尼奥罗非鱼杂种性别类型与性腺发育初步研究
2017-12-18赵永华赵金良陈晓武魏继海伞桂宝简伟业
赵永华,赵金良,陈晓武,王 燕,魏继海,伞桂宝,简伟业
( 1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306; 2.广州市番禺区农业科学研究所,广东 广州 511450; 3.茂名市伟业罗非鱼良种场,广东 茂名 525000 )
尼奥罗非鱼杂种性别类型与性腺发育初步研究
赵永华1,赵金良1,陈晓武1,王 燕1,魏继海1,伞桂宝2,简伟业3
( 1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306; 2.广州市番禺区农业科学研究所,广东 广州 511450; 3.茂名市伟业罗非鱼良种场,广东 茂名 525000 )
通过组织学方法,研究了0.5~4.5月龄尼奥罗非鱼杂种(尼罗罗非鱼♀×奥利亚罗非鱼♂)的性腺发育特征与性别类型。结果表明,0.5月龄尼奥罗非鱼原始性腺尚未分化;1月龄,精巢中原始生殖细胞开始分化成精原细胞;2.5月龄出现初级精母细胞;3月龄出现次级精母细胞;3.5~4月龄精巢中存在不同发育阶段的生殖细胞,壶腹空腔中分散着稀疏的精子;4.5月龄精巢壶腹腔内密集大量精子。在2.5、3.5、4、4.5月龄抽样中,均检测到雌鱼,2.5~3.5月龄卵巢以第Ⅱ时相卵母细胞为主;4~4.5月龄卵巢以第Ⅲ时相卵母细胞为主。研究结果初步表明,尼奥罗非鱼杂种中观察到雄性、雌性两种性别类型,以雄性性别为主;杂种雌、雄鱼的性腺均能正常发育。
尼奥罗非鱼;性别类型;性腺发育;组织切片
罗非鱼(Oreochromis)因其具有食性杂、生长快、繁殖力和抗病力强等优点,现已成为世界性养殖鱼类[1]。由于罗非鱼雄性比雌性生长速度快,为此,生产上一般采用雌性尼罗罗非鱼(O.niloticus)和雄性奥利亚罗非鱼(O.aureus)种间杂交的方式获得雄性率高的杂交一代——尼奥罗非鱼[2]。
有关尼奥罗非鱼雄性率高的性别形成机制,早期认为,尼罗罗非鱼是雄性异质型性染色体,雄性由异质的XY性染色体决定,雌性由同质的XX性染色体决定[3];奥利亚罗非鱼是雌性异质型性染色体,雄性由同质的ZZ性染色体决定,雌性由异质的ZW性染色体决定[4],因此,杂交一代为XZ型雄性。除性染色体外,Avtalion等[5]提出,一条常染色体上的一对等位基因(AA或aa)影响性染色体,尼罗罗非鱼雌性为AAXX,雄性为AAXY;奥利亚罗非鱼雌性为aaZW,雄性为aaZZ,这个基因平衡模型假设,当性别决定有关的染色体和常染色体位点的作用超过某一阈值时,个体发育为雄性,低于这一阈值时则为雌性。
目前研究表明,大多数鱼类不存在性染色体,性腺分化和发育受相关基因共同调控。罗非鱼中已发现与性别分化相关的基因有很多,且定位于不同的染色体上,如尼罗罗非鱼位于LG12染色体上的dmrt1基因,LG23染色体上的amh基因,LG1染色体上的cyp19a1基因,以及一些未成功定位的基因,这些单个或者多个基因共同参与罗非鱼性别决定与分化调控[6]。此外,环境因子如温度[7]、类固醇激素[8]等对罗非鱼性别分化和发育也都有显著影响。
