果蝇NOMPC相互作用蛋白的筛选与鉴定
2017-12-14郑红兰郭逸峰闫致强
郑红兰,郭逸峰,闫致强
(复旦大学 生命科学学院,上海 200438)
果蝇NOMPC相互作用蛋白的筛选与鉴定
郑红兰,郭逸峰,闫致强
(复旦大学 生命科学学院,上海 200438)
NOMPC是果蝇感受触觉的机械力敏感离子通道,属于瞬时受体电位(TRP)离子通道家族,机械力敏感离子通道通过怎样的机制被机械力激活?与NOMPC相互作用的蛋白有哪些?为了研究这些问题,本研究从果蝇NOMPC全长cDNA中扩增得到其N端的29个锚蛋白重复序列(ARs),ORF长度为3039bp.然后将这29个ARs构建到表达载体pUAST-EGFP上,转染果蝇S2细胞后,通过免疫沉淀、质谱分析和Western blot筛选和鉴定NOMPC相互作用的蛋白,实验筛选出NOMPC N端的29ARs和NOMPC全长的相互作用蛋白,通过免疫共沉淀(Co-IP)实验,对可能与NOMPC存在相互作用的蛋白质进行验证,鉴定出Annexin B9、CG3195、CG3731、CG5374与NOMPC存在相互作用,为进一步研究NOMPC介导触觉转导的分子机制提供了基础.
果蝇; 触觉; NOMPC; 结合蛋白
为了满足生存、繁衍等根本的需求,生物体将会感知外界的各种类型的刺激,进而做出相应的反应.至今,经过了漫长的进化,动物拥有了各种复杂的感觉器官.上世纪八十年代,Nathans等克隆了介导视觉转导的视紫红质(Rhodopsin)基因[1];在上个世纪九十年代初,Buck等克隆了介导嗅觉的嗅觉受体基因[2];在上个世纪九十年代末到本世纪初,Hoon等发现了介导味觉的受体[3].但是触觉和听觉的分子转导机制一直都不清楚,同样这也是一个重要的科学问题.其中触觉对于环境探测和社会交往是必不可少的.然而,触觉形成的基本机制大部分仍然是未知的.
机械力感觉是指生物体将体内体外的刺激转化成电信号,传入到神经系统后引起的一系列的感觉和生理学反应的过程,主要包括听觉、触觉、痛觉等[4-6].果蝇也能感受各种机械力,包括重力感觉、听觉、本体感觉、痛觉和触觉等[7-9].对于真核生物,其含有的离子通道中与机械感觉功能相关的基因家族至少有两个,分别是ENaC上皮钠离子通道超家族和TRP(Transient Receptor Potential)超家族.其中TRP超家族中至少有7个亚族,分别是TRPA、TRPC、TRPML、TRPM、TRPN、TRPP和TRPV.这些亚族之间由于具有相似的结构,因而被统称为TRP超家族[10].TRP家族通道蛋白大多数都定位在质膜上,具有6个疏水性的跨膜结构域,其中,第5个和第6个跨膜结构域构成孔道区[11].此外,TRP的N端具有很多锚蛋白重复序列(Ankyrin Repeats, ARs)[12],这些ARs可以把TRP通道蛋白与细胞骨架连接在一起,形成聚合体发挥作用.
NOMPC属于TRPN(Transient Receptor Potential N)离子通道蛋白家族,已经被证实在果蝇触觉和听觉中起作用[13].NOMPC是介导机械力转导的离子通道,也是在国际上第一次鉴定出介导触觉的离子通道[14].NOMPC阳离子通道N端的29个ARs大致起到了细胞内“弹簧”的作用,控制机械门控,连接NOMPC四聚体与细胞骨架[15].NOMPC是6次跨膜蛋白,4个NOMPC亚基组成一个阳离子通道,其中第5次和第6次跨膜结构域组成孔道区.
机械力转导往往通过蛋白复合体进行,例如线虫(C.elegans)中的触觉转导即是以蛋白复合物MEC-4、MEC-2、MEC-6以及MEC-10进行的[16-17].这对于我们研究果蝇NOMPC的转导机制起到参考作用.
