4周低氧运动对营养肥胖型大鼠骨骼肌自噬相关因子时序性变化的影响
2017-11-29张素娴朱磊冯连世刘洪珍路瑛丽
张素娴 朱磊 冯连世 刘洪珍 路瑛丽
1曲阜师范大学(曲阜 273165)
2国家体育总局体育科学研究所
自噬是真核生物中广泛存在的一种现象,是细胞将自身受损、衰老、畸形的细胞器或生物大分子包裹成自噬体,运往溶酶体进行分解,对清除体内的受损或者不再需要的细胞器或蛋白,维持细胞正常的机能具有重要意义。早前认为自噬是由细胞死亡性程序诱导的细胞死亡,但随着研究的深入,人们发现自噬是对低氧、营养不足、运动等条件的一种应激反应。低氧可以通过低氧诱导因子 1α(hypoxia-inducible factor 1α,HIF-1α)诱导自噬的发生,并且在低氧条件下由于氧供应不足造成活性氧(reactive oxygen species,ROS)的大量生成,ROS会攻击正常的细胞器造成自噬。机体自身存在一定的修复功能,可以通过调控氧化还原水平,使机体机能处于相对稳定的状态。细胞自噬就是其中极其重要的一种调控方式,通过降解受损或者老化的细胞器或细胞,维持机体正常氧化还原的相对平衡[1,2]。
Beclin-1是Atg6的同源基因,是最早发现的与自噬相关的基因之一,是自噬信号三大通路之一的Beclin-1信号通路的关键因子,通过与Ⅲ型磷脂酰肌醇3磷酸激酶(ClassIII PI3K)结合调节其他自噬基因编码蛋白与前自噬体膜的结合[3],启动自噬的发生。微管相关蛋白1轻链3(Linght Chain 3,LC3)在自噬体的形成与降解中发挥重要作用的蛋白,自噬小体的延伸离不开LC3的泛素化作用,LC3-Ⅱ与即将发生降解作用的自噬小体外膜上的磷脂酰乙醇胺(Phosphatidylethanolamine,PE)特异性结合定位于自噬体膜上,是自噬体形成的标志,是目前使用最广泛的自噬分子标记物,与自噬体膜的数量呈正相关[4]。泛素结合蛋白P62(Protein 62,P62)是一种重要的衔接蛋白,参与蛋白的选择性降解过程,与自噬体膜上LC3-Ⅱ特异性结合后将自噬体运往溶酶体进行降解[5]。
目前大量的研究证明适量的运动也可通过增强肌肉细胞的自噬,加速受损细胞的降解,为肌纤维的再生提供原料;但是过度训练则会造成细胞过度自噬,导致细胞内的蛋白与细胞器的降解量增大,机能正常的细胞器同样受到降解,引起肌肉的疲劳与损伤,免疫机能下降甚至造成大量细胞的自噬性凋亡[6,7],但是关于低氧运动对营养肥胖型大鼠骨骼肌自噬时序性变化影响的研究还相对较少,本实验从低氧训练的角度出发,探究在低氧环境下运动训练对自噬的影响,以及低氧运动诱导骨骼肌细胞发生自噬的可能机制。
1 材料与方法