已有的研究表明,尼罗罗非鱼早期性腺发育中出现两性性腺现象[9],也有学者提出雌性致死假设观点,为进一步探明尼奥罗非鱼杂交种性别偏离及性腺发育,本研究通过对0.5~4.5月龄尼奥罗非鱼性腺组织学特征连续观察,以期揭示尼奥罗非鱼的性别类型、性腺发育特征。
1 材料与方法
尼奥罗非鱼鱼苗2015年5月28日采自广州市番禺区农业科学研究所良种繁殖场,于上海海洋大学罗非鱼种质资源试验站的7.8 m×3.9 m×1.5 m水泥池中养殖,定时足量投喂,每个水泥池放养500尾,用罗非鱼专用配合饲料养殖。
每隔15 d采样一次,每次随机采集30~40尾。常规生物学测量体长、体质量。幼鱼前期,性腺细小无法剥取,截取性腺所在躯干部位,波恩氏液固定;当性腺可以剥取时,取完整性腺固定。组织经石蜡包埋,制成5~8 μm切片,苏木精—伊红染色,中性树胶封片,于显微镜(Nikon DS-Ril)观察拍照。用目微尺测量性腺横截面的宽度和厚度以及生殖细胞的大小、核径。卵巢、精巢的发育分期参照刘筠[10]分期标准。
试验所得数据用Excel 2013软件进行统计学分析,结果以平均值±标准差表示。
2 结 果
2.1 尼奥罗非鱼的生长
0.5~4月龄尼奥罗非鱼体长与体质量随月龄的增加而显著增加,体质量增长量随月龄的增加而增大(表1)。

表1 尼奥罗非鱼生长
2.2 尼奥罗非鱼性腺发育
在9次采样中,绝大多数为雄性个体,雌性个体仅在2.5、3.5、4、4.5月龄中检测到,且在这4次采样中分别只采集到1尾雌性个体,雄性率高达98.8%。尼奥罗非鱼性腺大小随生长而增大,同一批次采样的卵巢体积比精巢大(表2)。
2.2.1 性腺未分化期
0.5月龄:性腺细小,肉眼无法辨别,性腺处于原始性腺阶段。在显微镜下可观察到性腺原基上布满原始生殖细胞(3.21~5.15 μm),不能明显区分精巢、卵巢(图1a)。
2.2.2 精巢发育
Ⅰ期精巢(1~1.5月龄):精巢肉眼无法分辨,体积较小,白色。横切面呈椭圆形,生殖细胞核较大,染色较深,细胞质色质较淡。1月龄,原生殖细胞开始分裂为精原细胞(图1b);1.5月龄,精原细胞数目增多,无定向的分散在结缔组织中,精原细胞直径7.25~8.15 μm(图1c)。
Ⅱ期精巢(2月龄):精巢体积增大1倍,呈白色线状。部分单层精原细胞集中排列,形成精细小管,部分有管腔。精细小管大多呈圆形或椭圆形,直径13.19~18.21 μm,其内的精原细胞细胞核位于中央或稍微偏离中心,精原细胞直径6.15~7.94 μm,核径1.6~2.3 μm(图1d)。
Ⅲ期精巢(2.5月龄):精巢成肉色圆柱状,表面有细小血管分布,体积继续增大。精巢中产生数量较多的精原细胞,精小叶内出现空腔,精原细胞、初级精母细胞沿精小叶边缘单层或多层排列,初级精母细胞直径6.32~8.25 μm,核径3.21~4.19 μm。精巢中形成1个大型空腔和1~2个小型空腔(图1e)。
Ⅳ期精巢(3~4月龄):3月龄,精巢肉白色,扁平三棱柱状,表面血管分布明显。精小叶增大,其内可见由初级精母细胞、次级精母细胞及精细胞组成的精小囊,次级精母细胞直径3.14~4.21 μm,核径2.05~3.13 μm,此时小叶腔中已存在少量的精子,小型空腔数有所增加(图1f)。3.5~4月龄,精巢呈肉色或肉白色,精巢背部带有纵横血管。细胞分裂较为明显,部分精子通过精细小管壁经结缔组织向小叶空腔游动,腔内分散着稀疏的精子(图1g,h)。