已经发现NOMPC蛋白的N端有29个ARs,人类AnkyrinR蛋白中的ARs结构域作为接头蛋白将血影蛋白、肌动蛋白等细胞骨架蛋白连接到膜蛋白[18].但NOMPC中的29个ARs是通过胞内哪种蛋白与细胞骨架结合的尚不清楚.有文献[19]已经表明,NOMPC中的ARs可能与微管存在相互作用,从而介导机械力.通过对NOMPC中的29个ARs一级序列进行分析,发现每个AR中的氨基酸组成均不相同,说明这些ARs可能结合不同的蛋白,行使不同的功能.基于以上研究基础,本实验通过将29个ARs构建到表达载体pUAST-EGFP,使其与EGFP成为融合蛋白,转染果蝇S2细胞后,通过免疫沉淀、质谱分析和Western blot筛选,鉴定出与NOMPC相互作用的蛋白,而后进行蛋白相互作用分析、信号转导机制和在神经元细胞水平和动物的整体水平功能的研究,从而对触觉转导有一个系统、深入的理解.
1 材料与方法
1.1 菌株与质粒
大肠杆菌E.coliDH5α感受态细胞购自江苏康为世纪生物公司;pUAST, pUAST-EGFP, pActin-GAL4和pUAST-NOMPC-EGFP等质粒为本实验室保存质粒.
1.2 试剂、材料与仪器
Q5高保真聚合酶,dNTPs等购自NEB公司;D2000 DNA Marker,500bp DNA Marker等购自天根生化科技(北京)有限公司;蛋白胨,酵母粉,氯化钠,抗生素,NP-40等各种化学药品均购自生工生物工程(上海)股份有限公司;引物合成、DNA测序由上海华津生物科技有限公司完成;重组试剂盒ClonExpress II One Step Cloning Kit购自南京诺唯赞生物科技有限公司;质粒提取试剂盒,DNA片段胶回收试剂盒购自Axygen公司;去内毒素质粒提取试剂盒购自Omega公司.昆虫细胞培养基购自Sigma公司;双抗购自Gibco公司;血清购自Vian Saga公司;试剂GFP-Trap®购自ChromoTek公司;转染试剂MIR 6100Transit-Insect购自Mirus公司;10%或12%聚丙烯酰胺预混液购自江苏新赛美生物科技有限公司;PMSF购自Sigma公司;Rabbit anti-FLAG购自Sigma公司;Mouse anti-GFP购自Abmart公司;Goat anti-mouse抗体购自Abcam公司;Goat anti-rabbit购自金斯瑞公司.PCR扩增仪和蛋白电泳仪均为Bio-Rad产品;凝胶成像仪以及化学发光成像系统为ClinX产品.
1.3 克隆构建
从模板质粒pUAST-NOMPC-EGFP上扩增得到NOMPC 29ARs(引物序列NOMPC 29ARs-F: AACTCTGAATAGGGAATTGGGAATTCGCCACCATGTCGCAGCCGCGCGGAG;NOMPC 29ARs-R: CCCTTGCTCACCATGGTGGCGGTACCGCCGTAGGTGTCGTGCTCCTTG),pUAST-EGFP通过限制性内切酶EcoRⅠ和KpnⅠ双酶切,酶切产物和目的片段凝胶回收,使用一步克隆重组法进行连接,最后测序验证.
通过TRIzol法提取野生型果蝇W1118的总RNA,使用天根公司的cDNA合成试剂盒反转录得到cDNA,以cDNA为模板扩增Annexin B9、CG3731、CG3195和CG5374的cDNA片段,C端加入FLAG tag,引物序列见表1.将扩增得到的Annexin B9(Anx B9)、CG3195和CG5374的cDNA片段和质粒pUAST分别通过限制性内切酶EcoRⅠ/KpnⅠ进行双酶切,酶切产物凝胶回收,用T4DNA连接酶16℃过夜连接,最后测序验证.将扩增得到的CG3731 cDNA片段和质粒pUAST分别通过限制性内切酶EcoRⅠ/XbaⅠ进行双酶切,酶切产物凝胶回收,用T4DNA连接酶16℃过夜连接,最后测序验证.

表1 PCR引物Tab.1 Primers used for PCR

(续表)
1.4 S2细胞培养及转染
S2细胞培养使用Sigma Schneider’s Insect Medium,培养基中加入10%胎牛血清,以及100×青霉素-链霉素,置于28℃无需CO2恒温培养箱中培养.
