。抗性淀粉作为少数被认可的多糖类益生元,已有大量研究证实,膳食补充抗性淀粉可增加粪便中的双歧杆菌数量[22,37],同时减少厚壁菌门而增加拟杆菌门数量[37]。Cani等[48]发现膳食补充低聚果糖能够增加肥胖小鼠肠道内的双歧杆菌、乳杆菌数量以及内源性GLP-2的生成量,同时降低血浆LPS和细胞因子水平、减少炎症和氧化胁迫因子的表达量;此项研究证实了GLP-2的生成量与特定菌群数量的变化能够改善肥胖患者的肠道屏障功能。
Dewulf等[49]通过双盲干预实验,研究菊粉型果聚糖(inulin-type fructans,ITF)对肥胖女性肠道微生物组成的影响,结果表明,IFT能够增加双歧杆菌和普氏粪杆菌属数量,两种细菌数量与血清LPS水平呈负相关,而血清中LPS水平被认为与肥胖有关[50-51];同时,补充IFT能够减少拟杆菌和丙酸杆菌属数量。Salazar等[52]针对性地研究IFT对肥胖女性粪便中双歧杆菌的影响,结果表明IFT可选择性地增加长双歧杆菌、假小链双歧杆菌和青春双歧杆菌的数量,其中长双歧杆菌数量与血清LPS水平呈负相关;IFT还可减少肥胖女性粪便中的总SCFAs含量,降低与高水平SCFAs相关的代谢风险因子。
4.3 益生元与老年人
4.3.1 老年人肠道菌群变化特点
肠道微生物在人体青壮年时期可保持稳态,然而到了中老年阶段,由于衰老导致的肠道形态学和生理状态的改变使肠道机能下降、蠕动时间延长,pH值升高至7.0~7.5的弱碱性状态[53],加上营养状况、生活方式等外部因素的影响,微生物的数量和多样性开始发生根本性变化。尽管存在个体差异,但是Mariat等[54]认为老年人肠道微生物最显著的变化特征也是拟杆菌与厚壁菌门丰度相对比例的改变,研究发现老年人肠道菌群中拟杆菌门的比例更高,而年轻人的厚壁菌门则相对较高;也有报道指出老年人肠道中的双歧杆菌和梭状芽孢杆菌Ⅳ数量显著下降[55-57],而与肠道疾病相关的肠杆菌科和变形菌门特定菌群的数量有所增加[58]。Claesson等[59]通过对老年人肠道微生物的组成、变异性和稳定性进行系统性分析后发现,拟杆菌门在老年人肠道中比例更大,梭状芽孢杆菌IV丰度增加;大多数肠道菌群在有限的时间内保持稳定,只有个别稀有菌群表现出极强的变异性。
4.3.2 肠道菌群及活力变化对老年人健康的影响
老年人肠道菌群的多样性呈现逐渐下降和结构失衡的趋势,通常情况下,双歧杆菌、乳酸杆菌等益生菌通过产生具有抗炎活力的SCFAs,减少溃疡性结肠炎、克罗恩病等炎症性肠道疾病的发生,而老年人肠道中二者数量与健康成人相比显著下降,直接导致SCFAs生成量减少,这还可能导致肠道上皮细胞分泌黏蛋白进而增加病原菌侵入肠道黏膜的风险[60]。另一方面,肠杆菌等革兰氏阴性菌的丰度增加,这些细菌具有分泌LPS的能力,LPS作为内毒素可导致肠道炎症。此外,由于非消化性成分在老年人小肠中的蠕动时间延长,在大肠中的滞留时间相对缩短,这种平衡的改变可能影响大肠中主要菌群的代谢活力。研究发现,老人粪便中的BCFAs、氨、苯酚以及吲哚等腐败产物浓度增加,这表明老年人大肠中的微生物代谢活性从糖化菌发酵模式转变为对人体不利的致腐微生物代谢类型[58]。肠道微生物代谢活性的转变可导致肠道内容物pH值升高,增溶作用和矿物质利用率降低,增加肠道感染的风险,同时,腐败发酵物大量产生可能增加老年人患结肠癌的风险。
4.3.3 益生元对老年人肠道菌群的影响
大量研究表明,通过膳食补充益生元,能够改善老年人的肠道环境,促进肠胃蠕动,增加排便频率,防止便秘。这一方面是源于益生元在小肠中的不可消化性,其进入大肠被益生菌群选择性发酵,促进益生菌增殖,增加SCFAs含量使得粪便体积膨胀;另一方面,补充益生元能够改善糖化菌群活力退化,减少致腐发酵作用,降低肠道pH值和提高增溶作用。有证据显示,膳食补充益生元或与益生菌复配剂可提高老年人肠道中的内源性益生菌数量。动物模型显示,膳食补充高直链玉米抗性淀粉能够增加老年小鼠(18~20 月龄)肠道中拟杆菌、双歧杆菌、阿克曼斯卡菌属等菌群的比例,其中双歧杆菌和阿克曼斯卡菌属的比例与寄主高血糖原表达水平、肠道质量等显著相关[61]。王春敏等[62-63]以D-半乳糖制造Wistar大鼠和KM小鼠衰老模型,研究异麦芽低聚糖(isomaltooligosaccharides,IMO)灌胃30 d(0.3 g/(kg·d))对小鼠肠道菌群及免疫能力的影响,结果表明IMO可增加衰老模型组的双歧杆菌数量,降低肠杆菌和肠球菌数量,研究还发现IMO对前者乳酸杆菌数量没有显著影响,但却能显著增加KM小鼠衰老模型的乳酸杆菌数量。