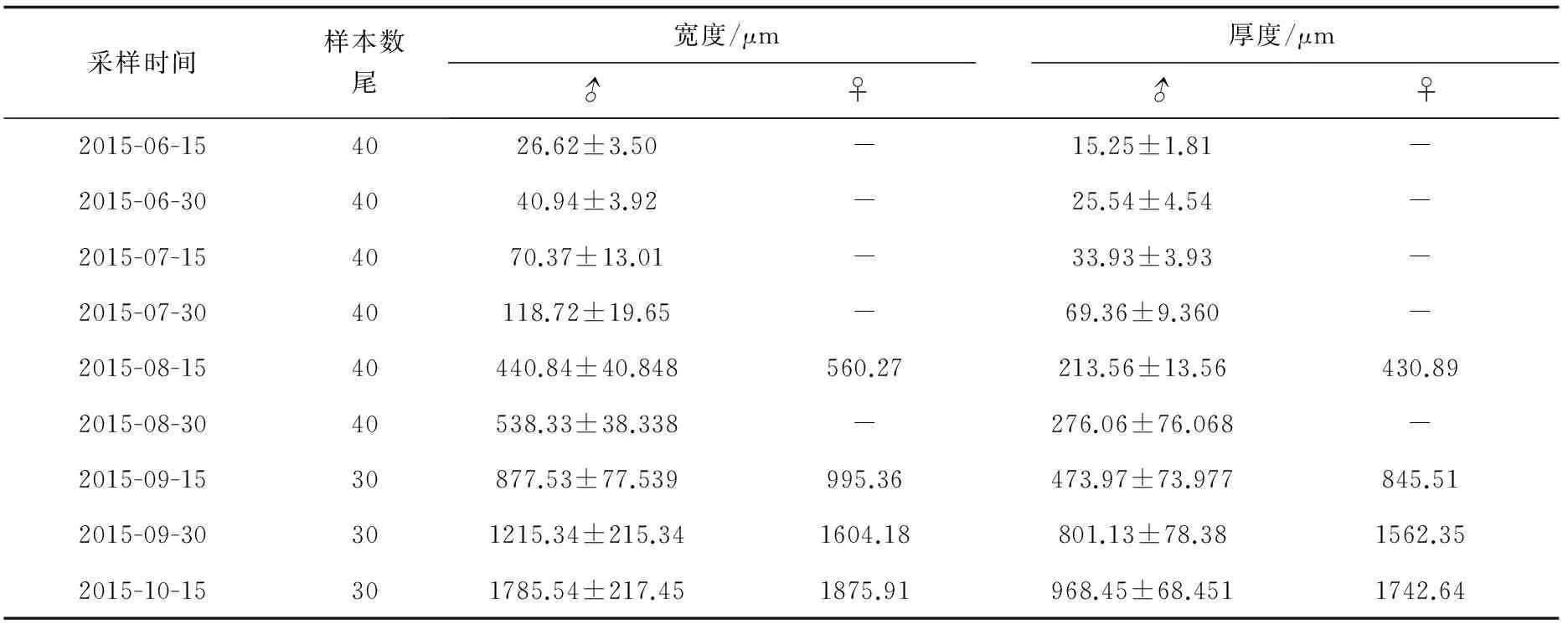
表2 尼奥罗非鱼不同发育阶段性腺宽度和厚度
Ⅴ期精巢(4.5月龄):精巢体积显著增大,呈乳白色,挤压腹部可挤出精子。小叶腔中一些相邻精小囊壁互相融合,小叶空腔很大,其内充满成熟的精子。精子嗜碱性极强,被苏木精染成深蓝色,直径2.1~2.5 μm(图1i)。
2.2.3 卵巢发育
Ⅱ期卵巢(2.5~3.5月龄):2.5月龄,卵巢呈白色细线状,肉眼无法辨认,性腺末端未见血管分布。卵母细胞大小均匀,圆形不规则排列,细胞核居中央,占卵细胞一半左右,卵细胞直径36.18~44.07 μm,核径16.79~22.23 μm,核仁10个以上,不规则地分散在细胞核中,卵细胞外具有一层滤泡膜,以第Ⅱ时相卵母细胞为主(图1j)。在3.5月龄卵巢中,卵母细胞直径33.18~57.07 μm,核径17.83~26.15 μm,出现少量卵黄核(图1k)。
Ⅲ期卵巢(4~4.5月龄):4月龄,卵巢增大,呈囊状,表面可见血管,肉眼已能够看清细小的卵粒。卵母细胞不规则的排列在卵巢基质中,卵径50~110 μm,核径20~50 μm,核仁数增多沿核膜分布,外侧由一层滤泡膜发育成二层滤泡膜,开始出现辐射带,切片中以第Ⅲ时相卵母细胞为主,有少部分第Ⅱ时相卵母细胞(图1l)。4.5月龄,卵巢表面血管增多,卵径为50~200 μm,核径40~95 μm,切片上出现少数Ⅳ时相卵母细胞,辐射带增厚,细胞质中开始出现许多空泡,卵黄颗粒数量和体积增加(图1m)。
3 讨 论
3.1 性腺未分化期
性腺形成的标志是原始生殖细胞的出现。本研究中,0.5月龄尼奥罗非鱼的性腺原基上已布满原始生殖细胞,原始性腺已经形成,尚处于性腺未分化期,此时还不能区分精巢和卵巢,性腺平均宽度为(26.62±3.50) μm,平均厚度为(15.25±1.81) μm,略低于同期的尼罗罗非鱼、莫桑比克罗非鱼(O.mossambicus)性腺大小[11-12]。研究表明,孵化后10日龄,尼罗罗非鱼性腺原基分化明显,至15日龄,原始性腺已形成[11];莫桑比克罗非鱼在孵化后10~11日龄,性腺原基分化明显,生殖细胞数目迅速增加,逐渐形成原始性腺[12]。这些表明,尼奥罗非鱼原始性腺形成与其他罗非鱼较为接近,形成时间约为孵化后10~15 d完成。
3.2 性腺分化期

3.3 性腺分化发育完整性
在尼罗罗非鱼性腺早期发育研究中,刘筠等[9,11]在Ⅰ、Ⅱ期精巢和卵巢中发现有精母细胞和卵母细胞共存的现象。但在本研究中,1~4.5月龄尼奥罗非鱼的精巢(或卵巢)中均未观察到两性细胞共存的现象,性腺明显分化为精巢或卵巢,未发现有“假雄”性别或“中间”性别类型。根据尼奥罗非鱼成熟精巢中存在不同发育阶段的生殖细胞,Ⅱ、Ⅲ期卵巢中同时存在多个时相的卵母细胞,初步认为,尼奥罗非鱼的精巢、卵巢也属于非同步发育型。

图1 尼奥罗非鱼性腺分化和发育