对于质谱分析方面细胞处理: 传代第二天进行转染,3个100mm培养皿,分别转染Actin-GAL4+pUAST-EGFP、Actin-GAL4+pUAST-NOMPC 29ARs-EGFP、Actin-GAL4+pUAST-NOMPC-EGFP;转染方法见转染试剂说明书.
对于免疫共沉淀(Co-IP)相互作用检测方面细胞处理: 传代第二天进行转染,8皿100mm培养皿细胞,分别转染质粒: Actin-GAL4+pUAST-Annexin B9-FLAG、Actin-GAL4+pUAST-NOMPC 29ARs-EGFP+pUAST-AnnexinB9-FLAG、Actin-GAL4+pUAST-CG3731-FLAG、Actin-GAL4+pUAST-NOMPC 29ARs-EGFP+pUAST-CG3731-FLAG、Actin-GAL4+pUAST-CG3195-FLAG、Actin-GAL4+pUAST-NOMPC 29ARs-EGFP+pUAST-CG3195-FLAG、Actin-GAL4+pUAST-CG5374-FLAG、Actin-GAL4+pUAST-NOMPC 29ARs-EGFP+pUAST-CG5374-FLAG;转染方法见转染试剂说明书.
1.5 免疫沉淀(Immunoprecipitation, IP)
转染48h后收获细胞,加入1mL裂解液(lysis buffer,如NP-40 buffer等,预先加入各种蛋白酶抑制剂),并置于4℃摇晃30min,4℃ 13000r/min 15min离心取上清.加入到20μL 50%体积的准备好的beads悬液中,混匀后置于4℃摇晃2~4h.低盐洗脱,弃上清,加入80~100μL 2×蛋白上样缓冲液(2×SDS loading buffer),95℃变性10min;10000r/min 30s离心,取上清上样进行电泳.
1.6 SDS-PAGE电泳,考马斯亮蓝染色以及质谱分析
取30μL样品,10%胶SDS-PAGE电泳,考马斯亮蓝染色,切下目的胶粒,进行质谱分析.质谱分析交由复旦大学遗传学国家重点实验室平台进行.
1.7 Western blot
Co-IP样品取10μL进行SDS-PAGE电泳,电泳结束后300mA恒流转膜1h,使用5%脱脂奶粉封闭1h,一抗(按照1∶5000稀释)4℃摇床过夜孵育,相应的二抗室温摇床孵育1h,CLINX显影仪显影拍照.
2 结 果
2.1 NOMPC一级结构分析结果
根据NCBI提供的NOMPC的蛋白质序列,分析发现其N端有29个ARs.通过对NOMPC蛋白含有的N端29个ARs一级序列进行分析,发现每个AR中的氨基酸组成不尽相同(图1,见第466页),说明这些ARs能结合不同蛋白,行使不同的功能.
2.2 NOMPC 29ARs的扩增结果
根据上述蛋白质序列分析结果,选取NOMPC N端的29ARs结构进行克隆(图2,见第466页),模板是实验室保存质粒pUAST-NOMPC-EGFP,29ARs的ORF长度为3039bp,PCR扩增完成后,根据一步法克隆,将其连接到目的载体pUAST-EGFP上,同时与EGFP成为融合蛋白.
2.3 对照组GFP、实验组NOMPC 29ARs和实验组NOMPC全长蛋白的纯化结果
将相关质粒转染到S2细胞,48h后收获细胞,经免疫沉淀(IP),以及SDS-PAGE和考马斯亮蓝染色或者银染,即可以显示目的蛋白条带,同时也会获得与目的蛋白相互作用的蛋白(图3,见第466页).由于相互作用的蛋白质含量很少,与银染相比(图3(c)),考马斯亮蓝染色的条带并不清晰(图3(a)、(b)).银染后,可以看到除了目的蛋白,还会有很多的条带,这些可能是与ARs结合的蛋白质,也有可能是目的蛋白降解导致的.由于全长NOMPC分子量比较大,而且NOMCP为膜蛋白,表达量较GFP和NOMPC 29ARs低,从而考马斯亮蓝染色得到的条带相对暗些.

图1 NOMPC N端29个锚蛋白重复序列示意图Fig.1 Sketch of NOMPC N-terminal 29 ARs阴影部分分别代表29个ARs;红色字母分别代表29个ARs中对应的氨基酸.