有国外研究报道还指出,健康的老年人(64~79 岁)通过膳食补充不同聚合度的低聚半乳糖混合物,可增加双歧杆菌、乳杆菌、肠球菌数量,同时减少拟杆菌、溶组织梭菌、大肠杆菌和脱硫弧菌等菌群的数量[64]。Ouwehand等[65]将嗜酸乳杆菌NCFM与乳糖醇复配使用,研究其对健康老年人(≥65 岁)肠道菌群和免疫指标的影响,二者共同干预下的结果表明,老人排便频率增加,其中双歧杆菌数量增高至7.8×109CFU/g,极显著高于对照组(P=0.001),这接近于健康成年人水平;而产气荚膜梭菌和细菌总数没有显著性变化。Björklund等[66]采取相同的干预措施研究其对使用非甾体抗炎药老人(≥65 岁)肠道微生物的影响,研究发现二者复配使用可增加内源性乳杆菌和双歧杆菌数量,抑制由于使用非甾体抗炎药所导致的球形梭菌-直肠真杆菌和梭状芽孢杆菌XIVab数量下降的趋势。
5 结 语
目前,抗生素被广泛应用于临床治疗肠道感染,通常具有立竿见影的效果,然而抗生素一般具有广谱性,对肠道有益菌同样也有负面作用;长期使用抗生素造成菌株变异所产生耐药性也是公众关心的问题。益生元是除抗生素和益生菌外比较受认可的对肠道菌群有调节作用的天然膳食性成分[67]。益生元的安全性和耐受性使其能够广泛应用于婴幼儿配方乳粉,减少或代替抗生素对新生儿肠道菌群的负面影响;肥胖已由西方发达国家扩展成为世界性的公共卫生问题,目前临床干预肥胖措施主要基于特定肥胖类型、饮食习惯、胰岛素抵抗性以及并发疾病进行治疗方案设计,具体治疗方式包括药物治疗、营养和行为干预[36]。然而大多数治疗干预措施往往收效甚微。近期研究证实益生元干预对肥胖个体的葡萄糖、脂质代谢和改善炎症具有有益影响,而这些代谢指标的改善又与以厚壁菌和拟杆菌门为主的菌群数量改变紧密相关。因此通过膳食补充益生元的营养干预措施,为改善与肥胖相关的肠道微生物组成与代谢紊乱、预防和治疗肥胖提供了新的思路。老年阶段的肠道菌群是人生中除婴儿期外的另一个不稳定时期,与婴儿期微生物定植的不确定性不同,老年时期的肠道菌群处于逐渐退化的状态,菌群的多样性降低且结构失衡。由此引发的肠道炎症性疾病将极大地提高老年人的患病概率,但通过有目的性地膳食性补充益生元是老年人保持肠道菌群结构稳定、防止特定菌种退化、减少抗生素等药物干预的理想选择。
基于寄主-微生物间的相互作用关系,通过膳食补充益生元为微生物调节肠道健康的策略提供了新的途径。然而,目前国内有关益生元对肠道菌群调节作用研究大多都集中于双歧杆菌、乳酸菌这两种特定益生菌以及肠杆(球)菌等常见条件性致病菌的研究。虽然益生元对肠道菌群的调节作用很大程度是由于选择性地促进了双歧杆菌与乳杆菌的增殖与活力,增加了乙酸和乳酸生成量。但是,丙酸和丁酸的生产者通常是梭状芽孢杆菌ⅪⅤa、Ⅳ,拟杆菌属以及Negativicutes纲等细菌[68-69]。因此,为了解益生元对于肠道菌群的调节作用,还应尽可能系统性地研究全部肠道菌群的总体变化,如某些特定菌群比例的变化规律。由于个体性差异,对膳食性益生元的补充还应依据不同地区人群的肠道微生物组成特点选择适合的益生元类型和干预剂量。
[1] WESTERBEEK E A M, VAN DEN BERG A, LAFEBER H N, et al.The intestinal bacterial colonisation in preterm infants: a review of the literature[J]. Clinical Nutrition, 2006, 25(3): 361-368. DOI:10.1016/j.clnu.2006.03.002.