1.1 实验动物及分组
SPF级5周龄雄性SD大鼠120只,体重176.4±10.75 g,由北京维通利华实验动物技术有限公司购得,许可证号:SCXK(京)2012-0001。所有大鼠饲养于国家体育总局体育科学研究所动物房,随机挑选出20只进行普通饲料饲养,100只高脂饲料(45%脂供能,Research Diets公司)饲养,两组大鼠间起始体重无明显差异。所有大鼠自由饮食,室温22℃ ±1℃,湿度55% ±2%,每天12 h轮换照明。高脂饮食8周后,挑选肥胖大鼠。营养肥胖型大鼠模型构建成功的判断标准:实验组个体体重与C组均值相比增加超20%,与脂代谢相关血液指标变化显著,Lee’s指数显著上升[8,9]。
建模成功大鼠继续高脂饮食,首先进行1周的适应训练(速度由16 m/min逐渐增加到25 m/min,时间由20 min/d逐渐增加到60 min/d)。从肥胖大鼠中选取50只能够适应跑台训练的大鼠随机分为对照组(C组,常氧未训练)、低氧训练1周组(E1组)、低氧训练2周组(E2组)、低氧训练3周组(E3组)、低氧训练4周组(E4组),每组10只。各组体重无显著性差异。低氧暴露模拟13.6%氧浓度,训练组进行跑台耐力训练,坡度为0,速度为20 m/min,60 min/d,5 d/w,周末休息。分别训练0周、1周、2周、3周、4周。
1.2 取材
各组大鼠于末次训练结束后恢复24 h进行取材,取材前12 h清空饲料,腹腔注射10%水合三氯乙醛溶液麻醉(0.4 ml/100 g体重);取右侧腓肠肌,置于冻存管中,先放入液氮中骤冷,后放入-80℃超低温冰箱保存。
1.3 主要实验仪器
匀浆机(IKA WERKE)、酶标仪(Labsystems Dragon)、电泳仪(BIO-RAD)、化学发光成像系统(BIORAD)、低温离心机(赛默飞)、RCP扩增仪(Applied Biosystems)、实时荧光PCR仪(Applied Biosystems)、核酸扩增仪(9700,ABI)。
1.4 测定指标及方法
1.4.1 Real- Time PCR 检测Beclin- 1、P62 和LC3mRNA的表达
所有引物序列均由上海生物工程技术有限公司以GeneBank数据库为参照模板,设计合成,Beclin-1-F引物序列ggccaataagatgggtctga,Beclin-1-R引物序列gctgcacacagtccaaaaa;LC3-F引物序列aacgtaggcacccacatagg,LC3-R引物序列gaagagactgcccctgacac;P62-F引物序列attaccaccccactgga,P62-R引物序列gtgacaaggaggactggcat。