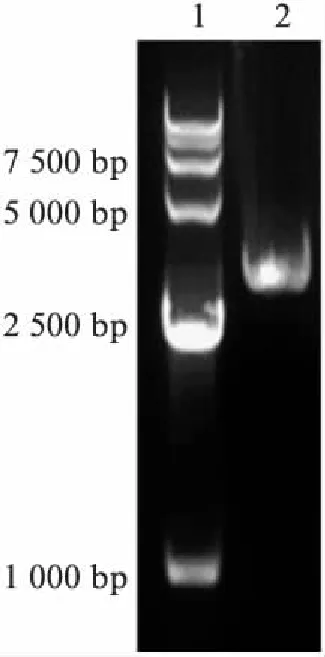
图2 NOMPC 29ARs的扩增Fig.2 PCR of NOMPC 29ARs1. Marker: 15000; 2. NOMPC29ARs(3039bp).

图3 (a) 对照组GFP、(b) 实验组NOMPC 29ARs和(c) NOMPC全长蛋白的纯化以及染色Fig.3 Purified protein samples: GFP, NOMPC 29ARs and NOMPC and staining(a) IP纯化后考马斯亮蓝染色1. 空白S2细胞, 2. GFP, 3. NOMPC 29ARs-GFP;(b) IP纯化后考马斯亮蓝染色1. GFP, 2. NOMPC-GFP;(c) IP纯化后银染1. 空白S2, 2. GFP, 3. NOMPC 29ARs-GFP, 4. NOMPC-GFP.