a:0.5月龄原始性腺,×100;b:1月龄Ⅰ期精巢,×100;c:1.5月龄精巢,×100;d:2月龄,Ⅱ期精巢,×40;e:2.5月龄,Ⅲ期精巢,×40;f:3月龄精巢,×40;g:3.5月龄精巢,×40;h:4月龄精巢,×40;i:4.5月龄,Ⅳ期精巢;j:2.5月龄,Ⅱ期卵巢,×40;k:3.5月龄卵巢,×40;l:4月龄,Ⅲ期卵巢,×40;m:4.5月龄,Ⅲ期卵巢,×40;PGCs:原始生殖细胞;SG:精原细胞;PS:初级精母细胞;SPG:精小囊;SS:次级精母细胞;St:精细小管;ST:精子细胞;Pi:壶腹腔;Og:卵原细胞;Nu:核仁;YN:卵黄核;FM:滤泡膜.
本研究结果初步表明,尼奥罗非鱼存在雌、雄两种性别类型,以雄性性别占主导(98%),雄鱼中精原细胞、初级精母细胞、次级精母细胞、精子细胞正常发育,能够产生成熟精子。据此认为,种间杂交对尼奥罗非鱼的性腺分化与生殖细胞发育并未产生明显影响,但杂交对杂种的精子数量、活力影响尚需进一步分析。许玉德等[17]对尼罗罗非鱼、奥利亚罗非鱼及尼奥罗非鱼精巢组织学观察,发现尼奥罗非鱼精巢内生殖细胞密度比亲本略疏,间质组织相对发达,间质细胞数目相对较多。万英杰等[18]对体质量400~500 g尼奥罗非鱼雌鱼解剖观察发现,卵巢分正常型和萎缩型,萎缩型的雌鱼数量约为正常型雌鱼的3倍,由于萎缩型雌鱼不能正常排卵,表现出“雄性化”现象。本研究中,由于雌鱼数量过少,也未能观察到“萎缩型”卵巢。今后,还需扩大样本数量,分析探讨杂交对杂种卵巢后期发育及卵母细胞成熟是否产生影响,理解尼奥罗非鱼杂种雄性率高的生理机制。
在尼奥罗非鱼性腺解剖中也发现,大多数个体的性腺为双侧对称型,也有些个体为双侧不对称型,一侧短小,甚至还出现单侧性腺发育,这类性腺比正常型短小,这种现象是与罗非鱼性腺不同步发育有关,或是由于杂交引起的杂种不育性,还需进一步研究确定。
[1] 张红燕, 袁永明, 贺艳辉, 等. 中国罗非鱼生产与贸易现状分析及建议[J].中国渔业经济,2015,33(3):95-100.
[2] 梁仁杰, 王晓斌. 奥尼鱼苗种雄性率的鉴定与生长测定试验[J]. 水利渔业,2004,24(4):33.
[3] Müller-Belecke A, Hörstgen-Schwark G. Sex determination in tilapia (Oreochromisniloticus): sex ratios in homozygous gynogenetic progeny and their offspring[J]. Aquaculture,1995,137(1):57-65.
[4] Desprez D, Mélard C. Effect of ambient water temperature on sex determinism in the blue tilapia,Oreochromisaureus[J]. Aquaculture, 1998,162(1):79-84.
[5] Avtalion R R, Hammerman I S. Sex-determination inSarotherodon(Tilapia) Ⅰ.Introduction to the theory of autosomal influences[J]. The Israeli Journal of Aquaculture Bamidgeh,1978,30(4):110-115.