RT-PCR检测:采用 Trizol法提取腓肠肌中的总RNA,使用反转录试剂盒(RR037A,TakaRa)将所提取的骨骼肌总RNA在核酸扩增仪中反转录获取cDNA,利用荧光定量 PCR仪进行样本cDNA的扩增与荧光定量检测。反应条件:起始模板变性95℃,30 s;PCR循环中变性95℃,5 s;60℃,31 s,40个循环。荧光定量PCR得到3个重复样本Ct值,取平均值,将实验数据代入公式计算2-△△Ct值进行相对定量计算。
1.4.2 Western Blot法检测Beclin-1、P62和LC3蛋白的表达
首先提取总蛋白,使用BCA法测试得到蛋白浓度并调至统一值,常规Westent blot法检测,一抗为(Anti-LC3A/B、Anti-SQSTM1/P62和 Anti-Beclin 1、内参为GAPDH,Abcam,美国),稀释比例均为1∶1000。HRP标记的抗鼠和抗兔的二抗(Abcam,美国)稀释,稀释比例均为1∶5000,ECL发光液(Millipore,美国)显影后用ImageJ软件进行图像灰度值分析。
1.5 统计学分析
所有数据均用SPSS 19.0软件进行统计学分析,结果采用平均数±标准差(±s)表示。P<0.05表示具有显著差异,P<0.01表示具有非常显著差异。采用单因素方差分析法比较组间差异,最终所得实验数据利用 GraghPad Prism 5.0软件汇总分析做出图像。
2 结果
2.1 各组大鼠Beclin-1相对表达量的时序性变化
与C组相比,E1组、E2组、E3组大鼠腓肠肌Beclin-1 mRNA 的相对表达量升高显著(P<0.05,图1A)。Beclin-1蛋白的相对表达量呈现先下降后上升的趋势,与C组相比,E2组Beclin-1蛋白的相对表达量显著下降(P<0.05),且E2组非常显著低于E4组(P<0.01),E3组显著低于E4组(P<0.05,图1B),E3组与E2组相比变化不显著。
2.2 各组大鼠P62相对表达量的时序性变化
E1组P62 mRNA的相对表达量显著高于C组(P<0.05);E2组P62 mRNA的相对表达量显著高于E1组(P<0.05),非常显著高于C对照组(P<0.01);E3组非常显著低于E2组(P<0.01),E4组非常显著高于C组、E1组合E3组(P<0.01,图2A);E3组P62蛋白相对表达量与E2组相比呈显著增高趋势(P<0.05,图2B)。
2.3 各组大鼠LC3相对表达量的时序性变化
与C组相比,E3组LC3 mRNA相对表达量显著升高(P<0.05),E2、E4组非常显著升高(P<0.01)(图3A);各组大鼠LC3蛋白中LC3-Ⅱ/LC3-Ⅰ比值没有显著性变化。