2.4 NOMPC相互作用蛋白质谱分析结果
上述得到的蛋白质重新做SDS-PAGE,样品跑至最大的Marker进入分离胶,切下胶条,切成1mm3的胶粒,送样进行质谱分析,质谱结果见表2.
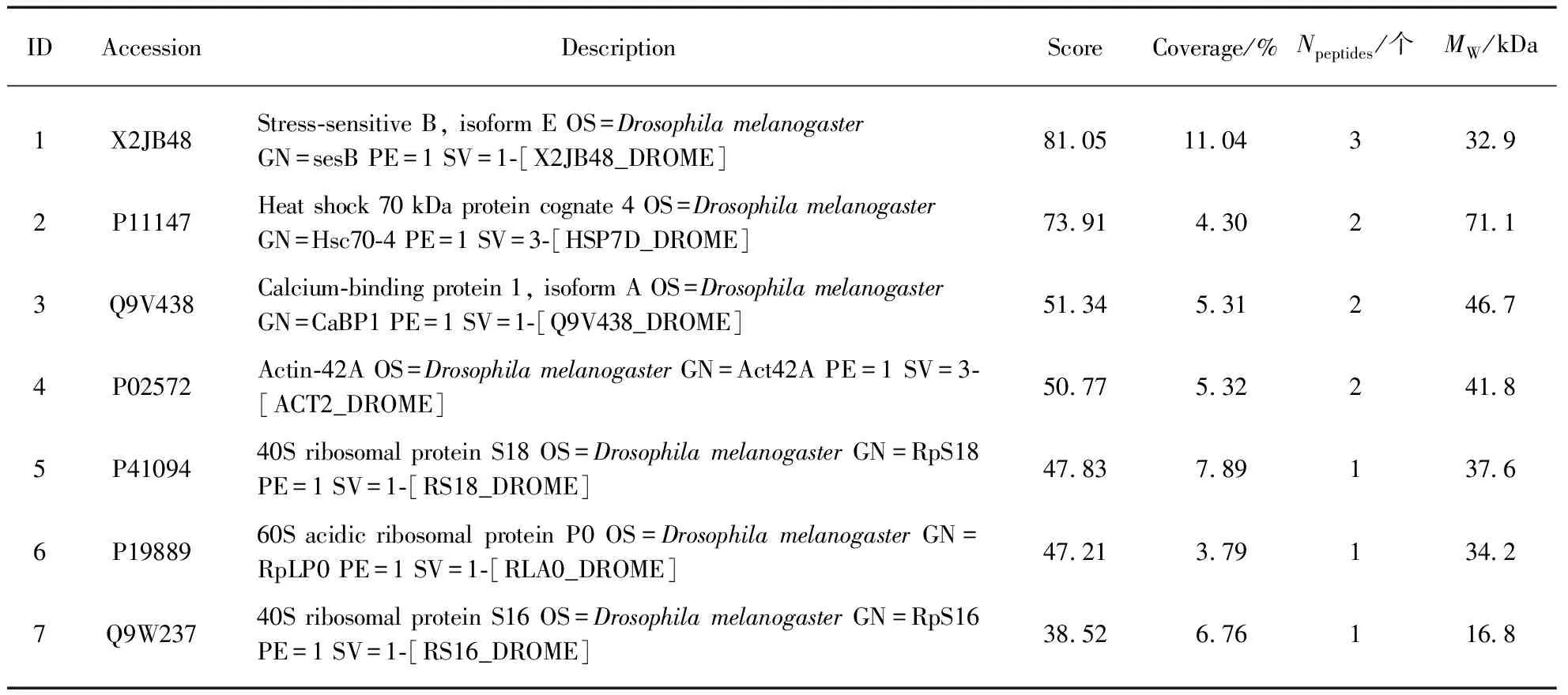
表2 NOMPC相互作用蛋白质谱分析结果Tab.2 Mass spectrum analysis results of NOMPC interaction protein
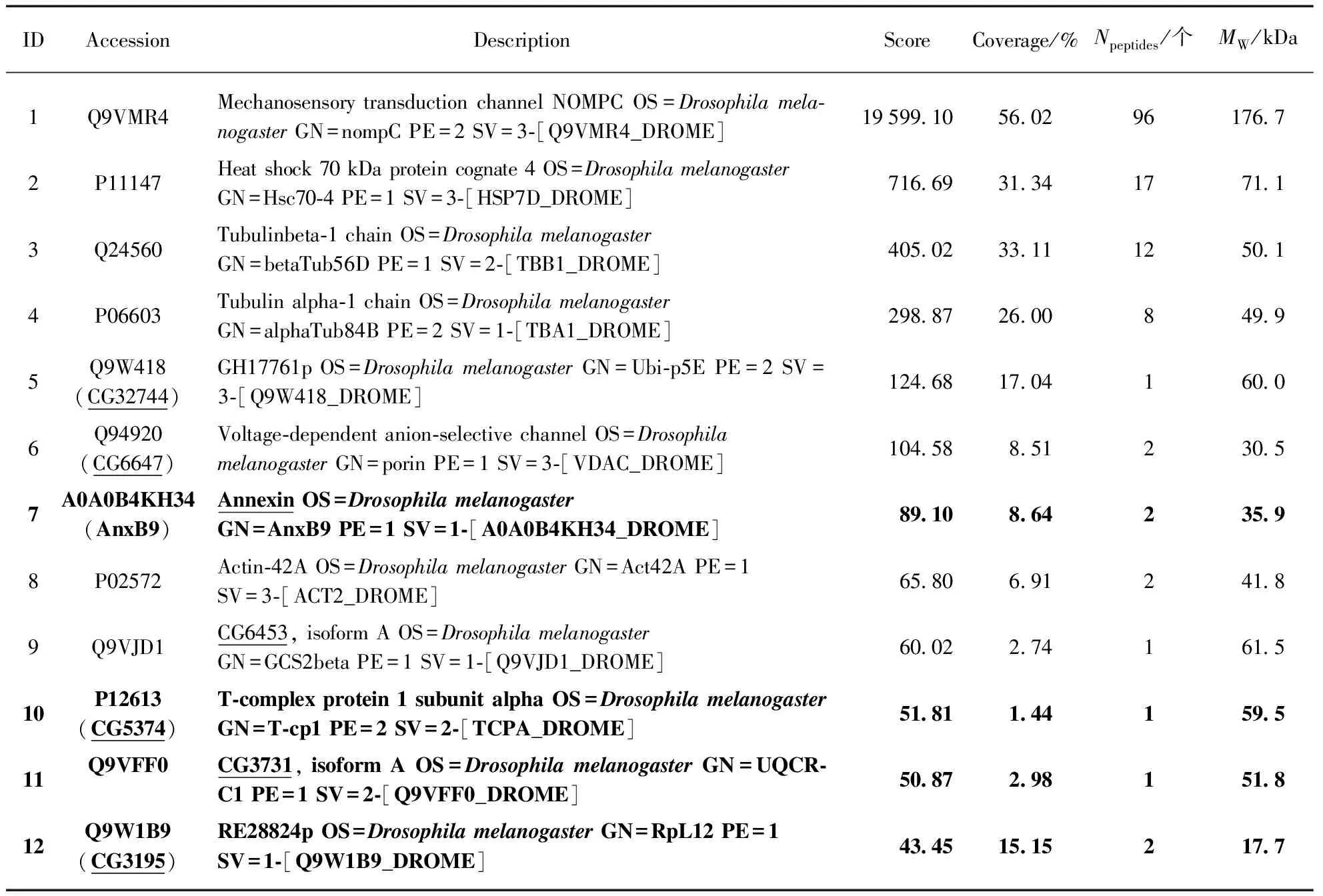
(b) NOMPC 29ARs相互作用蛋白质谱结果

(续表)
注: 表中标示黑体的部分表示与NOMPC 29ARs之间存在相互作用的蛋白质;标下划线的部分表示可能与NOMPC存在相互作用又比较有意义的蛋白质.