[6] 郑尧, 王在照, 陈家长. 调控鱼类性腺分化基因的研究进展[J]. 水生生物学报,2015, 39(4):798-809.
[7] 党广成,刘羽清,付佩胜,等. 高温诱导对尼罗罗非鱼性别分化及生长的影响[J]. 渔业科学进展,2011,32(5):32-37.
[8] Sun L N, Jiang X L, Xie Q P, et al. Transdifferentiation of differentiated ovary into functional testis by long-term treatment of aromatase inhibitor in Nile tilapia[J].Endocrinology,2014,155(4):1476-1488.
[9] 刘筠,刘国安, 陈淑群,等. 尼罗罗非鱼性腺发育的研究[J]. 水生生物学集刊,1983,7(1):17-32.
[10] 刘筠. 鱼类繁殖生物学[M]. 北京:中国农业出版社,1993:23-32.
[11] 王令玲, 仇潜如, 吴福煌. 尼罗罗非鱼性腺发育的研究[J]. 淡水渔业,1986(2):1-4.
[12] 陈玉琳, 胡秀敏, 朱雅珠. 莫桑比克罗非鱼幼鱼的性腺发育与分化[J]. 水产学报,1980,4(4):313-320.
[13] 马学坤, 柳学周, 温海深, 等. 半滑舌鳎性腺分化的组织学观察[J]. 海洋水产研究,2006,27(2):55-61.
[14] 代丽, 刘彬, 王志坚. 稀有鮈鲫性腺分化的组织学观察[J]. 西南师范大学学报,2013,38(1):55-61.
[15] 苏敏, 林丹军, 尤永隆, 等. 黑脊倒刺鲃性腺早期分化的研究[J]. 淡水渔业,2008,38(1):62-66.
[16] 汪学杰, 田媛, 刘奕, 等. 双须骨舌鱼性腺发育的组织学观察[J]. 水产学报,2014,38(4):524-530.
[17] 许玉德, 钟建兴, 郑森林, 等. 单性罗非鱼及其亲本精巢的组织学和LDH同工酶的比较[J]. 厦门大学学报:自然科学版,1999,38(4):628-632.
[18] 万英杰, 周之武. 雄性化奥尼鱼卵巢生物学的初步研究[J]. 水利渔业,1992(2):28-29.
GendersandGonadalDevelopmentinHybridsofNileTilapiaOreochromisniloticus♀×O.aureus♂
ZHAO Yonghua1, ZHAO Jinliang1, CHEN Xiaowu1, WANG Yan1, WEI Jihai1,SAN Guibao2,JIAN Weiye3
( 1. Laboratory of Freshwater Fisheries Germplasm Resources, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China; 2. Guangzhou Panyu Institute of Agricultural Sciences, Guangzhou 511450,China;3. Maoming Weiye Tilapia Seed Farm, Maoming 525000, China )
The gender type and gonadal development of 0.5—0.45 month old Nile tilapiaOreochromisniloticus(♀)×O.aureus(♂) were observed by microscopic histology. The results showed that primordial gonad of the tilapia was undifferentiated at 0.5-month-old.The primordial germ cells in the testis differentiated into spermatogonia around 1 month, primary spermatocytes occurred at the age of 2.5 month, while secondary spermatocytes presented around 3-month-old. The germ cells were present in different developmental stages in the testis and sparse sperm dispersed in ampulla cavity at the age of 3.5-4.5 months,while intensive sperm were dispersed in ampulla cavity around 4.5 months. Among the samples, female individuals were also detected at 2.5,3.5,4 and 4.5 months, respectively,the oocytes were in phase Ⅱat 2.5-3.5 months ovarian,and phase Ⅱat 4 and 4.5 month-old.The findings suggested that there were two kinds of gender in hybrids with the male dominated, and that the testis and ovarian in the hybrids developed normally.
Oreochromisniloticus♀×O.aureus♂;gender type; gonadal development; histological method
10.16378/j.cnki.1003-1111.2017.04.011
S965.125
A
1003-1111(2017)04-0467-05
2016-04-08;
2016-11-28.
国家现代农业产业技术体系专项资金资助项目(CARS-49);水产动物遗传育种中心上海市协同创新中心资助项目(ZF1206);广东省水产良种体系建设专项(2016).
赵永华(1990-),男,硕士研究生;研究方向:水产动物遗传育种与繁殖. E-mail:1099875884@qq.com. 通讯作者:赵金良(1969-),男,教授;研究方向:动物遗传育种与繁殖.E-mail:jlzhao@shou.edu.cn.