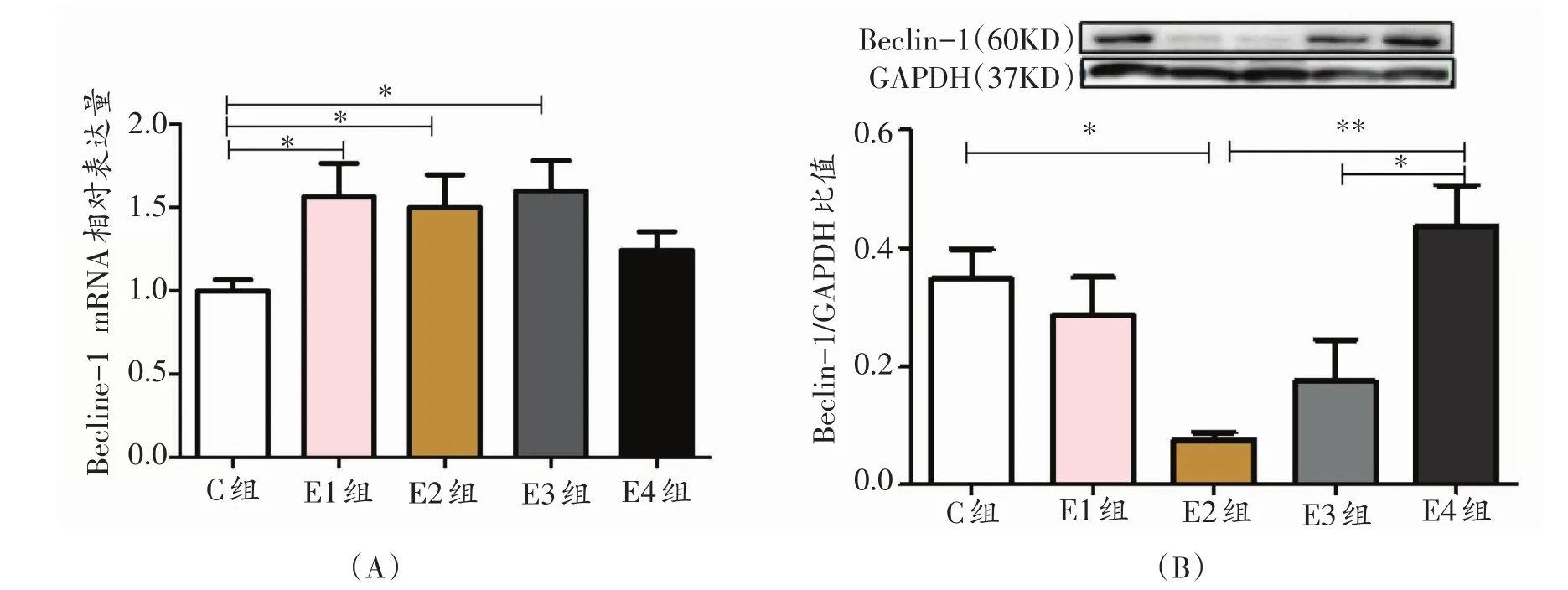
图1 各组大鼠自噬相关因子Beclin-1相对表达量
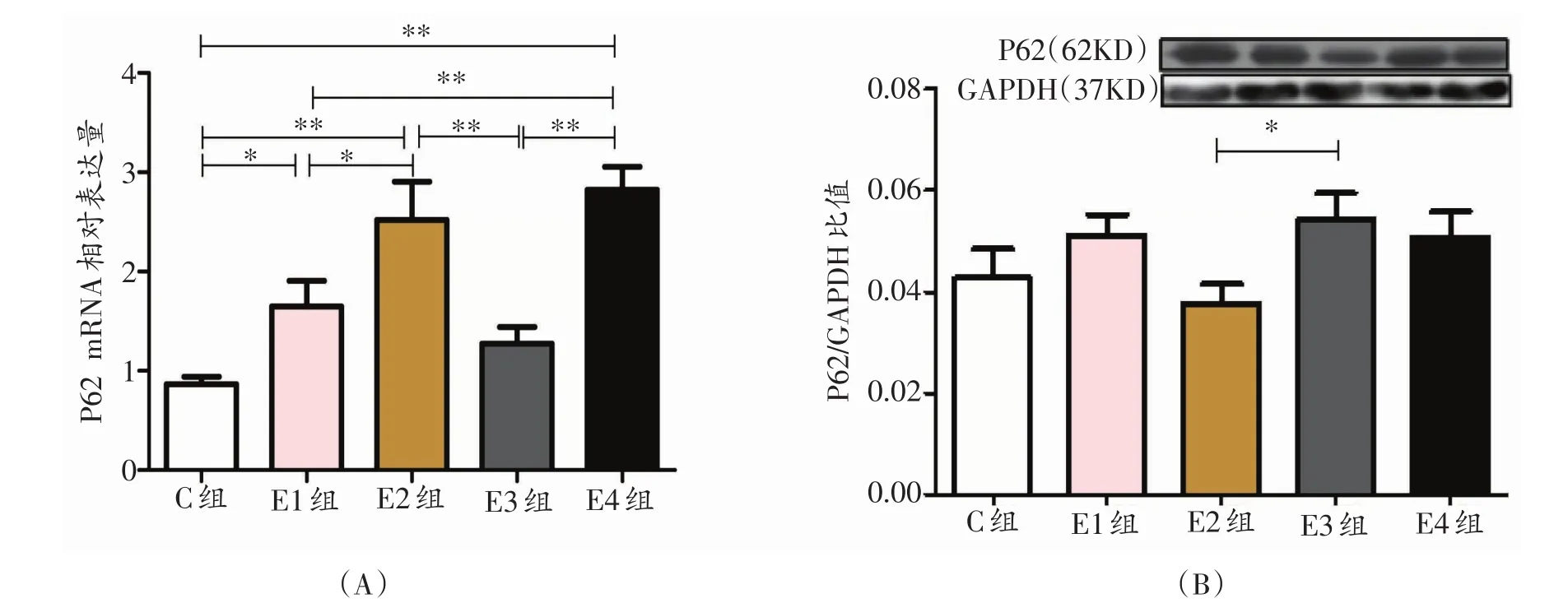
图2 各组大鼠自噬相关因子P62的相对表达量
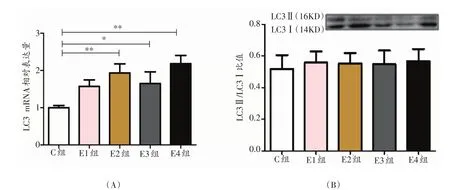
图3 各组大鼠自噬相关因子LC3的相对表达量
3 讨论
正常情况下,机体为了维持自身内环境的相对稳定,需不断降解功能失常、老化或者发生变异的细胞器、蛋白质以及胞质组分,通过形成自噬小体运往溶酶体进行降解。自噬的过程可分为自噬的启动、自噬前体的延伸、成熟以及自噬小体的降解4个不同的阶段[10]。正常情况下自噬处于较低水平,但是营养缺乏、衰老、低氧、运动等刺激可以增加自噬的发生,自噬启动后,磷酸肌酸三磷酸激酶Ⅲ型PIK(Vps34)与Beclin-1结合启动膜泡的成核[11]。自噬启动后的延伸主要依靠复合物与Atg12的结合以及与LC3的修饰过程[12],在自噬过程中起重要作用,主要是因为Atg12的结合过程与前自噬体结构的生成密切相关,LC3的修饰与自噬小体生成密不可分。LC3是Atg8的同系物,具有2个亚型,当自噬启动后,LC3-Ⅰ泛素化修饰形成LC3-Ⅱ,LC3-Ⅱ与自噬小体膜上的磷酸酰乙醇胺(PE)结合后定位到自噬体膜上,是自噬体存在的标志物。在自噬小体被运往溶酶体降解后,LC3-Ⅱ即被溶酶体中的水解酶分解消失[13]。SQSTM1/p62是位于自噬体膜上的受体蛋白,对自噬体的识别包裹具有重要作用[14]。P62可作为一种压力传感器,与LC3-Ⅱ相互作用,促进自噬小体向溶酶体的转运降解[15]。自噬小体与溶酶体结合后,溶酶体中的水解酶将自噬小体中包裹的内容物降解,一方面为细胞的再生提供原料,另一方面用于维持内环境的相对稳定。
Beclin-1是哺乳动物参与自噬的重要因子[16],不能单独作用于自噬细胞,而是要与细胞内的某些分子结合才能发挥作用,如促进自噬作用的mVPs34和有抑制自噬作用的Bcl-2等[17,18]。大量研究发现低氧环境可通过调节Beclin-1来调控自噬程度达到适应低氧环境的目的,低氧、缺血再灌注等刺激下机体会产生应激反应,Beclin-1活性升高,诱导细胞自噬的发生[19,20]。Gurusamy[21]等的研究证实缺血缺氧条件下可以增加自噬的发生,从而保护心肌细胞。Zhang[22]等实验研究证明,低氧条件下线粒体自噬活性的增强是通过HIF-1α实现的,提示线粒体自噬的发生可能是对低氧环境的一种适应性变化。