从表2(a)可以看出,对照组GFP组得到的蛋白质较少,排除实验组中得到的蛋白质是与GFP存在相互作用的可能性,排除非特异性结合的可能.表2(b)、(c)分别是与NOMPC 29ARs和全长NOMPC相互作用的蛋白质的质谱结果(只选取了一部分),对照组和实验组均重复至少3次实验.可以看出,无论是NOMPC 29ARs还是NOMPC全长蛋白,质谱结果中均说明这两者可能与α-tubulin(表1(b)中ID4,(c)中ID9)和β-tubulin(表1(b)中ID3,(c)中ID13)之间存在相互作用,验证了之前报道的结果,说明NOMPC有可能是在微管的介导下进行机械力的转导[19].此外,从质谱结果看,除了α-tubulin和β-tubulin,可能有互作的又相对有意义的蛋白有: Na pump、Microtubule-associated protein(MAP)、CG1532、CG3195、CG3731、CG5374、CG6453、CG6647、CG8320、CG11857、CG32744、Annexin B9和Calmodulin(表1(b)、(c)中下划线标记部分)等.
2.5 NOMPC 29ARs相互作用蛋白的鉴定
根据质谱结果筛选出的NOMPC相互作用蛋白中,已经验证出Annexin B9、CG3195、CG3731、CG5374(表2(b)中黑体标记部分)与NOMPC 29ARs之间存在直接或者间接的相互作用(图4).其中Annexin已被证明与TRPV4之间存在相互作用[20],而且其与NOMPC全长也存在相互作用(表2(c)中黑体标记部分).

图4 NOMPC 29ARs截短体与CG3195、CG5374、CG3731、Annexin B9相互作用的鉴定Fig.4 The interaction between NOMPC 29ARs truncations and CG3195,CG5374,CG3731,Annexin B9(a) NOMPC 29ARs-GFP与CG3195相互作用的检测: 第一道是转染Actin-GAL4+pUAST-CG3195-FLAG,排除非特异性结合;第二道是转染Actin-GAL4+pUAST-NOMPC 29ARs-GFP+pUAST-CG3195-FLAG;(b) NOMPC29ARs-GFP与CG5374-FLAG相互作用的检测;(c) NOMPC 29ARs-GFP与Annexin-FLAG相互作用的检测;(d) NOMPC 29ARs-GFP与CG3731-FLAG相互作用的检测.
3 讨 论
NOMPC是第一个在动物中发现的介导触觉的机械力感受通道,机械力转导往往通过蛋白复合体进行,AR是蛋白相互作用的一个结构,所以以NOMPC中的ARs为饵蛋白,利用IP手段鉴定其相互作用蛋白,而后分析具体作用,从而对触觉转导有一个系统的、深入的了解.