[2] DORÉ J, SIMRÉN M, BUTTLE L, et al. Hot topics in gut microbiota[J]. United European Gastroenterology Journal, 2013, 1(5):311-318. DOI:10.1177/2050640613502477.
[3] EVANS J M, MORRIS L S, MARCHESI J R. The gut microbiome:the role of a virtual organ in the endocrinology of the host[J]. Journal of Endocrinology, 2013, 218(3): 37-47. DOI:10.1530/JOE-13-0131.
[4] COYTE K Z, SCHLUTER J, FOSTER K R. The ecology of the microbiome: networks, competition, and stability[J]. Science, 2015,350: 663-666. DOI:10.1126/science.aad2602.
[5] GIBSON G R, ROBERFROID M B. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics[J]. Journal of Nutrition, 1995, 125(6): 1401-1412.
[6] BINDELS L B, DELZENNE N M, CANI P D, et al. Towards a more comprehensive concept for prebiotics[J]. Nature Reviews Gastroenterology & Hepatology, 2015, 12(5): 303-310. DOI:10.1038/nrgastro.2015.47.
[7] BÄCKHED F, LEY R E, SONNENBURG J L, et al. Host-bacterial mutualism in the human intestine[J]. Science, 2005, 307: 1915-1920.DOI:10.1126/science.1104816.
[8] CILIEBORG M S, BOYE M, SANGILD P T. Bacterial colonization and gut development in preterm neonates[J].Early Human Development, 2012, 88(1): 41-49. DOI:10.1016/j.earlhumdev.2011.12.027.
[9] BÄCKHED F, ROSWALL J, PENG Y Q, et al. Dynamics and stabilization of the human gut microbiome during the first year of life[J]. Cell Host & Microbe, 2015, 17(5): 690-703. DOI:10.1016/j.chom.2015.04.004.
[10] DUNCAN S H, LOUIS P, FLINT H J. Cultivable bacterial diversity from the human colon[J]. Letters in Applied Microbiology, 2007,44(4): 343-350. DOI:10.1111/j.1472-765X.2007.02129.x.
[11] MARTÍNEZ I, MULLER C E, WALTER J. Long-term temporal analysis of the human fecal microbiota revealed a stable core of dominant bacterial species[J]. PLoS ONE, 2013, 8(7): e69621.DOI:10.1371/journal.pone.0069621.
[12] GOEDERT J J, HUA X, YU G Q, et al. Diversity and composition of the adult fecal microbiome associated with history of cesarean birth or appendectomy: analysis of the American Gut Project[J]. EBioMedicine,2014, 1(2): 167-172. DOI:10.1016/j.ebiom.2014. 11.004.
[13] MOUNTZOURIS K C, MCCARTNEY A L, GIBSON G R. Intestinal microflora of human infants and current trends for its nutritional modulation[J]. British Journal of Nutrition, 2002, 87(5): 405-420.DOI:10.1079/BJNBJN2002563.
[14] POKUSAEVA K, FITZGERALD G F, VAN SINDEREN D.Carbohydrate metabolism in Bifidobacteria[J]. Genes & Nutrition,2011, 6(3): 285-306. DOI:10.1007/s12263-010-0206-6.