Band等[23]在随后的实验中证明Zhang实验成功的关键是具有BniP3,在低氧环境中通过HIF-1α诱导BniP3活性的增强,BniP3再通过激活下游受体蛋白Beclin-1从而提高线粒体自噬的活性。唐中园等[24]研究发现在常氧环境下,体外培养的卵癌细胞经质粒转染HIF-1α基因后Beclin-1的表达量与LC3-Ⅰ向LC3-Ⅱ转换的效率都有所增加,低氧环境中适应一段时间后敲除HIF-1α基因,Beclin-1蛋白浓度随后出现下降趋势,之前低氧环境下已经生成的LC3-II的活性又受到抑制。但是Weng等[25]的研究发现在12%氧浓度下,运动可促进Bcl-2表达增强,抑制Beclin-1的表达。Kim等[26]观察2月龄小鼠8周跑台运动后同样发现跖长伸肌中Beclin-1含量呈显著下降趋势,出现这种现象的原因可能是检测的组织不同,或者是运动强度的不同,但Weng和Kim的结果却与本实验中观察到的Beclin-1蛋白前两周变化趋势基本相同。本实验观察到的Beclin-1 mRNA表达量的变化与上述诸位学者的报道较为一致,但是需要注意的是低氧训练对肥胖大鼠骨骼肌Beclin-1 mRNA和蛋白的影响出现完全相反的变化趋势。引起这种变化的原因可能是Beclin-1在转录后水平受到某些未知因素的调控,例如microRNA或者siRNA引起基因沉默或者mRNA降解,导致Beclin-1蛋白表达的降低;蛋白的表达相较与基因的转录具有一定的延迟性,需要经过复杂的翻译过程以及翻译后的修饰过程才能成为具有生物活性的蛋白;此外Salminen[27,28]等的研究发现,在运动干预2~7天后自噬程度达到顶峰,自噬溶酶体中出现不同降解程度的线粒体,降解的底物重新被细胞吸收利用,本实验中Beclin-1蛋白在低氧运动前两周出现下降趋势,可能是在低氧运动1周后自噬程度达到最大值,随后出现下降,但随着运动时间的延长自噬程度又有所增加,可能是机体逐渐适应低氧运动的结果。
LC3是自噬的标志物,当自噬发生时,LC3被Atg4切割成LC3-Ⅰ,接着在Atg3的作用下转化为LC3-Ⅱ,定位在自噬体膜上,可与P62特异性结合。 LC3-Ⅱ/LC3-Ⅰ的比值可以更客观地反映自噬水平的高低,比值升高反映出自噬水平的增加,反之自噬水平降低[29]。Grumati[30]等在利用野生型小鼠进行一次性递增负荷运动时发现LC3-Ⅱ/LC3-Ⅰ比值增加了5.8倍,马晓雯等[31]在实验中发现16周长时间中等强度运动可以上调Beclin-1与LC3 mRNA表达量,提高LC3-Ⅱ/LC3-Ⅰ比值,长时间大强度运动会使自噬体异常增多,引起心肌过度自噬,造成损伤。运动方式不同,对自噬的结果的影响却是类似的。