膜联蛋白B9(Annexin B9)属于Annexin多基因家族-钙离子依赖的膜结合蛋白中的一员,Annexin B9可以与血影蛋白(Spectrin)结合[21].NOMPC ARs结构域作为接头蛋白可能将血影蛋白、肌动蛋白等细胞骨架蛋白连接到膜蛋白,Annexin B9可能通过血影蛋白的连接参与触觉的转导.此外,Annexin B9还可以与actin、钙离子和钙离子依赖的磷脂结合[22],参与钙离子依赖的相关过程.在触觉或者听觉机械转导过程中,钙离子发挥重要的角色,从而Annexin B9在触觉转导的过程中也可能与这些过程相关.CG3731是泛醇-细胞色素c还原酶核心蛋白1,影响间期微管结构[23],已经鉴定出NOMPC与微管存在相互作用,可能是在微管的介导下进行机械转导[19],所以CG3731可能通过影响微管的结构从而对果蝇触觉的转导起到一定的作用.CG5374能起到分子伴侣的作用,帮助蛋白质的折叠,对actin和tubulin的折叠起到一定作用,是微管相关复合物[23],可能也是通过actin或者tubulin对果蝇的触觉转导起到作用.CG3195是核糖体蛋白L12,是核糖体结构组成成分,参与蛋白质翻译过程,对于触觉转导的具体作用还需要进行下一步的实验验证.对于这些已经鉴定出的相互作用蛋白,下一步我们需要选择相应基因的启动子-GAL4果蝇进行细胞定位,观察是否在感音神经元或者ClassⅢ神经元中表达,此外还需要进行亚细胞定位,确定蛋白表达图谱.
在实验过程中,我们采用的是体外过表达体系来筛选NOMPC相互作用蛋白,与果蝇在体中NOMPC相互作用蛋白可能会有些差别,我们的实验并不能完全证实内源性蛋白也存在类似的相互作用,所以我们还需要在果蝇体内进行与NOMPC相互作用的鉴定.
4 展 望
在质谱结果中,还有很多可以供我们分析的蛋白,所以还可以选取一些有兴趣的蛋白质,鉴定是否与NOMPC之间存在相互作用.对于鉴定出的相互作用蛋白,后续可以研究NOMPC蛋白复合物组分在第三类多树突神经元对触觉刺激电生理反应和果蝇触觉行为中的作用.果蝇有非常丰富的遗传资源,全部的基因都有RNAi基因干扰的品系,因此,可以利用第三类多突触神经元特异19-12-Gal4以及相应基因的UAS-shRNA在第三类神经元中将这些基因失活或降低表达,测试突变后第三类多树突神经元对触觉的刺激反应.
[1] NATHANS J, HOGNESS D S. Isolation, sequence analysis, and intron-exon arrangement of the gene encoding bovine rhodopsin [J].Cell, 1983,34(3): 807-814.
[2] BUCK L, AXEL R. A novel multigene family may encode odorant receptors-a molecular-basis for odor recognition [J].Cell, 1991,65(1): 175-187.
[3] HOON M A, ADLER E, LINDEMEIER J,etal. Putative mammalian taste receptors: A class of taste-specific GPCRs with distinct topographic selectivity [J].Cell, 1999,96(4): 541-551.
[4] TSUBOUCHI A, CALDWELL J C, TRACEY W D. Dendritic filopodia, ripped pocket, NOMPC, and NMDARs contribute to the sense of touch inDrosophilalarvae [J].CurrentBiology, 2012,22(22): 2124-2134.
[5] LUMPKIN E A, MARSHALL K L, NELSON A M. The cell biology of touch [J].CellBiol, 2010,191(2): 237-248.
[6] CHALFIE M. Neurosensory mechanotransduction [J].NatRevMolCellBiol, 2009,10(1), 44-52.
[7] TRACEY W D, WILSON R I, LAURENT G,etal. Painless, aDrosophilagene essential for nociception [J].Cell, 2003,113(2): 261-273.
[8] GONG Z, SON W, CHUNG Y D,etal. Two interdependent TRPV channel subunits, inactive and Nanchung, mediate hearing inDrosophila[J].Neurosci, 2004,24(41): 9059-9066.
[9] KAMIKOUCHI A, INAGAKI H K, EFFERTZ T,etal. The neural basis ofDrosophilagravity-sensing and hearing [J].Nature, 2009,458(7235): 165-171.
[10] 邹文娟,黄桂芳,康利军.TRP 通道在生物体对机械性刺激响应中的功能及作用机制 [J].浙江大学学报(医学版),2012,41(002): 222-228.
[11] LIANG X, MADRID J, SALEH HS,etal. NOMPC, a member of the TRP channel family, localizes to the tubular body and distal cilium ofDrosophilacampaniform and chordotonal receptor cells [J].Cytoskeleton, 2011,68(1): 1-7.