[15] LOUIS P, HOLD G L, FLINT H J. The gut microbiota, bacterial metabolites and colorectal cancer[J]. Nature Reviews Microbiology,2014, 12(10): 661-672. DOI:10.1038/nrmicro3344.
[16] SCOTT K P, GRATZ S W, SHERIDAN P O, et al. The inf l uence of diet on the gut microbiota[J]. Pharmacological Research, 2013, 69(1):52-60. DOI:10.1016/j.phrs.2012.10.020.
[17] WU G D, CHEN J, HOFFMANN C, et al. Linking long-term dietary patterns with gut microbial enterotypes[J]. Science, 2011, 334: 105-108. DOI:10.1126/science.1208344.
[18] CONLON M A, BIRD A R. The impact of diet and lifestyle on gut microbiota and human health[J]. Nutrients, 2015, 7(1): 17-44.DOI:10.3390/nu7010017.
[19] SERINO M, LUCHE E, GRES S, et al. Metabolic adaptation to a high-fat diet is associated with a change in the gut microbiota[J]. Gut,2012, 61(4): 543-553. DOI:10.1136/gutjnl-2011-301012.
[20] BRINKWORTH G D, NOAKES M, CLIFTON P M, et al.Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations[J]. British Journal of Nutrition, 2009, 101(10): 1493-1502. DOI:10.1017/S0007114508094658.
[21] NEYRINCK A M, POSSEMIERS S, DRUART C, et al. Prebiotic effects of wheat arabinoxylan related to the increase in bif i dobacteria,roseburia and bacteroides/prevotella in diet-induced obese mice[J].PLoS ONE, 2011, 6(6): 230-234. DOI:10.1371/journal.pone.0020944.
[22] 王志凡, 杨秀琳, 陈旺盛, 等. 抗性淀粉对饮食诱导肥胖大鼠排便状况及肠道菌群的影响[J]. 动物营养学报, 2016, 28(5): 1626-1632.DOI:10.3969/j.issn.1006-267X.2016.05.041.
[23] ANITHA M, REICHARDT F, TABATABAVAKILI S, et al. Intestinal dysbiosis contributes to the delayed gastrointestinal transit in highfat diet fed mice[J]. Cellular and Molecular Gastroenterology and Hepatology, 2016, 2(3): 328-339. DOI:10.1016/j.jcmgh.2015.12.008.
[24] HE Yingying, LIU Shubai, SERENA L, et al. Human colostrum oligosaccharides modulate major immunologic pathways of immature human intestine[J]. Mucosal Immunology, 2014, 7(6): 1326-1339.DOI:10.1038/mi.2014.20.
[25] NEU J. Gastrointestinal development and meeting the nutritional needs of premature infants[J]. American Journal of Clinical Nutrition, 2007,85(2): 629S-634S.
[26] HUNTER C J, UPPERMAN J S, FORD H R, et al. Understanding the susceptibility of the premature infant to necrotizing enterocolitis(NEC)[J]. Pediatric Research, 2008, 63(2): 117-123. DOI:10.1203/PDR.0b013e31815ed64c.
[27] RIPOLL C, CHAPPUIS E, RESPONDEK F, et al. scFOS supplemented follow-on formula in healthy infants: impact on vaccine specif i c faecal secretory IGA response, faecal bif i dobacteria, growth and digestive tolerance[J]. Bioactive Carbohydrates and Dietary Fibre,2015, 5(2): 169-178. DOI:10.1016/j.bcdf.2015.03.006.
[28] WESTERBEEK E A, SLUMP R A, LAFEBER H N, et al. The effect of enteral supplementation of specif i c neutral and acidic oligosaccharides on the faecal microbiota and intestinal microenvironment in preterm infants[J]. European Journal of Clinical Microbiology and Infectious Diseases, 2013, 32(2): 269-276. DOI:10.1007/s10096-012-1739-y.
[29] MODI N, UTHAYA S, FELL J, et al. A randomized, double-blind,controlled trial of the effect of prebiotic oligosaccharides on enteral tolerance in preterm infants (ISRCTN77444690)[J]. Pediatric Research,2010, 68(5): 440-445. DOI:10.1203/PDR.0b013e3181f1cd59.
[30] UNDERWOOD M A, GAERLAN S, DE LEOZ M L A, et al. Human milk oligosaccharides in premature infants: absorption, excretion, and influence on the intestinal microbiota[J]. Pediatric Research, 2015,78(6): 670-677. DOI:10.1038/pr.2015.162.