Lira[32]发现经过4周自主运动后,LC3-Ⅱ/LC3-Ⅰ比值显著增大,P62蛋白表达量下降,认为自噬对骨骼肌正常功能的维持有重要意义,是骨骼肌对运动的适应性反应,可提高机体运动能力。Chen[33]对心梗小鼠进行跑台实验4周后,发现实验组LC3-Ⅱ/LC3-Ⅰ比值较对照组增加210%,并推测运动可能是通过激活细胞自噬活性,加速损伤细胞器与蛋白的降解,减轻运动对心肌的损伤。这些都与本实验中LC3 mRNA表达量结果相一致,适宜的运动对机体机能有一定程度的促进作用,但是运动量过大则会造成自噬过度激活引起损伤,Feng[6]等的研究发现训练过度可使肌肉组织中的自噬程度激活过度,Beclin-1、LC3表达量明显升高,过度降解肌肉组织中完好的蛋白或细胞器,导致免疫力下降,肌肉出现萎缩。本实验中发现的LC3 mRNA表达量与上述学者的研究结果相一致,Smuder[34]等利用大鼠进行强度为70%VO2max的运动干预5天后发现Atg12、Atg5-Atg1、LC3、LC3-Ⅱ/LC3-Ⅰ等与自噬相关蛋白的含量均无显著性变化,这与本实验的LC3-Ⅱ/LC3-Ⅰ蛋白比值变化趋势相同,出现mRNA表达量与蛋白表达量不一致的原因可能是在基因转录生成mRNA再翻译成目的蛋白的过程中还存在转录后水平的调控,转录后的mRNA还需要有进一步的加工过程,在此过程中一些RNA会发生沉默或者降解,翻译成前体蛋白后还要经过一定程度的修饰,蛋白的半衰期较mRNA的半衰期长,不容易降解,并且在机体内本身就存在一定数量的前期积累蛋白,由此导致LC3蛋白表达量未能出现与mRNA的表达量相同的趋势。
P62是溶酶体识别自噬小体的受体蛋白,是一种泛素化蛋白,参与自噬小体的降解过程,同时在泛素蛋白酶系统的蛋白降解过程也发挥极其重要的作用[15]。它在自噬活动中受到双向调节,一方面受细胞自身自噬活性的影响,一方面又可以通过mTORC1信号通路负向调控自噬活性。可与LC3-Ⅱ结合促进自噬小体的降解[15]。机体在营养供应不足的情况下为了保证细胞功能的运转会非选择性地降解部分细胞,增加自噬作用,而P62是参与选择性的自噬作用,保证正常细胞的质量,降解受损的蛋白与细胞器[35]。P62还与细胞的抗氧化、衰老、肥胖、癌症等有重大关联[35]。P62的浓度与自噬呈负相关,反映自噬溶酶体的活性,也是自噬强弱的标志性分子。祖靓等[36]研究发现力竭运动后即刻,LC3-Ⅰ、LC3-Ⅱ蛋白相对表达量以及LC3-Ⅱ/LC3-Ⅰ蛋白相对比值显著增加,LC3-Ⅱ/LC3-Ⅰ比值在恢复期下调,P62蛋白在运动后即刻增加,恢复期下降。崔迪等[37]通过对营养肥胖型小鼠进行6周的耐力训练后发现,普通饮食联合运动干预组P62蛋白以及mRNA表达量均显著低于普通饮食干预组,并且LC3-Ⅱ/LC3-Ⅰ比值显著增加。