[12] SHIN J B, ADAMS D, PAUKERT M,etal.XenopusTRPN1(NOMPC) localizes to microtubule-based cilia in epithelial cells, including inner-ear hair cells [J].ProcNatlAcadSciUSA, 2005,102(35): 12572-12577.
[13] ZHANG W, YAN Z, Jan L Y,etal. Sound response mediated by the TRP channels NOMPC, NANCHUNG, and INACTIVE in chordotonal organs ofDrosophilalarvae [J].ProcNatlAcadSciUSA,2013,110(33): 13612-13617.
[14] YAN Z, ZHANG W, HE Y,etal.DrosophilaNOMPC is a mechanotransduction channel subunit for gentle-touch sensation [J].Nature, 2013,493(7431): 221-225 .
[15] LIANG X, MADRID J, GARTNER R,etal. A NOMPC-dependent membrane-microtubule connector is a candidate for the gating spring in fly mechanoreceptors [J].CurrentBiology, 2013,23(9): 756-763.
[16] DRISCOLL M, CHALFIE M. Developmental and abnormal cell death inC.elegans[J].TrendsNeurosci, 1992,15(1): 15-19.
[17] HUANG M, CHALFIE M. Gene interactions affecting mechanosensory transduction inCaenorhabditiselegans[J].Nature, 1994,367(6462): 467-470.
[18] MICHAELY P, TOMCHICK D R, MACHIUS M,etal. Crystal structure of a 12 ankyrin repeat stack from human AnkyrinR [J].EMBO, 2002,21(23): 6387-6396.
[19] ZHANG W, CHENG L E,KITTELMANN M,etal. Ankyrin repeats convey force to gate the NOMPC mechanotransduction channel [J].Cell, 2015,162(6): 1391-1403.
[20] HUAI J, ZHANG Y, LIU Q M,etal. Interaction of transient receptor potential vanilloid 4 with annexin A2 and tubulin beta 5 [J].NeuroscienceLetters, 2012,512(1): 22-27.
[21] TJOTA M, LEE S K, WU J,etal. Annexin B9 binds to {beta}H-spectrin and is required for multivesicular body function inDrosophila[J].JournalofCellSci, 2011,124(17): 2914-2926.
[22] GOLDSTEIN L S B, GUNAWARDENA S. Flying through theDrosophilacytoskeletal genome [J].JournalofCellBiol, 2000,150(2): F63-F68.
[23] HUGHES J R, MEIRLES A, FISHER K H,etal. A microtubule interactome: Complexes with roles in cell cycle and mitosis [J].PLoSBiology, 2008,6(4): 785-795.
ScreeningandIdentificationofProteinsInteractingwithDrosophilaNOMPC
ZHENGHonglan,GUOYifeng,YANZhiqiang
(SchoolofLifeSciences,FudanUniversity,Shanghai200438,China)
DrosophilaNOMPC, a member of the transient receptor potential(TRP) family of ion channels, is a mechanotransduction channel subunit for gentle-touch sensation.However, the transduction mechanism of NOMPC and the protein complexes of touch transduction are largely unclear. In order to study these questions, we obtained NOMPC 29 Ankyrin Repeats(ARs)at N-terminal from NOMPC cDNA, and the open reading frame(ORF) of NOMPC 29 Ankyrin Repeats(ARs)was 3039bp in length. Then we constructed pUAST-NOMPC 29 ARs-EGFP expression vector, and transfected it into S2 cells. Using immunoprecipitation(IP), mass spectrometry(MS) and Western blot approaches, we screened many proteins interacting with NOMPC 29 ARs or NOMPC full length. The interactions between NOMPC and some selected proteins were confirmed by Co-Immunoprecipitation(Co-IP). We found that there were interactions among Annexin B9, CG3731,CG3195,CG5374 and NOMPC in S2 cell. Our data provides clues for further understanding the transduction mechanism of NOMPC.
Drosophilamelanogaster; touch; NOMPC; binding proteins
0427-7104(2017)04-0463-09
2016-11-02
科技部重大科学研究计划(2016YFA0502800),国家自然科学基金面上项目(31571083),“千人计划”青年人才(中组部,2015),上海高校特聘教授(“东方学者”)(TP2014008),上海市青年科技启明星计划(14QA1400800)
郑红兰(1991—),女,硕士研究生;闫致强,男,研究员,通信联系人,E-mail: zqyan@fudan.edu.cn.
Q26
A