[31] KOLEVA P T, BRIDGMAN S L, KOZYRSKYJ A L. The infant gut microbiome: evidence for obesity risk and dietary intervention[J].Nutrients, 2015, 7(4): 2237-2260. DOI:10.3390/nu7042237.
[32] RIDAURA V K, FAITH J J, REY F E, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice[J]. Science,2013, 341: 1241214. DOI:10.1126/science.1241214.
[33] SUÁREZ-ZAMORANO N, FABBIANO S, CHEVALIER C, et al.Microbiota depletion promotes browning of white adipose tissue and reduces obesity[J]. Nature Medicine, 2015, 21(12): 1497-1501.DOI:10.1038/nm.3994.
[34] GREENBLUM S, TURNBAUGH P J, BORENSTEIN E.Metagenomic systems biology of the human gut microbiome reveals topological shifts associated with obesity and inflammatory bowel disease[J]. Proceedings of the National Academy of Sciences, 2011,109(2): 594-599. DOI:10.1073/pnas.1116053109.
[35] BATES J M, AKERLUND J, MITTGE E, et al. Intestinal alkaline phosphatase detoxif i es lipopolysaccharide and prevents inf l ammation in zebraf i sh in response to the gut microbiota[J]. Cell Host & Microbe,2007, 2(6): 371-382. DOI:10.1016/j.chom.2007.10.010.
[36] EVERARD A, CANI P D. Diabetes, obesity and gut microbiota[J].Best Practice & Research Clinical Gastroenterology, 2013, 27(1):73-83. DOI:10.1016/j.bpg.2013.03.007.
[37] SCHWIERTZ A, TARAS D, SCHÄFER K, et al. Microbiota and SCFA in lean and overweight healthy subjects[J]. Obesity, 2010, 18(1):190-195. DOI:10.1038/oby.2009.167.
[38] TURNBAUGH P J, LEY R E, MAHOWALD M A, et al. An obesityassociated gut microbiome with increased capacity for energy harvest[J]. Nature, 2006, 444: 1027-1031. DOI:10.1038/nature05414.
[39] HILDEBRANDT M A, HOFFMANN C, SHERRILL-MIX S A,et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity[J]. Gastroenterology, 2009,137(5): 1716-1724. DOI:10.1053/j.gastro.2009.08.042.
[40] TURNBAUGH P J, HAMADY M, YATSUNENKO T, et al. A core gut microbiome in obese and lean twins[J]. Nature, 2009, 457: 480-484.DOI:10.1038/nature07540.
[41] ARMOUGOM F, HENRY M, VIALETTES B, et al. Monitoring bacterial community of human gut microbiota reveals an increase in lactobacillus in obese patients and methanogens in anorexic patients[J]. PLoS ONE,2009, 4(9): e7125. DOI:10.1371/journal.pone.0007125.
[42] DUNCAN S H, LOBLEY G E, HOLTROP G, et al. Human colonic microbiota associated with diet, obesity and weight loss[J]. International Journal of Obesity, 2008, 32(11): 1720-1724.DOI:10.1038/ijo.2008.155.
[43] KASAI C, SUGIMOTO K, MORITANI I, et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing[J]. BMC Gastroenterology,2015, 15(1): 1-10. DOI:10.1186/s12876-015-0330-2.
[44] ARCHER B J, JOHNSON S K, DEVEREUX H M, et al. Effect of fat replacement by inulin or lupin-kernel fibre on sausage patty acceptability, post-meal perceptions of satiety and food intake in men[J]. British Journal of Nutrition, 2004, 91(4): 591-599.DOI:10.1079/BJN20031088.
[45] BOULANGÉ C L, NEVES A L, CHILLOUX J, et al. Impact of the gut microbiota on inf l ammation, obesity, and metabolic disease[J]. Genome Medicine, 2016, 8(1): 1-12. DOI:10.1186/s13073-016-0303-2.
[46] LEY R E. Obesity and the human microbiome[J]. Current Opinion in Gastroenterology, 2010, 26(1): 5-11. DOI:10.1097/MOG.0b013e328333d751.