这与本实验的P62 mRNA与蛋白表达量的变化存在一定的差异性,P62的这种变化可能是由于转录后水平的调控;也可能是其他别的原因,如肥胖抑制自噬的发生;另外,机体还存在一定的蛋白储备,这些蛋白不参与机体的调节作用。而且RT-PCR检测的灵敏性高于Westent blot蛋白的检测水平,这些都有可能是mRNA的变化与蛋白的变化不一致的原因。值得注意的是E3组P62 mRNA较E2以及E4组极显著性降低,具体原因尚不清楚,有待进一步研究。
自噬与肥胖之间同样存在密切关联。实验发现先天性肥胖与高脂饮食造成的肥胖小鼠中,LC3、Beclin-1、Atg5、Atg7表达水平明显减少,电镜下样本中自噬体的数量显著减少[38],自噬程度降低。但是还有研究发现,营养肥胖型大鼠下丘脑中的Atg7蛋白减少49%,Atg5蛋白减少36%,LC3-Ⅱ/LC3-Ⅰ比值下降,但是Beclin-1、Atg5、Atg7、Atg12 mRNA表达量在实验前后变化不大,猜测营养肥胖型小鼠下丘脑的自噬水平的变化可能是发生在转录后水平的调控[39,40]。另外,薄海[41]等研究发现,与低氧对照组相比,4周低氧联合运动组大鼠PINK1蛋白表达量下降显著,同时线粒体膜电位与ATP输出能力显著升高,ROS生成速率明显下降,说明低氧联合运动抑制了低氧环境对线粒体的损伤[41]。
此外,4周低氧训练对高脂饮食诱导的营养肥胖型小鼠的骨骼肌细胞自噬相关基因表达量变化的影响较大,但对自噬相关蛋白的表达影响不大。提示有可能是基因转录后水平的调控所造成的这种不一致性,在基因转录生成mRNA到翻译成蛋白的过程中,存在一定的时间与位置的差异,在这个过程中可能发生一系列的变化,例如,microRNA或者 siRNA引起基因沉默或者 mRNA降解;mRNA在机体存在时间较短,翻译成蛋白后相对稳定,导致自噬相关蛋白与mRNA表达量不一致;并且RT-PCR检测比Westent blot检测的灵敏性要高;机体本身也有一定的蛋白储备且不参与机体的调节;也可能是肥胖造成了自噬相关蛋白表达量的降低,具体是哪种原因造成的这种差异性,有待进一步的研究。
4 结论
低氧运动能显著提升自噬基因的相对表达量,在一定程度上促进自噬的发生,对于维持机体细胞内环境的稳态具有重要作用。自噬蛋白与基因表达具有不同步性,可能是由于基因转录后水平的调控,也可能是由于肥胖对自噬造成一定的影响,但最终行使功能的是蛋白,所以最终要根据蛋白水平的变化判断其对相应功能的影响。在低氧运动的刺激下,骨骼肌细胞的自噬程度在前2周出现增强的趋势,但是随着运动时间的延长,自噬的程度又有一定的降低,可能是机体对低氧运动刺激逐渐适应的结果。