[47] LEY R E, TURNBAUGH P J, KLEIN S, et al. Microbial ecology:human gut microbes associated with obesity[J]. Nature, 2006, 444:1022-1023. DOI:10.1038/4441022a.
[48] CANI P D, POSSEMIERS S, VAN DE WIELE T, et al. Changes in gut microbiota control inf l ammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability[J]. Gut,2009, 58(8): 1091-1103. DOI:10.1136/gut.2008.165886.
[49] DEWULF E M, CANI P D, CLAUS S P, et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women[J]. Gut, 2013, 62(8):1112-1121. DOI:10.1136/gutjnl-2012-303304.
[50] AMAR J, CHABO C, WAGET A, et al. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: molecular mechanisms and probiotic treatment[J].EMBO Molecular Medicine, 2011, 3(9): 559-572. DOI:10.1002/emmm.201100159.
[51] AMAR J, SERINO M, LANGE C, et al. Involvement of tissue bacteria in the onset of diabetes in humans: evidence for a concept[J]. Diabetologia,2011, 54(12): 3055-3061. DOI:10.1007/s00125-011-2329-8.
[52] SALAZAR N, DEWULF E M, NEYRINCK A M, et al. Inulin-type fructans modulate intestinal Bif i dobacterium species populations and decrease fecal short-chain fatty acids in obese women[J]. Clinical Nutrition, 2015, 34(3): 501-507. DOI:10.1016/j.clnu.2014.06.001.
[53] BARCZYNSKA R, BANDURSKA K, SLIZEWSKA K, et al.Intestinal microbiota, obesity and prebiotics[J]. Polish Journal of Microbiology, 2015, 64(2): 93-100.
[54] MARIAT D, FIRMESSE O, LEVENEZ F, et al. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age[J]. BMC Microbiology, 2009, 9(1): 1-6. DOI:10.1186/1471-2180-9-123.
[55] HOPKINS M J, SHARP R, MACFARLANE G. Age and disease related changes in intestinal bacterial populations assessed by cell culture, 16S rRNA abundance, and community cellular fatty acid prof i les[J]. Gut, 2001, 48(2): 198-205. DOI:10.1136/gut.48.2.198.
[56] ZWIELEHNER J, LISZT K, HANDSCHUR M, et al. Combined PCR-DGGE fingerprinting and quantitative-PCR indicates shifts in fecal population sizes and diversity of Bacteroides, bifidobacteria and Clostridium cluster IV in institutionalized elderly[J].Experimental Gerontology, 2009, 44(6/7): 440-446. DOI:10.1016/j.exger.2009.04.002.
[57] GAVINI F, CAYUELA C, ANTOINE J M, et al. Differences in the distribution of bifidobacterial and enterobacterial species in human faecal microflora of three different (children, adults, elderly) age groups[J]. Microbial Ecology in Health and Disease, 2001, 13(1):40-45. DOI:10.3402/mehd.v13i1.7998.
[58] WOODMANSEY E J. Intestinal bacteria and ageing[J]. Journal of Applied Microbiology, 2007, 102(5): 1178-1186. DOI:10.1111/j.1365-2672.2007.03400.x.
[59] CLAESSON M J, CUSACK S, O’SULLIVAN O, et al. Composition,variability, and temporal stability of the intestinal microbiota of the elderly[J]. Proceedings of the National Academy of Sciences, 2011,108(Suppl 1): 4586-4591. DOI:10.1073/pnas.1000097107.
[60] SCHIFFRIN E J, MORLEY J E, DONNET-HUGHES A,et al. The inflammatory status of the elderly: the intestinal contribution[J]. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, 2010, 690(1/2): 50-56. DOI:10.1016/j.mrfmmm.2009.07.011.
[61] SYBILLE T, ZHOU J, MICHAEL K, et al. The intestinal microbiota in aged mice is modulated by dietary resistant starch and correlated with improvements in host responses[J]. FEMS Microbiology Ecology,2013, 83(2): 299-309. DOI:10.1111/j.1574-6941.2012.01475.x.
[62] 王春敏, 李丽秋, 任继秋, 等. 异麦芽低聚糖对D-半乳糖致衰老大鼠肠道菌群、血清IgG和肠黏膜sIgA的影响[J]. 中国微生态学杂志,2009, 21(1): 7-8; 12. DOI:1005-376X (2009)01-0007-02.
[63] 王春敏, 马淑霞, 佟丽波, 等. 异麦芽低聚糖对衰老模型小鼠肠黏膜免疫功能调节作用的研究[J]. 中国微生态学杂志, 2012, 24(1): 6-8;12. DOI:1005-376X(2012)01-0006-03.
[64] VULEVIC J, DRAKOULARAKOU A, YAQOOB P, et al. Modulation of the fecal microflora profile and immune function by a novel trans-galactooligosaccharide mixture (B-GOS) in healthy elderly volunteers[J]. The American Journal of Clinical Nutrition, 2008, 88(5):1438-1446.
[65] OUWEHAND A C, TIIHONEN K, SAARINEN M, et al. Influence of a combination of Lactobacillus acidophilus NCFM and lactitol on healthy elderly: intestinal and immune parameters[J]. British Journal of Nutrition, 2009, 101(3): 367-375. DOI:10.1017/S0007114508003097.
[66] BJÖRKLUND M, OUWEHAND A C, FORSSTEN S D, et al. Gut microbiota of healthy elderly NSAID users is selectively modif i ed with the administration of Lactobacillus acidophilus NCFM and lactitol[J].Age, 2012, 34(4): 987-999. DOI:10.1007/s11357-011-9294-5.
[67] TIIHONEN K, OUWEHAND A C, RAUTONEN N. Human intestinal microbiota and healthy ageing[J]. Ageing Research Reviews, 2010,9(2): 107-116. DOI:10.1016/j.arr.2009.10.004.
[68] LOUIS P, YOUNG P, HOLTROP G, et al. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene[J]. Environmental Microbiology, 2010,12(2): 304-314. DOI:10.1111/j.1462-2920.2009.02066.x.
[69] REICHARDT N, DUNCAN S H, YOUNG P, et al. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota[J]. Isme Journal, 2014, 8(6): 1323-1335.DOI:10.1038/ismej.2014.14.
Advances in Intestinal Flora Regulatory Mechanisms of Prebiotics in Premature Infants,Obesity Subjects and Elderly Populations
ZHENG Zhichang, CHEN Yingtong, GUO Juanjuan, GUO Zebin, ZHENG Baodong, LU Xu*
(College of Food Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China)
The intestinal microflora is an indispensable part of the human body. Intestinal microbial diversity and the stability of microbial community structure are of great importance for human health. Clinical studies have shown that intestinal microbial populations play an important role in the development of disease for special populations. Prebiotics, as an important part of human diet, are mainly derived from non-digestible oligosaccharides and dietary fi ber from plants. It has been demonstrated that prebiotics could selectively promote intestinal microf l ora proliferation and enhance metabolic activity. This paper is focused on the differences in intestinal microbial colonization among special populations including premature infants, obese people and the elderly based on the intestinal microbiota changes with age, as well as proposed mechanisms of prebiotics for regulating the the intestinal microbiota in the populations. We hope this review can provide a theoretical guideline for further research on the mechanism underlying prebiotic regulation of the intestinal microbiota and the development of prebiotic functional foods.
prebiotics; intestinal fl ora; premature infants; obesity; the elderly
10.7506/spkx1002-6630-201801045
TS201.4
A
1002-6630(2018)01-0297-08
郑志昌, 陈映彤, 郭娟娟, 等. 益生元对早产儿、肥胖及老年群体肠道菌群调节机制的研究现状[J]. 食品科学, 2018,39(1): 297-304.
10.7506/spkx1002-6630-201801045. http://www.spkx.net.cn
ZHENG Zhichang, CHEN Yingtong, GUO Juanjuan, et al. Advances in intestinal fl ora regulatory mechanisms of prebiotics in premature infants, obesity subjects and elderly populations[J]. Food Science, 2018, 39(1): 297-304. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201801045. http://www.spkx.net.cn
2016-10-05
国家自然科学基金青年科学基金项目(31601473);福建省省属高校专项基金项目(JK2015012);
福建省高等学校科技创新团队支持计划项目(闽教科[2012]03号);福建农林大学高水平大学建设项目(61201404227);
福建农林大学科技发展资金项目(KF2015099)
郑志昌(1992—),男,硕士研究生,研究方向为食品化学与营养。E-mail:1792124420@qq.com
*通信作者简介:卢旭(1986—),男,讲师,博士,研究方向为碳水化合物。E-mail:lxvfst@yeah.net
